Neural mechanisms of memory encoding and reconstructive retrieval
Introduction: Dismantling the Epistemological Myth of the Playback Model
For the better part of a century, the public understanding of memory - and indeed, much of early psychological theorizing - has been dominated by an intuitive but fundamentally flawed epistemological metaphor: the brain functioning as a video camera or a computer hard drive. In this persistent misconception, everyday experiences are purportedly recorded as immutable files, stored in a static cognitive archive, and later played back with objective, high-definition fidelity during recall 11. When errors in memory occur, they have traditionally been attributed to the progressive "decay" of the recording medium over time or a mechanical failure of the retrieval process, rather than a fundamental alteration of the stored data itself 12. However, modern cognitive psychology and high-resolution neuroscience have thoroughly dismantled this playback model. The overwhelming scientific consensus now recognizes that memory is not a passive repository but an active, dynamic, and highly reconstructive biological process 124.
The initial paradigm shift away from static storage was driven by a triad of foundational classic researchers. Eric Kandel's Nobel-winning research on the marine mollusk Aplysia californica demonstrated that associative learning and memory involve structural, synaptic plasticity and anatomical changes at the neuronal level, rather than mere energetic or electrical reverberation 3. Subsequently, Karim Nader's pioneering work on fear conditioning revealed the phenomenon of memory reconsolidation, proving that the act of recalling a memory actually returns the neural trace to a highly labile state, requiring de novo protein synthesis to be maintained and stored once again 4. Parallel to these biological discoveries, Elizabeth Loftus's extensive behavioral experiments on the misinformation effect proved that human episodic memories can be wholly fabricated or severely distorted by post-event suggestions, definitively establishing the fallibility of eyewitness testimony 15.
Building upon these historical pillars, research conducted from 2023 onward has accelerated the field into an unprecedented era of mechanical precision. Through advanced optogenetic engram cell tracking, high-resolution functional magnetic resonance imaging (fMRI), and sophisticated molecular profiling, contemporary neuroscience has mapped the exact mechanisms that govern how memories are encoded at event boundaries, destabilized during recall, integrated across networks, and physically pruned by the immune system during sleep 678. This exhaustive research report investigates these recent advancements. It maps the complete neurobiological lifecycle of memory, from the subcellular dependencies of consolidation to the systems-level orchestration of the neocortex. Furthermore, it addresses the limitations of translating simple animal models to human narrative complexity, explores how geographically diverse cultural structures shape autobiographical memory, and outlines the profound practical implications of memory malleability for modern legal standards and psychiatric exposure therapies.
The Reconstructive Framework and Schema Theory
The cognitive architecture of human episodic memory fundamentally rejects the notion of verbatim playback. When an individual perceives an event, the brain does not passively copy the external world; rather, it constructs a representation by binding together highly distributed sensory, emotional, and spatial inputs into a fragile, transient network 12. Consequently, remembering an original experience requires a subsequent reconstruction of those distributed neural traces.
This reconstructive effort is primarily driven by schemas - pre-existing cognitive frameworks, initially conceptualized by Sir Frederick Bartlett in the 1930s, that help individuals categorize, interpret, and navigate the world 14. During retrieval, the brain utilizes these schemas to fill in inevitable gaps in the memory trace, relying on general semantic knowledge, logical inferences, and current beliefs to create a narrative that is coherent, if not strictly accurate 112. While this makes human memory highly efficient and adaptive for predicting future scenarios, it inherently introduces the risk of profound errors 2. Because a memory is essentially constructed anew during each instance of recall, it is highly vulnerable to systemic confabulation, confirmation bias, and external contamination from leading questions 12.
Modern neuroimaging indicates that this reconstruction process relies heavily on the default mode network (DMN), an interconnected system of brain regions that becomes highly active during introspection, mental time travel, and wandering thought 13. The DMN integrates conceptual and emotional details with the perceptual fragments stored in the hippocampus 13. When the brain retrieves a memory, it relies on the DMN to apply an overarching meaning to the sensory fragments, fundamentally altering how the memory is re-encoded for future use.
To clearly delineate the paradigm shift that has occurred within cognitive psychology, the following table contrasts the outdated assumptions of the playback model against the modern, evidence-based reconstructive framework.
