Neurobiology of dreaming and the subconscious
Neural Architecture of Sleep States
The phenomenon of dreaming represents one of the most complex cognitive states generated by the human brain. Occurring largely independent of external sensory input, dreaming is a state of consciousness in which the brain constructs rich, multimodal narrative experiences 123. Historically relegated to the domains of mysticism or early psychoanalysis, the study of dreaming has matured into a rigorous discipline within cognitive neuroscience. To understand how dreams are generated, and what they reveal about subconscious processing, it is necessary to examine the brain's activity across the entire sleep-wake cycle, tracking the profound shifts in regional brain activation, oscillatory dynamics, and neurochemistry 245.
Wakefulness and Sleep Initiation
The regulation of sleep and wakefulness is governed by a delicate balance of ascending arousal systems and sleep-promoting nuclei. During wakefulness, a network of brainstem and hypothalamic nuclei - often referred to as the ascending reticular activating system (ARAS) - secretes high levels of monoamines. This includes norepinephrine from the locus coeruleus, serotonin from the dorsal raphe nucleus, and dopamine from the ventral tegmental area and substantia nigra 567. Additional wake-promoting neurochemicals include histamine from the tuberomammillary nucleus and orexin (hypocretin) from the lateral hypothalamus 568. Together, these neurotransmitters maintain cortical desynchronization, high-frequency brainwaves, and alertness 45. Concurrently, acetylcholine originating from the basal forebrain and brainstem is highly active, facilitating sensory processing, memory, and active cognition 58.
As the brain transitions into non-rapid eye movement (NREM) sleep, specifically the initial sleep-onset phase of Stage N1, the neurochemical environment shifts drastically. Sleep-promoting neurons in the ventrolateral preoptic nucleus (VLPO) and median preoptic nucleus (MNPO) release gamma-aminobutyric acid (GABA) and galanin. These inhibitory neurotransmitters actively suppress the ascending arousal systems 56. During Stage N1, which lasts approximately five to ten minutes, individuals lose awareness of their surroundings and experience a slowing of brain waves into the theta band (four to seven cycles per second) 810.
Progression into Stage N2 is marked by further disconnection from the external environment, characterized electrophysiologically by sleep spindles and K-complexes - waveforms believed to protect sleep continuity and facilitate memory consolidation 68. Finally, the brain enters Stage N3, also known as slow-wave sleep. This stage is defined by high-amplitude, low-frequency delta waves (0.5 to 4.0 Hz) generated by corticothalamocortical loops 4109. During deep NREM sleep, the levels of norepinephrine, serotonin, histamine, and orexin decline steadily 710. Acetylcholine levels also drop to their lowest point, allowing the cortex to engage in synchronized oscillatory rhythms that facilitate physical restoration and the replenishment of adenosine triphosphate (ATP) 789.
Rapid Eye Movement Sleep
The transition to rapid eye movement (REM) sleep introduces a paradoxical neurochemical and electrophysiological state. REM sleep is characterized by the near-complete cessation of serotonin, norepinephrine, and histamine release, as the neurons producing these chemicals are directly inhibited by GABAergic pathways 710. However, acetylcholine release surges to levels equal to or greater than those seen during active wakefulness 67. This intense cholinergic activity initiates ponto-geniculo-occipital (PGO) waves - electrical discharges originating in the pons and propagating to the lateral geniculate nucleus and occipital cortex 8911.
These PGO waves drive intense cortical activation, generating the vivid, hallucinatory visual imagery typical of REM dreams 89. The brain's metabolic rate during REM sleep can increase by up to 20% compared to deep NREM sleep, rivaling the energy consumption of a waking brain 6. Furthermore, dopamine neurons in the ventral tegmental area switch to a burst-firing mode during REM sleep, a mechanism thought to contribute to the emotional intensity, hyper-associativity, and reward-processing aspects of dream content 12. During this heightened cerebral activity, descending inhibitory signals from the subcoeruleus nucleus actively paralyze the skeletal muscles, preventing the dreamer from physically acting out their oneiric narratives 612.
