Neuroscience of the Unconscious Mind and Repressed Memories
The conceptualization of the unconscious mind and the mechanics of memory have undergone a profound theoretical and empirical evolution over the past century. Originating with nineteenth-century psychoanalytic theory, the proposition that human behavior is predominantly driven by forces residing outside of conscious awareness revolutionized psychological thought. Sigmund Freud posited an active, dynamic unconscious - a mental reservoir of repressed drives, traumatic memories, and unacceptable impulses kept hidden by psychological defense mechanisms 12. Today, modern cognitive neuroscience, supported by advanced neuroimaging, connectomics, and computational modeling, has fundamentally re-evaluated these claims. Through the lenses of predictive processing, active inference, and the neurobiology of memory retrieval, contemporary science has largely discarded the Freudian concept of a spatially distinct, shadow-mind governed by primitive drives 34. Instead, it recognizes a highly distributed cognitive unconscious where identical neural networks govern both conscious and non-conscious processing 45.
This report provides an exhaustive analysis of how repressed memories, trauma-induced dissociation, and non-conscious expectations function within modern neurobiology. By examining the neural circuitry of motivated forgetting, the reconstructive nature of memory, and cross-cultural neurodevelopmental variances, this analysis delineates which elements of Freudian theory find empirical support in contemporary neuroscience and which have been supplanted by modern cognitive paradigms.
Theoretical Evolution of the Unconscious Mind
The division between classical psychoanalysis and modern neuroscience fundamentally centers on the architecture, intent, and function of non-conscious mental processing. Contemporary research establishes the existence of a robust unconscious, but its nature diverges sharply from early theoretical models 52.
The Psychoanalytic Dynamic Unconscious
In psychoanalytic theory, the unconscious mind is defined primarily by the mechanism of repression. It is theorized as an active, compartmentalized region of the psyche containing conflictual, anxiety-producing content - frequently rooted in early developmental stages - that exerts constant upward pressure toward consciousness but is actively blocked by the ego's defense mechanisms 12. The content of this dynamic unconscious is considered inaccessible to direct introspection, revealing itself only through disguised manifestations such as dreams, neurotic symptoms, or slips of the tongue 1.
Historically, this framework faced severe scientific criticism. Philosophers of science, such as Karl Popper, argued that the Freudian unconscious was unfalsifiable, as any contradictory evidence could simply be interpreted as further resistance or disguised repression 2. Furthermore, early critics like Pierre Janet warned that Freud erred by generalizing phenomena observed in a small sample of abnormal, highly distressed patients to the everyday mental life of all human beings 2. In response to the lack of replicable methods for studying internal mental states, early twentieth-century behaviorism discarded the study of the unconscious entirely, focusing solely on observable stimuli and responses 2. It was not until the cognitive revolution of the 1960s that internal mental processes were legitimized as objects of scientific inquiry, leading to the current understanding of implicit cognition 2.
The Modern Cognitive Unconscious
The modern cognitive unconscious encompasses all information processing that occurs without conscious awareness yet measurably influences behavior, decision-making, and emotional regulation 35. This framework includes implicit memory, subliminal perception, automatic motor control, and heuristic decision-making 15. Cognitive neuroscience has demonstrated that there is no separate "unconscious mind" playing by distinct rules or residing in a privileged neural region 34. Instead, identical brain regions are activated, and the same underlying computational processes occur, regardless of whether stimuli are presented supraliminally (consciously perceived) or subliminally (below the threshold of awareness) 4.
The cognitive unconscious lacks the intentionality to actively hide specific psychological conflicts in the Freudian sense; rather, it reflects the brain's inherent architecture for processing massive amounts of sensory and associative data efficiently. While some cognitive scientists previously viewed the unconscious merely as a passive shadow of conscious thought, current evidence suggests it is equally complex and flexible, seamlessly integrating cognitive, affective, and motivational processes without requiring conscious oversight 32. Recent advances in connectomics - the comprehensive mapping of the brain's neural connections via diffusion magnetic resonance imaging (dMRI) - further support this by localizing psychiatric conditions to structural alterations in white matter networks rather than localized repositories of repressed trauma 5.