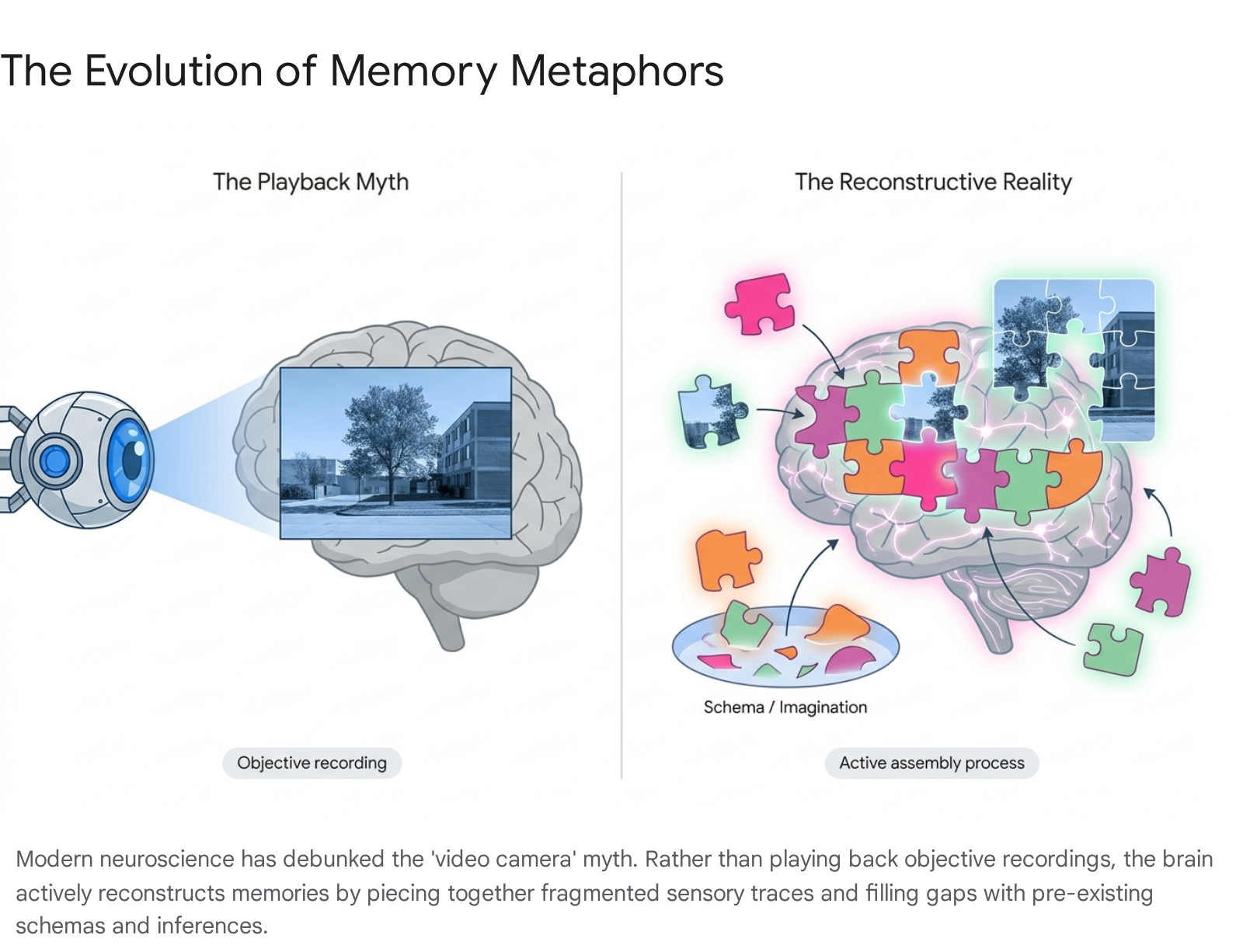
Table 1: The Outdated Playback Model vs. Modern Reconstructive Evidence
| Architectural Feature | Outdated Playback Model (The "Video Camera" Myth) | Modern Reconstructive Model (Evidence-Based Framework) |
|---|---|---|
| Mechanism of Encoding | Passive, continuous recording of all sensory inputs as a unified stream of objective reality. | Active construction; the brain encodes fragmented, highly salient details heavily filtered by attention and existing schemas 24. |
| Storage Architecture | Memories are stored as permanent, intact, localized "files" or snapshots in a specific neural hard drive. | Memories exist as distributed neural ensembles (engrams) across the cortex, requiring continual spatiotemporal binding to maintain cohesion 2. |
| Process of Retrieval | Literal, objective playback or readout of the original, unedited event without alteration. | Imaginative reconstruction; the brain actively pieces together fragments, inferring and fabricating missing details based on the current context 1112. |
| Nature of Memory Errors | Errors are viewed solely as "forgetting" caused by the decay of the storage medium over time. | Errors include confabulation, the misinformation effect, and false memories introduced actively during the reconstruction process 12. |
| Impact of Recall Activity | Recalling a memory is a read-only process that does not alter its fundamental stored state in the brain. | Recalling a memory destabilizes the engram, rendering it chemically labile and open to editing before it must be reconsolidated 414. |
| Primary Evolutionary Function | To maintain an impeccably accurate, historical archive of the past for retrospective analysis. | To utilize past experiences adaptively to simulate, predict, and successfully navigate future evolutionary scenarios 91011. |
Subcellular Mechanics: The Dichotomy of Consolidation and Reconsolidation
At the molecular and subcellular level, the persistence of memory is dictated by specific signaling pathways that govern synaptic plasticity. When a novel experience occurs, the resulting memory trace is initially highly labile and susceptible to disruption from pharmacological or behavioral interference 1213. To become a stable long-term memory, the trace must undergo a process called consolidation, a time-dependent mechanism that stabilizes the morphological changes at the synapse through de novo protein synthesis and gene transcription 121314.
However, the act of retrieving an established long-term memory returns it to a transiently labile state 121516. This phenomenon, known as reconsolidation, is not merely a repetition of the initial consolidation event, but rather an entirely distinct biological cycle composed of two phases: destabilization and restabilization 12.