Non-Rapid Eye Movement Dreaming
Early sleep research operated on the assumption that dreaming was an exclusive property of REM sleep. However, subsequent awakening studies demonstrated substantial recall of mental experiences from NREM sleep, with some estimates placing NREM recall rates at approximately 42.5%, compared to 81.8% for REM sleep 9. While NREM dreams are generally reported as shorter, more thought-like, less bizarre, and less emotional than REM dreams, their existence indicates that dream generation is not strictly confined to a single physiological state 913.
Researchers suggest that "covert REM" processes - localized activations of the cortex that occur outside of polysomnographically scored REM sleep - may account for NREM dreaming 9. Event-related functional magnetic resonance imaging (fMRI) studies have demonstrated that NREM sleep is not a state of uniform neural quiescence. Instead, it involves phasic increases in brain activity synchronized to specific NREM oscillations, such as sleep spindles, which may support the generation of less vivid, episodic dream fragments 14.
| Neurotransmitter / Neuromodulator | Role in Wakefulness | Levels in NREM Sleep | Levels in REM Sleep | Primary Anatomical Source |
|---|---|---|---|---|
| Acetylcholine (ACh) | High (promotes alertness and cognition) | Low | High (drives cortical activation and PGO waves) | Basal forebrain, pontine tegmentum |
| Norepinephrine (NE) | High (attention, stress response) | Decreased | Minimal/Absent | Locus coeruleus |
| Serotonin (5-HT) | High (mood, arousal) | Decreased | Minimal/Absent | Dorsal raphe nucleus |
| Histamine | High (wakefulness promotion) | Decreased | Minimal/Absent | Tuberomammillary nucleus |
| Dopamine (DA) | High (motivation, motor control) | Decreased | Burst Firing | Ventral tegmental area |
| Orexin (Hypocretin) | High (sustains wakefulness) | Decreased | Minimal/Absent | Lateral hypothalamus |
| GABA | Low | High (inhibits arousal systems) | High (maintains sleep state) | VLPO, anterior hypothalamus |