Neural Mechanisms of Motivated Forgetting
While neuroscience rejects the concept of an autonomous repressed unconscious, it provides strong empirical support for the human capacity to intentionally exclude unwanted memories from awareness - a phenomenon modern cognitive psychology terms "motivated forgetting" 34. The ability to halt the retrieval of an unwelcome memory shares a conceptual lineage with Freudian repression, but it is studied as an active, executive control mechanism rather than an involuntary, passive defense 510.
Research utilizing the "Think/No-Think" (TNT) paradigm has identified the precise neural circuitry utilized when individuals attempt to suppress the retrieval of a memory in the presence of a strong reminder 36. Functional magnetic resonance imaging (fMRI) reveals two distinct, opposing mechanisms that impair long-term retention by limiting momentary awareness of unwanted memories: direct suppression and thought substitution 612.
Direct Retrieval Suppression
The first mechanism, direct suppression, disengages episodic retrieval through systemic neural inhibition. When an individual actively suppresses an intrusive memory, there is a pronounced increase in the activation of the right dorsolateral prefrontal cortex (rDLPFC) and the right ventrolateral prefrontal cortex (VLPFC) 3612. These lateral prefrontal regions are functionally analogous to the cortical areas responsible for overriding and stopping physical motor actions, suggesting that memory suppression engages a domain-general inhibitory mechanism to stop retrieval 35.
Crucially, dynamic causal modeling and effective connectivity analyses demonstrate the spatial pathways of this inhibition. The rDLPFC exerts direct top-down inhibitory control over the hippocampus, which is the core medial temporal lobe structure required for conscious episodic recollection 56. During direct suppression, hippocampal activity is significantly reduced below baseline levels 312. Simultaneously, this downward inhibitory signal targets the amygdala, systematically dampening the emotional valence and affective arousal associated with the memory 512. This prefrontal-hippocampal-amygdala pathway provides a concrete biological mechanism for the psychological phenomenon of blocking an unbearable representation from conscious awareness 5.
Thought Substitution
The second mechanism of motivated forgetting is thought substitution. Instead of directly inhibiting the neural trace of the unwanted memory, the individual actively retrieves a substitute memory to occupy the limited focus of conscious awareness 612. Thought substitution engages an entirely different neural network, heavily relying on the left caudal prefrontal cortex and the left inferior frontal gyrus (IFG) 36. Unlike the inhibitory action of direct suppression, thought substitution is characterized by excitatory signals sent from the left IFG to the hippocampus. This results in increased hippocampal activation, as the brain works to sustain the active retrieval processes of the alternative, non-threatening memory, thereby outcompeting the unwanted memory for conscious access 3612.
Trauma, Dissociation, and Repression
The clinical debate regarding how extreme trauma affects memory function relies on differentiating the classical concept of repression from the clinically observed phenomenon of dissociation. Psychoanalytic models suggest that repression is a neurotic defense associated with the left-brain mental-self, which actively buries internal conflict 137. In contrast, dissociation is recognized as a profound psychobiological defense mechanism deployed when physical escape from a traumatic event is impossible, heavily engaging right-brain limbic and somatic networks 138.
The Neurobiology of Trauma and Dissociation
During acute trauma, extreme stress responses dysregulate the hypothalamic-pituitary-adrenal (HPA) axis. The subsequent flood of catecholamines and glucocorticoids heightens amygdala activation while simultaneously inhibiting the retrieval mechanisms of the hippocampus 169. Because the hippocampus is responsible for organizing experiences into coherent, time-bound episodic memories, its inhibition results in the traumatic event being encoded as fragmented sensory or emotional impressions rather than integrated narratives 1618. This explains why traumatized individuals may experience intense somatic flashbacks triggered by environmental cues - such as a specific sound or smell - without a coherent narrative memory of the original event 16.