The destabilization phase is fundamentally a protein degradation-dependent process. It is initiated by calcium influx through N-methyl-D-aspartate receptors (NMDARs) and L-type voltage-gated calcium channels (LVGCCs) 162317. This calcium signaling triggers the ubiquitin-proteasome system (UPS) to actively degrade pre-existing post-synaptic scaffold proteins, essentially unmooring the memory from its stable synaptic connections 2325. If this destabilization process is pharmacologically blocked, the memory trace is never rendered labile during retrieval; it remains impervious to modification and persists in its original state, failing to incorporate any new information present in the environment 1617. Current computational models and neurobiological studies from 2024 and 2025 also suggest that homeostatic plasticity and synaptic downscaling may act as primary drivers for this destabilization, balancing the Hebbian plasticity that originally formed the memory 2526.
Once a memory is successfully destabilized, it enters the restabilization phase, which is required to place the updated trace back into long-term storage. Like initial consolidation, restabilization is heavily reliant on new protein synthesis, regulated by the mammalian target of rapamycin (mTOR) pathway, which facilitates local protein translation within dendritic spines 12.
Despite sharing the overarching requirement for protein synthesis, consolidation and reconsolidation exhibit distinct molecular dependencies and boundary conditions. Foundational research established a critical double dissociation: the expression of Brain-Derived Neurotrophic Factor (BDNF) is strictly required for initial consolidation but is entirely dispensable for reconsolidation 1215. Conversely, the transcription factor Zif268 (also known as Egr-1) is absolutely necessary for the reconsolidation of updated memories but plays no vital role in initial consolidation 1215. Furthermore, highly recent neurobiological studies (2024 - 2025) have implicated the atypical protein kinase C isoform, PKM$\zeta$, specifically in the reconsolidation of spatial memories within the hippocampus 18. Administration of the zeta-inhibitory peptide (ZIP) following memory retrieval induces long-lasting amnesia for the reactivated spatial trace, yet ZIP has no effect on the passive maintenance of the memory if the trace is never retrieved, highlighting its specific role in the reconsolidation window 18.
The initiation of reconsolidation is heavily constrained by "boundary conditions" such as memory age, initial training strength, and the presence of a prediction error 121619. For instance, extremely robust or highly aged memories are often resistant to destabilization 1219. However, modern pharmacological interventions are exploring ways to artificially induce destabilization in these stubborn traces by manipulating NMDARs prior to retrieval, thereby overriding the natural boundary conditions 19.
Table 2: Comparative Molecular Architecture of Consolidation vs. Reconsolidation
| Biological Parameter | Initial Memory Consolidation | Memory Reconsolidation (Updating) |
|---|---|---|
| Triggering Event | The novel encoding of a stimulus or unpredicted experience 13. | The targeted retrieval or reactivation of a previously stored long-term memory 12. |
| Destabilization Phase | Not applicable; the structural trace is being formed entirely de novo. | Strictly required. Actively mediated by NMDARs, LVGCCs, CB1 receptors, and UPS-driven protein degradation 1617. |
| Primary Transcription Factors | CREB, NF-$\kappa$B, C/EBP$\beta$ 121415. | CREB, NF-$\kappa$B, Zif268 (uniquely specific to the reconsolidation pathway) 1215. |
| Specific Molecular Markers | Heavily dependent on BDNF expression 12. | Heavily dependent on PKM$\zeta$ activity and Ca2+-permeable AMPA receptors 1618. |
| Boundary Conditions | Dependent primarily on the intensity, emotion, and salience of the initial encoding event. | Heavily constrained by memory age, memory strength, and the absolute requirement of prediction errors (novel information) 121619. |
| Psychological Outcome | The transformation of fragile short-term memory (STM) into a stable long-term memory (LTM) 13. | The dynamic integration of new data; resulting in the strengthening, weakening, or qualitative alteration of the existing LTM 1215. |
Tracking the Engram: Optogenetics and High-Resolution Synaptic Imaging
For much of the 20th century, the "engram" - the physical, localized neural substrate of a specific memory - was considered a theoretical abstraction. However, recent advancements in activity-dependent genetic tagging and optogenetics have transitioned the engram from a concept into an observable, manipulable reality 623. Research published through 2023 and 2024 has enabled scientists to isolate, track, and artificially control the sparse, distributed ensembles of neurons that fire simultaneously during a specific learning episode.
In modern engram tracking, excitatory neurons that are active during a training event are genetically tagged using precise temporal systems. For example, recent studies utilize the scFLARE2 activity-dependent tagging system to examine memory specificity 6. To gain causal control over these specific cells, researchers microinject viral vectors, such as the herpes simplex virus vector HSV-NpACY, into targeted brain regions like the lateral nucleus of the amygdala 6. This viral delivery allows for the expression of spectrally compatible opsins exclusively within the allocated engram ensemble. Specifically, the vectors express Channelrhodopsin-2 (ChR2), an excitatory opsin sensitive to blue light, and eNpHR3.0, an inhibitory halorhodopsin sensitive to red light 6.