Topographical Brain Activity During Dreaming
Advanced neuroimaging techniques, including fMRI and positron emission tomography (PET), reveal that the dreaming brain is characterized by highly specific regional activations and deactivations. The phenomenological experience of dreaming is directly correlated with this unique topographical profile.
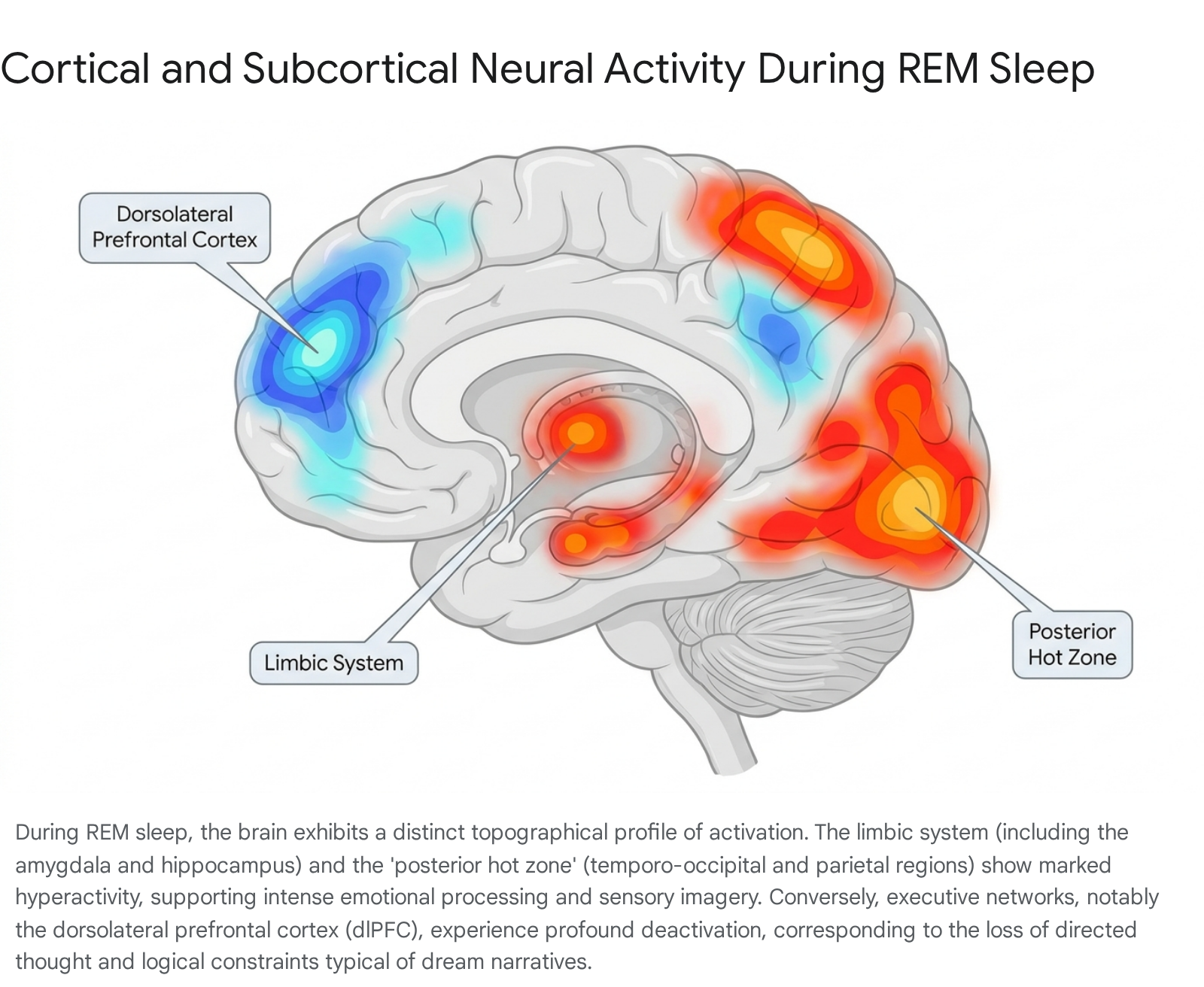
The Posterior Hot Zone
Recent high-density EEG and fMRI studies have identified a "posterior hot zone" critical for the generation of conscious experiences during sleep. This network encompasses the temporo-occipital and posterior parietal cortices 2913. When individuals report dreaming upon awakening from either NREM or REM sleep, neuroimaging consistently shows a localized decrease in low-frequency EEG activity and a simultaneous increase in high-frequency gamma (30-80 Hz) and fast beta band oscillatory activity within this hot zone 913.
This high-frequency activity reflects increased neuronal firing and is hypothesized to mediate the perceptual and spatial features of dream imagery 913. Specifically, activation in the visual, auditory, and somatosensory cortices within this posterior network correlates with the vividness of dreamt perceptions, such as faces, locations, and acoustic phenomena 215. The temporo-parietal junction (TPJ), a region responsible for multisensory integration and visual imagery during wakefulness, is also deeply involved in synthesizing these internally generated stimuli into a cohesive experiential environment 11.
Limbic System Hyperactivity
Concomitant with the activation of the posterior hot zone, REM sleep dreaming involves intense activation of limbic and paralimbic structures. Metabolic rates and blood oxygen level-dependent (BOLD) signals are significantly elevated in the amygdala, hippocampus, parahippocampal gyrus, and anterior cingulate cortex 61415. The amygdala, a core center for emotional processing and fear conditioning, shows metabolic activity during REM sleep that often exceeds waking baseline levels 1216.