Neuroimaging studies of patients experiencing dissociative amnesia or dissociative identity disorder reveal specific network alterations. While findings are heterogeneous, resting-state functional connectivity (RSFC) studies consistently identify an altered default mode network (DMN), a hypo-activated right inferolateral prefrontal cortex, and abnormal connectivity between the orbitofrontal cortex and the retrosplenial cortex 1021. Additionally, patients asked to retrieve memories from an amnestic period often fail to activate hippocampal and para-hippocampal structures efficiently 21. In specific clinical contexts, such as the dissociative auras preceding seizures, researchers utilizing intracranial recordings have identified an objective electrophysiological signature: a 3-hertz oscillating signal generated by nerve cells in the posteromedial cortex, which directly correlates with the subjective experience of dissociation and disconnection from reality 11.
The Dissociative Amnesia Debate
Despite these neurobiological correlates, the clinical validity of dissociative amnesia as a mechanism for entirely repressed memories remains heavily contested 2324. Critics point out that many fMRI studies of purported dissociative amnesia rely on individual case reports and fail to rule out organic amnesia or malingering 2324. Furthermore, some researchers argue that the brain activation patterns observed in dissociative amnesia closely mirror the prefrontal activation seen in active, voluntary memory suppression tasks 1226. This overlap suggests that some cases diagnosed as involuntary dissociative amnesia may actually be instances of chronic, motivated forgetting orchestrated by the prefrontal cortex 1226.
Memory Distortion and Source Monitoring Errors
The debate over whether traumatic memories can be literally repressed or if they are subject to reconstructive distortion is colloquially known as the "Memory Wars" 23. Cognitive neuroscience emphasizes the highly reconstructive nature of memory, warning that "recovered" memories are exceedingly susceptible to distortion 2728. Memories are not stored as perfect, immutable video recordings; rather, they are reconstructed from distributed cortical networks during each retrieval event.
Mechanisms of False Memory Generation
False memories frequently occur due to source monitoring errors - cognitive failures to accurately determine the origin of a mental experience 2930. When the brain attempts to recall an event, it utilizes heuristic shortcuts to decide whether an activated representation originated from a real external event, a dream, a story, or internal imagination 30. If the contextual details are fuzzy - often exacerbated by stress, trauma, distraction, or the passage of time - the brain may misattribute internally generated imagery or externally supplied misinformation as a genuine autobiographical memory 2830.
This vulnerability is demonstrated through the Deese-Roediger-McDermott (DRM) paradigm, a standard laboratory procedure where subjects study lists of semantically related words. Participants consistently generate false memories of "critical lures" (words highly associated with the studied list but never actually presented) because they fail to monitor the source of the spreading semantic activation within their neural networks 2729.
Trauma and Suggestibility
Cognitive theories, including the Activation-Monitoring theory and Fuzzy-Trace theory, suggest that stress and trauma can impair the brain's consolidation processes, making individuals more reliant on the "gist" of an event rather than specific item details 2729. Studies have found that individuals who self-report higher levels of trait dissociation and past trauma tend to exhibit lower cognitive ability scores and a higher susceptibility to false memory errors in DRM paradigms 311333. Research by Elizabeth Loftus on the misinformation effect further demonstrates that misleading post-event information can effectively overwrite or distort original memory traces 2728.
However, neurobiological models of genuine trauma-induced dissociation challenge the assertion that all recovered traumatic memories are artifacts of suggestion. Evidence indicates that while peripheral details of a traumatic event may blur, survivors typically recall the core traumatic experience confidently 8. Furthermore, experimental attempts to implant completely false traumatic scenarios in survivors generally fail, suggesting that extreme trauma creates a deeply encoded, albeit fragmented, somatic trace that resists simple verbal misinformation 8.