The behavioral results of these optogenetic interventions are profound and unambiguous. By exposing the tagged engram neurons to precise pulses of blue light (e.g., 473 nm, 20 Hz, 5 ms pulse width), researchers can induce artificial memory recall in a novel context, entirely in the absence of the original sensory retrieval cues 6. Rodents undergoing artificial optogenetic recall exhibit behavioral profiles that are strikingly identical to those experiencing natural retrieval, including high levels of defensive freezing and matching ultrasonic vocalization (USV) densities in the 20 - 30 kHz range 6. Conversely, illuminating the engram with red light (660 nm) effectively silences the targeted neurons, reliably disrupting natural memory retrieval and inducing temporary amnesia 6.
Beyond optogenetics, 2025 research published in Nature Neuroscience by Harvard University teams has unveiled the Extracellular Protein Surface Labeling in Neurons (EPSILON) technique 20. This groundbreaking methodology allows for the high-resolution mapping of AMPA receptors (AMPARs) - proteins absolutely vital for the transmission of signals across synaptic connections during plasticity 20. Utilizing sequential labeling with specialized fluorescent dyes, EPSILON enables researchers to monitor the physical movement and behavior of these receptors in real-time, providing an unprecedented lens into the synaptic architecture of memory formation and revealing the rules governing how the brain decides which specific synapses to strengthen or weaken 20.
Systems-Level Dynamics: Event Boundaries and fMRI Retrieval Networks
While optogenetics illuminates the behavior of cellular engrams in isolated regions, high-resolution functional MRI studies from 2024 and 2025 have mapped the broader systems-level retrieval networks, fundamentally altering our understanding of how narrative memories are segmented, encoded, and retrieved 821.
A central computational challenge for the human brain is understanding how it processes a continuous stream of sensory input into discrete, manageable episodic events. Recent neuroimaging data demonstrates that the brain does not record experiences continuously like a rolling tape; instead, it relies heavily on "event boundaries." These boundaries are moments of high uncertainty, prediction error, or sudden shifts in spatial or narrative context 312233.
A paradigm-shifting 2024 study by Ranganath and colleagues, published in the high-impact journal Neuron, investigated hippocampal-cortical interactions during the encoding of complex naturalistic narratives (such as cartoon movies) 8. The fMRI study revealed that functional connectivity between the hippocampus and the neocortex - specifically the Posterior Medial Network (PMN) - is highly dynamic and heavily biased toward the offset of an event 831. The researchers found that heightened functional connectivity between the hippocampus and the PMN precisely at an event boundary is highly predictive of subsequent recall success, as well as the granularity of detail retrieved after a two-day delay 831. Conversely, high hippocampal-PMN connectivity during the mid-point of a continuous event is paradoxically associated with poorer memory retention 31. This confirms that the hippocampus acts as a complementary learning system, taking critical "snapshots" of neocortical activity at moments of transition, rather than recording the entirety of the experience 22.
Furthermore, traditional cognitive theories assumed a strict neurological dichotomy between episodic memory (the recall of personal, contextual life experiences) and semantic memory (the recall of general, detached factual knowledge). However, a breakthrough 2026 study conducted by the University of Nottingham and the University of Cambridge, published in Nature Human Behaviour, entirely debunked this division 2324. Utilizing task-based experiments combined with fMRI, the team revealed no measurable difference in brain activity between successful semantic and episodic retrieval 2324. The brain activates vastly overlapping neural networks - including the medial temporal lobe (MTL) convergence hubs, the prefrontal cortex control networks, and the parietal precuneus - regardless of whether a subject is remembering a specific life event or recalling a general fact 2123. This data reinforces the reconstructive theory at a systems level: all memory retrieval relies on a centralized, holistic network that actively binds distributed fragments into conscious awareness, rather than utilizing separate anatomical filing cabinets for different types of data 2324.
The Architecture of Sleep: Consolidation and Microglial Synaptic Pruning
The stabilization, refinement, and organization of these complex neural networks are highly dependent on the physiological mechanisms of sleep. While sleep was once viewed as a passive state of rest, research from 2023 and 2024 highlights that the two primary phases of sleep - Non-Rapid Eye Movement (NREM) and Rapid Eye Movement (REM) - serve highly active, complementary functions in memory consolidation and synaptic pruning 25.
NREM Sleep: Network Stabilization and Coupling
NREM sleep, particularly the deep slow-wave stages, acts as the primary driver for memory stabilization. This phase is characterized by the intricate temporal coupling of cortical slow-oscillations, thalamic sleep spindles, and hippocampal sharp-wave ripples 25. During NREM, dynamic cortical-hippocampal coupling facilitates the persistent, synchronized reactivation of newly formed memory traces 1325. These spindle trains effectively migrate the fragile, hippocampus-dependent episodic codes to more permanent, distributed neocortical storage areas 1325. The neuromodulator norepinephrine (NE) plays a vital role during NREM, orchestrating the selective downscaling of highly active neurons to maintain overall homeostatic balance while preserving the relatively potentiated memory trace 25.
REM Sleep: Abstraction and Immune-Mediated Pruning
As the brain transitions into REM sleep, dominated by theta oscillations and the release of dopamine (DA), the mnemonic focus shifts away from basic stabilization toward integration and abstraction 25. REM sleep enables the prefrontal cortex to process inferred knowledge, fitting newly acquired episodic memories into broader semantic frameworks to support cognitive flexibility and creativity 25.