This limbic hyperactivity correlates directly with the heightened emotionality, anxiety, and elation frequently reported in dream narratives. The concurrent activation of the hippocampus ensures that recent episodic memories and emotional experiences are accessed and integrated into the unfolding dream, albeit often in fragmented or hyper-associative ways 2615.
Prefrontal Deactivation and the Default Mode Network
While sensory and emotional cortices operate at high capacity, executive control networks experience profound suppression. Meta-analyses of functional neuroimaging data demonstrate consistent, significant deactivations in the dorsolateral prefrontal cortex (dlPFC) and the inferior, superior, and middle frontal gyri during both REM and deep NREM sleep 171819. The dlPFC is fundamentally responsible for working memory, executive function, reality testing, volitional control, and logical reasoning 1720. The uncoupling of the hyperactive limbic system from the regulatory oversight of the deactivated prefrontal cortex explains the hallmark phenomenology of standard dreams: an acceptance of bizarre or impossible scenarios without critical reflection, a lack of voluntary control over the narrative, and an inability to maintain stable working memory across scene transitions 1321.
Furthermore, research indicates complex alterations in the Default Mode Network (DMN) - a highly interconnected set of brain regions, including the medial prefrontal cortex, posterior cingulate cortex (PCC), and precuneus, associated with self-referential thought and mind-wandering 1722. While the DMN is active during rest, event-related fMRI studies show that specific DMN nodes exhibit modest, phase-locked deactivations that coincide precisely with the rapid eye movements of REM sleep 222324. This phasic deactivation is theorized to facilitate hierarchical predictive coding, allowing the primary visual cortex to scan and interact with the internally generated virtual environment of the dream without interference from higher-order self-monitoring systems 24.
Theoretical Frameworks of Dream Generation
The scientific consensus has definitively shifted away from the early twentieth-century view that dreams are purely chaotic artifacts of sleep. Instead, contemporary theoretical frameworks attempt to map the neurophysiological phenomena of dreaming to specific evolutionary, cognitive, and affective functions.
The Activation-Synthesis and AIM Models
Proposed in 1977 by neuroscientists John Allan Hobson and Robert W. McCarley, the Activation-Synthesis hypothesis offers a strictly neurobiological explanation for dream generation. The theory argues that dreams originate from spontaneous, semi-random neural discharges - specifically the cholinergic PGO waves - originating in the brainstem during REM sleep 1516. According to this model, these ascending signals indiscriminately stimulate the forebrain. The cortex, striving to impose order on this chaotic sensory and motor information, attempts to synthesize it into a coherent narrative, utilizing fragments of memory and current emotional states 16.
Hobson later refined this into the Activation-Input-Modulation (AIM) model, which maps states of consciousness across three dimensions: overall brain activation, the source of input (external sensory vs. internal generated), and neuromodulatory balance (aminergic vs. cholinergic) 916. While Activation-Synthesis accurately maps the bottom-up physiological initiation of REM sleep, it is widely criticized by cognitive psychologists. Critics argue that if dreams were merely the byproduct of random neural noise, dream reports would be highly disjointed and nonsensical. Instead, dreams frequently feature robust, continuous narratives, clear thematic structures, and highly specific incorporations of waking concerns, suggesting a top-down organizational principle that the theory fails to address 16.
Threat Simulation Theory
Approaching dreaming from an evolutionary psychology perspective, Antti Revonsuo introduced the Threat Simulation Theory (TST). This theory posits that dreaming is an ancient, highly organized biological defense mechanism selected by evolution to repeatedly simulate threatening events in a safe, offline environment 252627. By placing the dream-ego in virtual peril, the brain rehearses threat-perception and evasive motor patterns, thereby enhancing waking survival skills 2628.