| Memory Phenomenon | Level of Awareness | Putative Mechanism and Neural Substrates | Clinical and Behavioral Manifestation |
|---|---|---|---|
| Motivated Forgetting (Suppression) | Conscious / Voluntary | Top-down inhibition; right DLPFC and VLPFC actively down-regulating Hippocampus and Amygdala activity 56. | Intentional avoidance of distressing thoughts; successful down-regulation of emotional and autonomic responses 1034. |
| Classical Repression | Unconscious / Involuntary | Theorized to relate to left-brain mental-self operations; empirical neural markers highly overlap with implicit suppression 1335. | Unconscious blocking of unacceptable impulses; memory remains wholly inaccessible without specific clinical intervention 1035. |
| Trauma-Induced Dissociation | Unconscious / Involuntary | Right-brain limbic dominance; HPA axis dysregulation; posteromedial cortex 3Hz rhythmic oscillations 131811. | Feeling of profound detachment, depersonalization, or derealization; fragmented sensory encoding; profound amnesia for trauma 81135. |
| Source Monitoring Error | Unconscious (Believed True) | Breakdown in prefrontal evaluation heuristics; reconstructive failure during hippocampal retrieval 2930. | Confident recollection of events that never occurred; misattribution of imagined details or misinformation to historical reality 3036. |
Predictive Processing and Memory Reconsolidation
To explain how the unconscious processes expectations, synthesizes past trauma, and anticipates the future, modern neuroscience has heavily adopted the predictive coding (or predictive processing) framework 37. This framework provides a rigorous, computational update to the Freudian concept of "unconscious expectations," reframing the brain not as a passive receiver of sensory inputs, but as a proactive inference engine 3738.
The Bayesian Brain and Active Inference
Originating conceptually from Hermann von Helmholtz's nineteenth-century theory of "unconscious inference," predictive coding postulates that the brain continuously generates hierarchical generative models of the environment 3738. These models follow Bayesian brain hypotheses, where higher cortical levels actively predict the flow of sensory data ascending from lower sensory organs 3739. When descending top-down predictions meet ascending bottom-up sensory inputs, the brain calculates the difference.
If the internal prediction is perfectly accurate, the sensory input is effectively "explained away" via descending cortical inhibition, resulting in highly efficient metabolic processing 14. If there is a mismatch, a "prediction error" (or neural "surprise") is generated. This prediction error signal propagates up the cortical hierarchy to force an update to the internal generative model 3741. The framework of active inference extends this concept by proposing that organisms can also minimize prediction errors (or variational free energy) by altering their behavior to actively seek out sensory states that confirm their existing models 4142.
Prediction Error and Memory Destabilization
Predictive coding fundamentally alters the neurobiological understanding of memory storage. In this framework, memory is not a static archival record of the past but a dynamic tool optimized for generating future predictions 4344. The well-documented phenomenon of memory reconsolidation demonstrates that established long-term memories are not permanent. When a memory is reactivated, it enters a temporary, labile state during which it can be altered, strengthened, or disrupted before being re-stabilized (reconsolidated) via new protein synthesis in the hippocampus 4345.
Recent computational and behavioral models demonstrate that memory destabilization is strictly governed by prediction errors 4515. When individuals encounter an "incomplete reminder" - a retrieval cue that partially replicates a past experience but conspicuously omits an expected outcome - it generates a robust prediction error 4547. This surprising mismatch violates the brain's internal model, signaling that the existing memory trace is no longer perfectly adaptive. Consequently, the memory trace is destabilized, engaging hippocampal Zif268 protein expression and proteasome degradation, allowing new, updated information to be integrated into the neural network 4547.
In cases of chronic pain, post-traumatic stress, or phobia, predictive coding explains how rigid, maladaptive internal models are maintained. If an individual has a long history of trauma associated with a specific context, their brain builds an internal model that heavily predicts extreme threat. These top-down predictions of threat can become so heavily weighted (high precision) that they override benign bottom-up sensory data, causing the individual to perceive danger or trigger autonomic stress responses even in objectively safe environments 163914. Healing trauma, within this framework, requires introducing precision-weighted prediction errors - experiences that violate the expectation of harm, forcing the brain to update its hyper-vigilant generative model 3914.
Neurobiological Equivalents of Defense Mechanisms
Sigmund Freud and Anna Freud outlined numerous defense mechanisms - unconscious psychological strategies deployed by the ego to protect the individual from anxiety and internal conflict 248. Early experimental psychology struggled to quantify these highly subjective phenomena. However, modern cognitive and social psychology have identified empirical and neural correlates for several of these classical defenses, while others have been reclassified as standard cognitive biases or generalized physiological responses 49. Assessment tools, such as the Defense Style Questionnaire (DSQ), categorize these into mature, neurotic, and immature defense styles, correlating their overuse with specific psychopathologies 1651.