Crucially, modern research (2024 - 2025) has identified REM sleep as a vital biological window for structural network refinement via microglia-mediated synaptic pruning 725. Microglia, the resident macrophage immune cells of the central nervous system, actively patrol the brain, physically engulfing and digesting less active or redundant synapses, thereby refining the memory network to reduce noise and enhance signal efficiency 72526.
This microglial phagocytosis is strictly regulated by highly specific molecular signaling cascades. Weak or immature synapses display "eat-me" signals, such as externalized phosphatidylserine (PS) and components of the innate immune system's complement cascade, primarily the proteins C1q, C3, and its cleavage fragment C3b 72627. The microglia utilize specific surface receptors - including CR3, MERTK, and GPR56 - to recognize these tags and initiate engulfment of the synapse 72639.
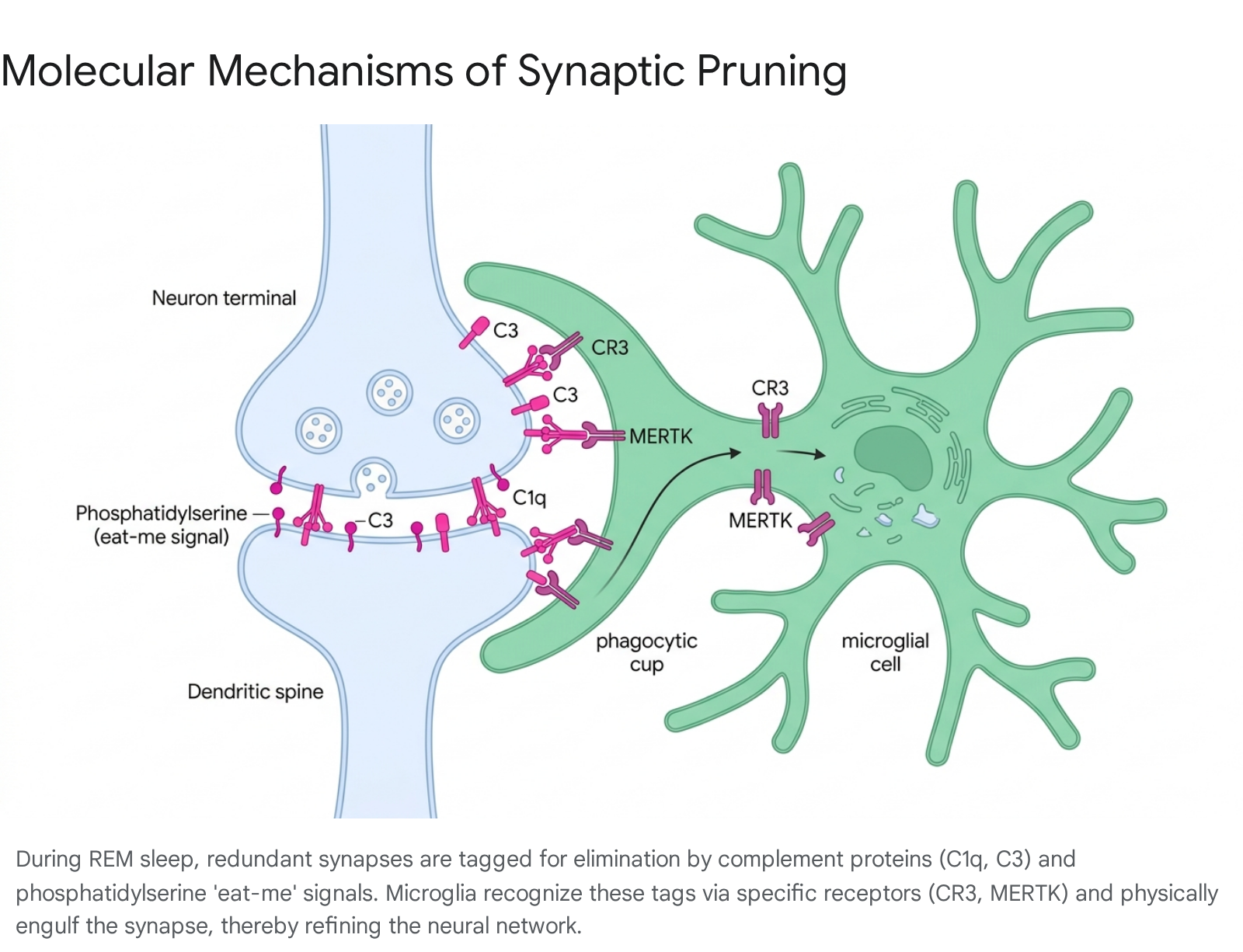
The process is driven by chemokine signaling, particularly the CX3CL1/CX3CR1 axis, which facilitates communication between neurons and microglia 27. Conversely, robust, task-relevant synapses are protected by "don't eat-me" signals, such as the classical complement pathway inhibitor SRPX2 and the CD47-SIRP$\alpha$ complex, which actively block microglial processes 726.
Pathological disruptions to this delicate balance can be catastrophic; for instance, 2025 research indicates that in conditions like Post-Traumatic Stress Disorder (PTSD), an excess of the neurotransmitter GABA produced by astrocytes (support cells) impairs the brain's ability to prune and extinguish fear memories in the medial prefrontal cortex 28. Thus, sleep is fundamentally a biological crucible that chemically and immunologically sculpts the physical architecture of the engram.