Empirical support for TST is robust within specific paradigms. Content analyses of normal dreams consistently reveal an over-representation of negative emotions, physical dangers, pursuits, and aggressive encounters compared to waking life 2528. Furthermore, TST predicts that waking exposure to severe, real-world threats will trigger a hyper-activation of the simulation system. A landmark 2005 study comparing severely traumatized Kurdish children living in conflict zones to non-traumatized Finnish children confirmed this prediction. The traumatized cohort reported a significantly higher volume of dreams, and these dreams contained threats that were far more frequent, realistic, and severe 26272930.
However, TST is not universally accepted. Detractors point to a 2008 study comparing populations in high-crime areas of South Africa with those in low-crime areas of Wales. Despite higher exposure to real-world threats, the South African participants reported fewer threat-based dreams 26. Furthermore, comprehensive analyses of dream databases reveal that less than 15% to 20% of dreams depict realistic survival situations, and successful evasive actions are exceedingly rare, raising questions about the efficiency of dreams as an evolutionary training mechanism 26.
Emotion Regulation and Memory Consolidation
Currently, one of the most empirically supported models posits that dreaming plays an active, indispensable role in sleep-dependent memory consolidation and emotion regulation. According to this framework, dreams facilitate the integration of novel, emotionally charged waking experiences into long-term cortical networks while simultaneously dampening the acute affective distress originally associated with those memories 313233. This process is hypothesized to protect the individual from chronic waking anxiety, serving as a form of nocturnal exposure therapy 3435.
A seminal 2024 study conducted by Jing Zhang and Sara Mednick at the UC Irvine Sleep and Cognition Lab provided direct empirical evidence for this regulatory function. Participants were exposed to highly negative emotional images before sleep and tested the following morning. The researchers documented a specific "trade-off" effect: emotionally charged negative memories were prioritized for consolidation (retained at the expense of neutral memories), yet the participants' subjective emotional reactivity to those same images was significantly reduced upon waking 31.
Crucially, this adaptive pattern - enhanced memory retention coupled with decreased emotional reactivity - was entirely absent in subjects who did not recall dreaming 31. The data also revealed that the emotional valence of the dream directly influenced next-day mood; participants who reported more positive dream affect rated negative images much less severely the following morning 3132.
This psychological mechanism aligns perfectly with the neurobiology of REM sleep. The hyper-activation of the amygdala and hippocampus allows traumatic or emotional memories to be accessed and replayed. However, because this neural replay occurs in a neurochemical environment completely devoid of the stress-mediating neurotransmitters norepinephrine and serotonin, the brain is able to process and integrate the memory without triggering the peripheral physiological stress response 6710. When this regulatory system fails - often due to an overwhelming accumulation of waking distress (affect load) combined with a fragmented sleep architecture - the emotion regulation process breaks down, resulting in the distressing, unmitigated threat simulations recognized as clinical nightmares 323335.
| Theoretical Model | Core Proposed Mechanism | Hypothesized Biological/Psychological Function | Key Empirical Evidence and Observations |
|---|---|---|---|
| Activation-Synthesis / AIM | Brainstem PGO waves activate forebrain; cortex synthesizes narratives from random neural noise. | Epiphenomenal; the brain's attempt to impose meaning on physiological maintenance processes. | High brainstem and cortical activity during REM; occurrence of highly bizarre, discontinuous dream elements 16. |
| Threat Simulation Theory | Offline generation of highly organized, virtual threat scenarios involving the dream-ego. | Evolutionary rehearsal of threat-perception and physical avoidance behaviors. | Over-representation of threats in normative dreams; highly activated simulation systems in traumatized populations 252728. |
| Emotion Regulation | Reactivation of affective memories within an aminergic-depleted neurochemical environment. | Downregulation of emotional arousal; integration of traumatic memories without stress response. | Waking stressors reduce next-day emotional reactivity only in dream recallers; memory trade-off effects 3134. |
| Continual-Activation | Continuous data transfer from short-term to long-term memory structures during sleep. | Maintenance and organization of procedural and declarative memory networks. | Consistent incorporation of recent waking life events into dream content (Continuity Hypothesis) 16. |
Cognitive Paradigms of the Subconscious
While modern neuroscience relies on objective, quantifiable metrics, recent empirical findings have increasingly validated modernized interpretations of early psychoanalytic theories regarding the subconscious mind. Historically, psychoanalysis viewed dreams as obfuscated expressions of the unconscious, heavily utilizing mechanisms like condensation and displacement to disguise repressed desires 3836. Modern neuropsychoanalysis suggests that while the rigid Freudian concept of active "manifest disguise" is largely unsupported by cognitive science 37, the foundational premise - that dreams are highly structured, meaning-laden self-representations of the psyche - remains scientifically valid 383940.