Empirically Supported Defense Mechanisms
- Reaction Formation: This defense involves converting a socially unacceptable impulse into its behavioral opposite (e.g., displaying exaggerated, disproportionate affection to conceal intense feelings of hatred or jealousy) 5253. Empirical studies in social psychology have amply demonstrated reaction formation in normal populations, particularly regarding self-esteem protection and the suppression of unacceptable social biases 49. Neurophysiologically, this mechanism aligns with the brain's capacity for rapid emotional reappraisal and prefrontal-mediated impulse override, requiring significant cognitive load to maintain the artificial opposite state 17.
- Denial and Isolation of Affect: Denial (the outright refusal to accept a traumatic reality) and isolation of affect (discussing or recalling a traumatic event devoid of any emotional experience) show significant empirical backing 5253. In neuroimaging, these defenses correlate with increased activation in prefrontal executive control networks that actively dampen limbic and amygdala reactivity, effectively severing the cognitive awareness of an event from its autonomic emotional response 1655.
Recontextualized Defense Mechanisms
- Projection: Projection involves attributing one's own unacceptable feelings or traits to another person, effectively externalizing the internal conflict 3548. While prevalent in human behavior, modern psychology often recharacterizes projection less as a targeted ego defense and more as a byproduct of cognitive biases and failed thought suppression. When individuals attempt to suppress thoughts about an undesirable trait in themselves, those specific concepts remain highly active in semantic networks (chronic cognitive accessibility). This priming makes the individual far more likely to perceive and interpret that trait in others, a phenomenon driven by neural activation patterns rather than a direct, targeted defense mechanism 49.
- Displacement: Defined as transferring an emotional reaction from a threatening original source to a less threatening substitute target (e.g., experiencing anger toward an abusive authority figure but expressing it toward a subordinate or spouse) 5356. While emotional carryover is empirically real - largely due to lingering physiological arousal and elevated circulating catecholamine levels - there is minimal evidence that displacement serves a true defensive function of protecting self-esteem or reducing primary anxiety. It is generally viewed as an issue of emotional dysregulation and state-dependent arousal transfer rather than an orchestrated unconscious defense mechanism 49. Sublimation (channeling unacceptable impulses into socially acceptable, productive activities) similarly lacks strong empirical backing as a specific physiological mechanism, often being viewed merely as an artifact of standard goal-oriented behavioral redirection 249.
| Defense Mechanism | Classical Psychoanalytic Definition | Modern Cognitive and Neuroscientific Interpretation | Empirical Support Level |
|---|---|---|---|
| Reaction Formation | Expressing the exact opposite of unacceptable inner impulses or drives 5253. | Active suppression of a primary impulse coupled with deliberate over-expression of a socially acceptable alternative 49. | High. Consistently observed in self-esteem protection and prejudice suppression paradigms 4957. |
| Denial | Blocking external traumatic events or stressors from conscious awareness 4852. | An extreme form of motivated forgetting; prefrontal down-regulation of sensory and emotional salience 5253. | High. Observable as a standard psychological coping strategy in clinical populations 4955. |
| Projection | Externalizing undesirable aspects of the self onto other people 3548. | Cognitive bias resulting from failed thought suppression; hyper-accessibility of suppressed concepts in semantic networks 3549. | Moderate. Exists as a behavioral phenomenon, but its primary function as a pure defense mechanism is debated 49. |
| Displacement | Redirecting a dangerous emotion to a safer, subordinate target 256. | Residual physiological arousal and hormone circulation transferring to new contexts; lacks specific defensive anxiety-reduction 49. | Low. Observed as simple emotion-transfer and arousal spillover, but not supported as a targeted psychological defense 49. |
Cross-Cultural Variances in Neurodevelopment and Trauma
A critical limitation in early psychoanalytic and neuroscientific models of memory, the unconscious, and trauma was an overwhelming over-reliance on WEIRD (Western, Educated, Industrialized, Rich, and Democratic) populations. Modern cross-cultural neuroscience reveals that human information processing is not entirely universal; rather, it is fundamentally shaped by culturally derived self-representations, affecting memory encoding styles, structural brain maturation timelines, and trauma responses 5859.