The Translational Gap: Animal Models vs. Human Narrative Complexity
While the use of simple animal models, such as optogenetics in rodents and conditioning in Aplysia, has been instrumental in mapping the precise molecular mechanisms of synaptic plasticity, translating these reductionist findings to the sprawling complexity of human episodic memory remains a subject of intense scientific debate 32930.
The primary epistemological limitation of animal models lies in the concept of "autonoesis." First articulated by Endel Tulving, autonoesis refers to the uniquely human capacity to consciously project oneself backward in time, re-experiencing an event from a subjective, first-person perspective 329. Because autonoetic consciousness can only be assessed through linguistic introspection and verbal self-reporting, it presents an insurmountable methodological challenge; it cannot be definitively proven or measured in non-human animals that lack language 29.
Consequently, behavioral neuroscientists must rely on experimental proxies. The most common is the measurement of "What-Where-When" (W-W-W) memory, frequently referred to as "episodic-like" memory 2930. In typical rodent paradigms, such as the object-place-context recognition (OPCR) task, researchers rely on a rat's innate behavioral preference for novelty. By observing exploration times, scientists infer whether the animal remembers the spatial location of a single object or reward 29.
While these models successfully demonstrate basic spatiotemporal binding, critics argue they fail entirely to capture the true narrative complexity of human memory 2930. Humans encode vast, sequentially structured narratives heavily inflected by linguistic meaning, deep cultural contexts, and an overarching, continuous concept of selfhood 3144. Furthermore, differentiating between distinct retrieval processes - such as spontaneous recall versus strategic, effortful recall - is highly complex in animal models, whereas humans can simply be instructed to freely recall a verbal list 29. The developmental timelines also differ drastically; human W-W-W memory begins to functionally emerge around age 2 and matures slowly into the school years, a trajectory that is difficult to map onto rodent ontogeny 2930. Therefore, while animal models accurately reflect the molecular mechanics of the engram, they are largely insufficient for studying how episodic traces are linguistically organized into cohesive autobiographical stories.
Cross-Cultural Narratology: The Architecture of Autobiographical Memory
The manner in which human beings structure their episodic memories into autobiographical narratives is not a universal biological constant; it is heavily mediated by cultural life scripts and geographic environments 313233. Recent comparative studies from 2024 and 2025 exploring cross-cultural narratology demonstrate that cultural frameworks dictate what specific information is deemed salient during initial encoding, and how that information is organized during the reconstructive process of recall 4748.
Linear vs. Non-Linear Narrative Structures
In Western societies, which generally foster an "independent" or individualistic view of the self, autobiographical memory relies heavily on linear, chronological structures 324749. Storytelling and memory reconstruction in these cultures often mimic a classical three-act structure centered on direct conflict, individual achievement, overcoming obstacles, and personal growth 4750. Consequently, when Western individuals are prompted to recall their past, they tend to retrieve highly specific, detailed episodic memories focusing heavily on their own autonomous actions and personal agency 4734.
Conversely, in many East Asian cultures, which prioritize an "interdependent" or collectivistic self, autobiographical reconstruction often employs non-linear, cyclical, or thematic organization 3247. For instance, the traditional Kishōtenketsu narrative structure - common in Japanese storytelling - emphasizes harmony, shifting perspectives, and the absence of direct conflict 50. Autobiographical recall in these cultures frequently yields more generalized, mosaic-like memories that highlight social roles, communal relationships, and relational contexts over highly specific, ego-driven individual actions 474934.
Global Storytelling and Identity Formation
These profound structural differences extend globally, demonstrating how the brain adapts its reconstructive schemas to local environments. Native American storytelling often features non-linear fragments that might start at the end of an event, aimed at reinforcing community bonds and moral connection to nature; in these narratives, the protagonist's identity frequently remains static rather than undergoing Western-style "character arcs" 50. In African traditions, narratives emphasize active audience participation and straightforward moral resolutions, while Middle Eastern frame stories (stories nested within stories) are used to explore complex, layered philosophical themes 50.
Furthermore, studies utilizing populations in the Global South (e.g., South America and Africa) show that culturally expected timelines - known as life scripts - heavily dictate the "reminiscence bump," the period of life from which an individual recalls the most memories 3348. For example, research on the Nunggubuyu people of Australia revealed multiple reminiscence bumps specifically linked to community teaching roles in later adulthood, rather than the single, early-adulthood bump typically observed in Western populations 48. Similarly, studies comparing sexual minority parents in Brazil and the United Kingdom found that the presence of minority stress events significantly altered the timing and content of their reminiscence bumps compared to heterosexual peers 33. Ultimately, memory reconstruction is fundamentally a cultural act: the brain utilizes the prevailing cultural blueprint to architect the narrative of the self.
Practical Implications I: Shifting Legal Standards for Eyewitness Testimony
Understanding that memory is a malleable, reconstructive, and highly susceptible biological process has triggered profound paradigm shifts in the justice system, an arena that historically treated eyewitness testimony as empirical, incontrovertible evidence akin to a video recording. The growing recognition of memory reconsolidation and the misinformation effect is forcing systemic judicial reform globally.