Structural Dream Analysis and Psychotherapy
The application of Structural Dream Analysis (SDA) in clinical psychotherapy provides a robust, quantitative framework for tracking changes in subconscious processing. SDA moves beyond interpreting specific symbols, focusing instead on the underlying architecture of the dream and the behavioral patterns of the "dream-ego" - the representation of the self within the oneiric environment 383941.
Research indicates that the dream-ego's capacity for agency, emotional expression, and social interaction closely mirrors the individual's waking psychological state. In a longitudinal study of patients undergoing psychoanalytic psychotherapy, researchers documented a statistically significant upward movement in the hierarchical typology of dream patterns as clinical improvement occurred 3839. As patients resolved waking trauma, their dream-egos exhibited increased initiative, transitioning from passive victimization to active problem-solving within the dream narrative 3842. Furthermore, the appearance of specific archetypal patterns, such as Jung's "child motif," consistently correlated with transformative turning points in the psychotherapeutic process, signaling a structural alteration in the patient's subconscious conflict resolution 42. This body of evidence strongly supports the "continuity hypothesis," confirming that subconscious processing in dreams is not severed from waking reality but is deeply intertwined with ongoing emotional development 4243.
Cultural Encoding of Dream Narratives
If dreams reflect the foundational structures of the subconscious, recent cross-cultural structural analyses demonstrate that the subconscious mind is fundamentally shaped by cultural self-construal. A 2025 study utilizing SDA compared dream reports from American, Japanese, and Chinese populations to determine how cultural frameworks influence the behavior of the dream-ego 41.
The analysis revealed stark divergences. American dreams, reflecting Western independent self-construal, are characterized by high dream-ego agency, proactive behavior, and a strong emphasis on individual achievement. The American dream-ego frequently confronts threats directly and actively manipulates its environment to achieve social status or success 41.
Conversely, Japanese dreams reflect an interdependent but culturally self-effacing orientation. The Japanese dream-ego exhibits a marked "fragility of agency," tending to be passive, vague, and prone to fading or dissolving into crowds. Structurally, Japanese dreamers are statistically more likely to report the dream-ego suffering diffuse harm or wounding without mounting active resistance 41.
Chinese dreams present a distinct third pattern. While also reflecting East Asian interdependent self-construal through a generally passive stance toward spontaneously arising challenges (e.g., being forced to take an examination), the Chinese dream-ego is deeply sustained by relational support. The foundation of selfhood in Chinese dreams is maintained by moving toward affirming social figures, such as family members, cultivating a shared sense of humanity rather than the ego dissolution seen in the Japanese sample 41. These findings indicate that the subconscious mind does not operate on purely universal, biological algorithms; rather, the foundational concepts of agency, selfhood, and social relation are culturally encoded and manifest vividly during sleep 4144.
Metacognition and Lucid Dreaming
Lucid dreaming - a state in which an individual becomes consciously aware that they are dreaming while remaining asleep - provides a unique window into the intersection of unconscious generation and conscious executive control 20.