Cultural Differences in Autobiographical Memory
Behavioral and neuroimaging studies consistently demonstrate that individuals from Western (individualistic) cultures exhibit higher levels of autobiographical memory specificity than those from East Asian (collectivistic) cultures 5860. American adults tend to recount memories focused heavily on the autonomous self, personal achievements, and specific internal emotional states. Conversely, Chinese adults recount memories focused predominantly on social relationships, historical context, and group dynamics 5818.
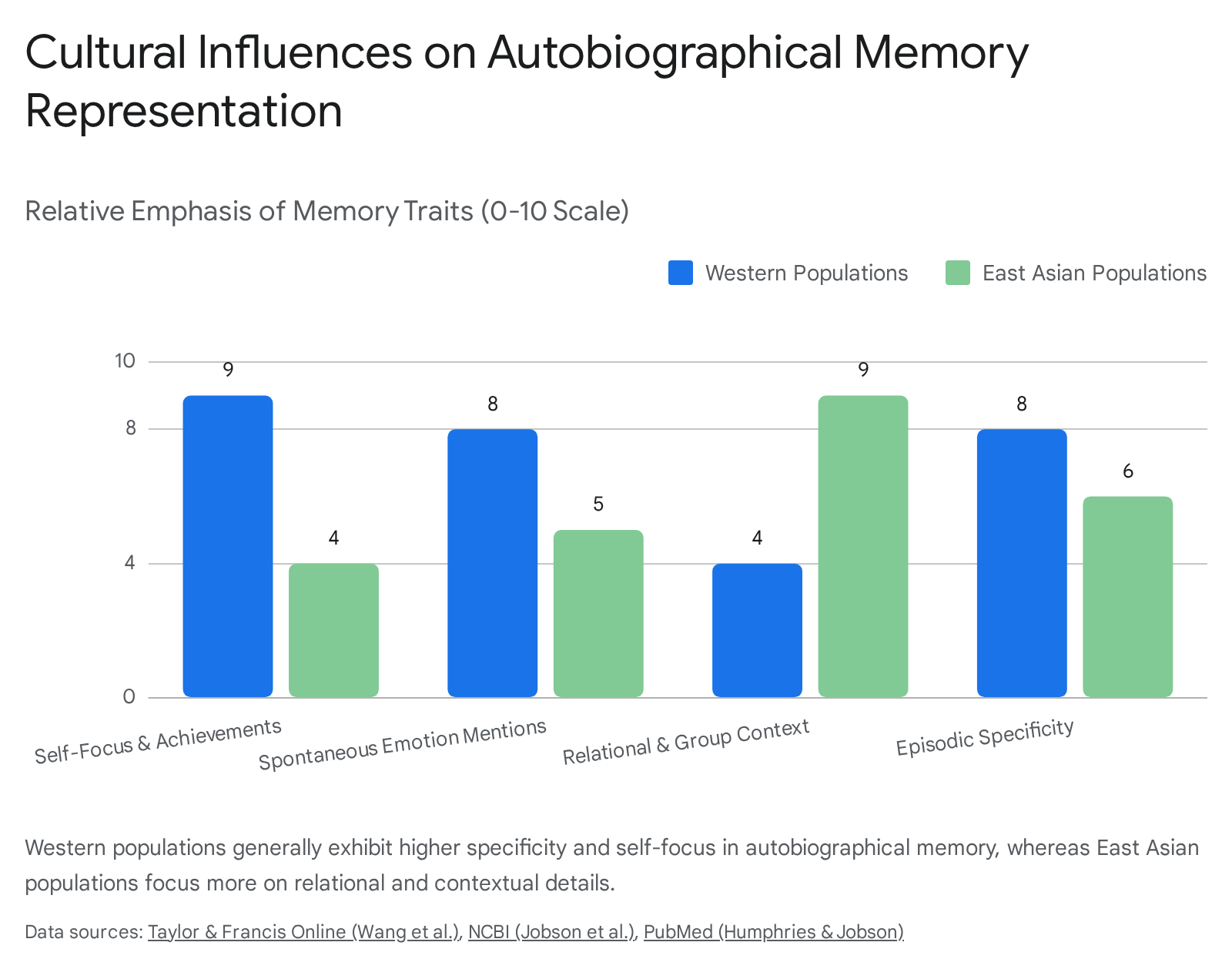
These distinct psychological focuses are directly reflected in functional neuroanatomy during retrieval tasks. During object recognition and mnemonic discrimination tasks, functional MRI reveals that American and Taiwanese individuals diverge significantly in their neural activity. Americans demonstrate distinct activation patterns in the left fusiform gyrus, superior parietal cortex, and hippocampus when distinguishing novel objects from previously seen lures. This indicates a greater reliance on visual processing and attention-modulating regions to isolate specific, individual object features 60. In older populations, these cultural memory differences become even more pronounced, with Taiwanese older adults showing larger age-related declines in highly specific object discrimination compared to American counterparts 18.