A landmark 2024 survey of global memory experts, led by Dr. Travis Seale-Carlisle at the University of Aberdeen, revealed a massive shift in scientific consensus regarding the utility of eyewitness confidence 35. In a previous survey conducted in 2001, nearly 90% of experts believed that an eyewitness's expressed confidence had little to no relationship to their actual accuracy 35. However, by 2024, the consensus entirely flipped: over 90% of memory experts now agree that high confidence is strongly predictive of accuracy, but strictly under one critical condition: the confidence statement must be collected at the initial identification attempt 35.
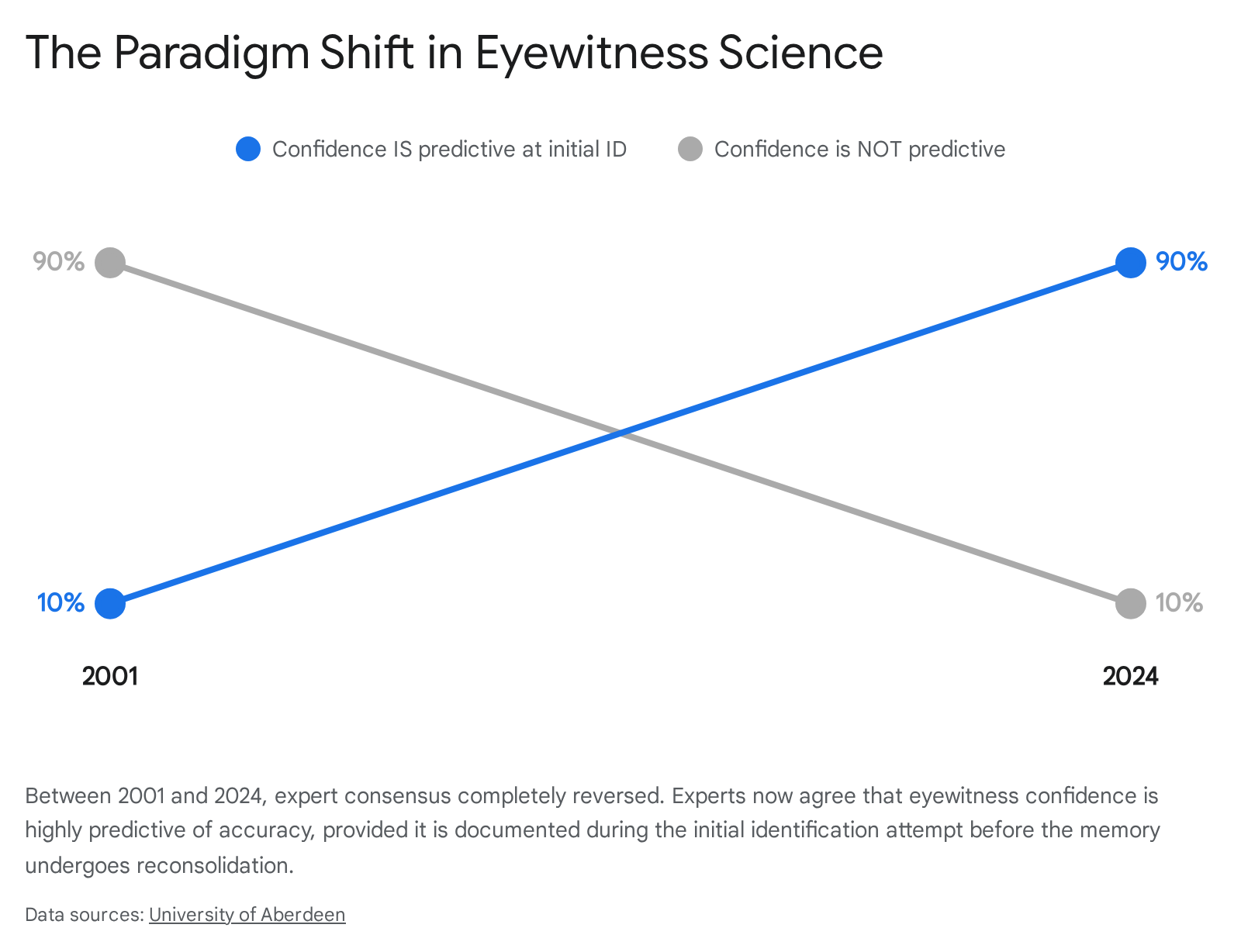
Once a witness is questioned multiple times, the memory enters the reconsolidation window, becoming contaminated by post-event information, leading to artificially inflated confidence at trial months later 35. Despite this overwhelming consensus, policies in England, Wales, and Scotland do not currently mandate the collection of confidence statements during initial video lineups, highlighting a dangerous lag between scientific data and legal practice 35.
Simultaneously, major courts are enacting new precedents to protect against memory failure. The New York Court of Appeals issued four critical rulings in 2024 (Vaughan, Williams, Mosley, and Watkins) that significantly modernized how courts handle visual and memory evidence 53. The Vaughan ruling established that expert testimony regarding the science of memory identification should be accessible to juries regardless of the presence of other corroborating evidence 53. The Mosley ruling mandated strict two-part tests and limiting instructions before police officers are permitted to identify defendants from video footage as non-eyewitnesses, ensuring jurors understand that human memory and perception of video is opinion, not objective fact 53.
Internationally, the documentation of memory and atrocities is adapting to strict digital standards. In 2024, the eyeWitness to Atrocities application enabled human rights defenders to capture over 13,641 secure, tamper-proof photos and videos in conflict zones, ensuring the evidence meets the strict authentication standards of mechanisms like the International Criminal Court (ICC), bypassing the reliance on highly fallible human trauma recall 54. Furthermore, frameworks like the Hague Convention and newly updated 2024 post-Brexit guidelines in the UK are formalizing how remote video testimony is collected across borders, ensuring that the integrity of the witness's memory is preserved and that participation is voluntary and uncoerced by local jurisdictions 5556.
Practical Implications II: Evolving Exposure Therapies for PTSD
In the realm of clinical psychiatry, the recognition of the memory reconsolidation boundary conditions is being actively exploited to treat severe trauma, Post-Traumatic Stress Disorder (PTSD), and debilitating phobias. Because intense traumatic memories are often stubbornly resistant to standard extinction therapy, clinicians are increasingly utilizing the Memory Reconsolidation Updating Procedure (MRUP) 3637.
In MRUP, a patient is asked to briefly retrieve the traumatic narrative to intentionally trigger the molecular destabilization phase. Once the memory trace is chemically labile, pharmacological agents are administered to block the restabilization process. For example, administering Propranolol (a lipophilic beta-blocker) during this critical 1-to-2-hour window blocks noradrenergic signaling in the amygdala 1659. This strips the memory of its visceral, panic-inducing emotional valence while leaving the declarative factual narrative intact 1659. Recent 2025 clinical trials have even expanded this reconsolidation therapy to treat chronic nociplastic low-back pain, attempting to reverse plastic changes in the limbic system that mirror PTSD pathology 59.
Furthermore, breakthrough psychedelic-assisted therapies are redefining trauma treatment by drastically enhancing neuroplasticity and altering how memory pathways are consolidated. A 2024 meta-analysis in Clinical Neuropsychiatry revealed that intravenous Ketamine infusions provide rapid-onset relief (often within 24 hours) for chronic PTSD 60. As an NMDA receptor antagonist, Ketamine blocks glutamate regulation, leading to a massive increase in synaptic plasticity and the release of BDNF in the hippocampus and prefrontal cortex 60. This allows the brain to rapidly rewire trauma associations. Expanding on this, a massive 2025 real-world study by Mindbloom of at-home ketamine therapy reported that 79% of PTSD patients responded to the treatment, with 60% achieving full remission, radically outpacing the efficacy of traditional SSRI antidepressants 61.
Similarly, Phase 3 clinical trials published in 2025 utilizing MDMA-assisted psychotherapy achieved an unprecedented 71% long-term remission rate in patients with severe, treatment-resistant PTSD 62. MDMA promotes the massive release of serotonin, dopamine, and oxytocin, which profoundly decreases amygdala hyperactivity 62. This unique pharmacological state allows patients to actively retrieve, explore, and reconsolidate highly traumatic memories without triggering the overwhelming fear responses that typically stall traditional talk therapy 62.
Finally, cutting-edge drug development is targeting the subcellular environment of the memory trace itself. In August 2025, researchers at the Institute for Basic Science discovered that the failure to extinguish fear memories in PTSD is driven by an excess of the neurotransmitter GABA produced not by neurons, but by astrocytes (star-shaped support cells) in the medial prefrontal cortex 28. The researchers identified a new drug, KDS2010 (an MAOB inhibitor), which selectively blocks this abnormal GABA production 28. Already in human trials, KDS2010 rapidly restored cerebral blood flow and re-enabled fear extinction mechanisms in models, offering a highly targeted molecular "eraser" for the emotional weight of maladaptive memories 28.
Conclusion
The scientific paradigm of human memory has fundamentally and irreversibly evolved from the passive playback of a static, video-like archive to the active, molecularly regulated reconstruction of a dynamic neural tapestry. At the cellular level, the double dissociation of initial consolidation and retrieval-induced reconsolidation proves that memories are not merely stored away; they are continuously maintained, destabilized, and rewritten through intricate protein synthesis, degradation pathways, and the activity of kinases like PKM$\zeta$ 1218. At the systems level, high-resolution fMRI reveals that cortical networks do not record continuous streams, but rather utilize event boundaries to capture episodic snapshots, which are subsequently integrated and physically pruned by microglial immune cells during the complex architecture of sleep 78.
Crucially, this intricate neurobiological scaffolding is ultimately shaped by the human narrative condition. The semantic structures we use to reconstruct our past are deeply influenced by cultural life scripts, proving that memory is as much a sociocultural phenomenon as a strictly biological one 3248. By fully embracing the reconstructive nature of memory, society can permanently abandon the myth of the video camera. This paradigm shift paves the way for more just legal systems that recognize the fragility and temporal limits of eyewitness testimony, and it heralds a new era of clinical therapies that leverage memory's inherent malleability to systematically heal the deep neurological scars of trauma.