Structural Brain Maturation Timelines
Beyond functional activation, recent large-scale, population-specific brain charting initiatives (such as the Chinese Lifespan Brain Mapping Consortium, which analyzed neuroimaging data from over 43,000 participants) reveal that foundational neurodevelopmental trajectories are not entirely universal 6263. When comparing Chinese structural neuroimaging data to Western normative charts, researchers identified distinct spatiotemporal maturation timelines. Chinese populations exhibit prolonged cortical and subcortical maturation, highly accelerated cerebellar growth peaking in adolescence, and a later development of higher-order association cortices relative to primary visual cortices 6263. These architectural findings underscore that standard psychiatric models of brain function, memory processing, and cognitive development must be calibrated to specific genomic, environmental, and cultural interactions rather than assuming a single global baseline 63.
Pan-Cultural Effects of Severe Trauma
Despite significant baseline cultural variances in daily memory processing and brain maturation, the fundamental neurobiological impact of severe trauma appears highly conserved across populations. When individuals from highly diverse cultures - including Australian, British, Iranian, and Chinese cohorts - develop Post-Traumatic Stress Disorder (PTSD), they exhibit universally similar deficits and distortions in global autobiographical remembering 6465. Specifically, individuals with PTSD across all tested cultures show significantly fewer expressions of autonomy, self-determination, and episodic specificity in their memory narratives compared to non-traumatized controls from their own cultures 6465. Thus, while the phenomenological expression and focus of an episodic memory is culturally moderated, the disruption of the memory encoding system by severe hyper-arousal and fear-circuitry dysfunction remains a pan-cultural biological reality 59.
Conclusions
Modern neuroscience indicates that Sigmund Freud was largely incorrect about the specific architecture and intent of the unconscious. There is no isolated, shadow-mind governed by primitive sexual and aggressive drives operating under entirely different rules from conscious thought. Furthermore, the classical psychoanalytic concept of pristine memory repression - where a highly complex traumatic event is perfectly walled off from the mind but remains historically intact and fully recoverable in its original form - is heavily contested by modern understandings of reconstructive memory, source monitoring errors, and the fragmented nature of trauma-induced dissociation.
However, Freud was profoundly correct in his overarching, revolutionary premise: the vast majority of human cognition, emotional regulation, and behavioral motivation occurs completely outside of conscious awareness. Modern neurobiology validates the existence of sophisticated defense mechanisms, confirming that the brain actively deploys top-down prefrontal inhibition to exclude painful memories from awareness through motivated forgetting. Furthermore, the modern predictive processing framework validates the concept that deeply held, non-conscious expectations actively shape, filter, and sometimes distort our perception of reality. By mapping these complex psychological phenomena to specific, observable neural circuits and acknowledging cross-cultural developmental variations, modern neuroscience has not erased the unconscious; rather, it has integrated it into a unified, biologically grounded model of the human mind.