Neurobiologically, lucid dreaming represents a hybrid state of consciousness. It is characterized by the localized reactivation of the dorsolateral prefrontal cortex (dlPFC) and the anterior prefrontal cortex, areas that are normally profoundly suppressed during REM sleep 2021. This prefrontal reactivation is accompanied by a marked increase in high-frequency gamma band oscillations (30 - 36 Hz) and alpha activity (8 - 12 Hz) across fronto-parietal networks 20.
The return of prefrontal activity reintroduces metacognitive awareness and higher-order cognitive control to the sleeping brain. The dreamer regains the capacity to evaluate incongruous information, allowing them to recognize the bizarre, hallucinatory nature of the dream environment 21. Because this state permits volitional control over the unfolding narrative, researchers and clinicians are actively exploring lucid dream therapy (LDT). By training patients with Post-Traumatic Stress Disorder (PTSD) to achieve lucidity, patients can actively confront and rewrite the narratives of recurrent nightmares, thereby regulating their subconscious fear responses and demonstrating significant long-term reductions in waking anxiety 2045.
Environmental and External Modulators
The subconscious mind does not process information in an environmental vacuum. External variables significantly influence sleep architecture, which in turn alters the generation, thematic content, and regulatory efficacy of dreams. Recent epidemiological and environmental research highlights how urbanization, artificial illumination, and climate-driven temperature changes impact the sleeping brain 46474849.
Artificial Light at Night and Noise
Artificial light at night (ALAN) is a pervasive environmental disruptor, particularly in rapidly urbanizing environments. Exposure to ALAN disrupts the natural circadian rhythm by suppressing the synthesis of melatonin, the hormone responsible for timing sleep onset 50. This suppression fragments overall sleep architecture, often reducing the total time spent in restorative NREM deep sleep and altering REM latency 54.
Similarly, urban transportation noise - particularly road and rail noise exceeding 65 decibels - causes micro-arousals throughout the night. These arousals prevent the brain from sustaining the prolonged, uninterrupted REM cycles necessary for effective emotional memory consolidation 47. When the physical environment disrupts sleep continuity, the psychological function of dreaming is compromised. Nightmares and distressing dreams are frequently the result of an elevated "affect load" combined with these environmental interruptions, leading to increased daytime irritability and cognitive impairment 3351.
Ambient Temperature and Climate Disruptions
Ambient environmental temperature plays a critical, yet historically under-examined, role in sleep architecture. The human body requires a slight drop in core temperature to initiate and maintain deep sleep 10. Increased ambient nighttime temperatures - exacerbated by global climate change and urban heat islands - severely impede this thermoregulatory process.
A massive 2026 epidemiological study utilizing sensor data from 219,808 global consumers tracked sleep patterns through the record-breaking heatwaves of 2023. The data revealed that extreme heat events correlate with a 30% to 300% increase in the prevalence of poor sleep (defined as less than 6 hours per night), accounting for an estimated loss of 2.2 million disability-adjusted life years globally 4852. High temperatures disproportionately suppress REM sleep, the stage most heavily associated with complex dreaming and emotional depotentiation 5253. Consequently, climate-induced sleep fragmentation not only blunts the brain's ability to engage in subconscious affect regulation but also contributes to rising rates of waking anxiety, depression, and eco-anxiety 51535455.
Methodological Advances in Dream Decoding
Historically, the primary limitation of dream research was its absolute reliance on subjective, retrospective reporting upon waking. This reliance made the science vulnerable to waking amnesia, narrative confabulation, and the inherent difficulty of translating highly visual and emotional experiences into waking language 23. However, recent breakthroughs in functional neuroimaging, machine learning, and computational linguistics are enabling researchers to map neural firing patterns directly to oneiric narratives, bypassing the limitations of subjective recall.
Functional Neuroimaging and Visual Reconstruction
Advanced fMRI decoding techniques leverage the principle of representational similarity to reconstruct dream imagery. Researchers have established that the neural activity patterns observed when a person perceives a specific visual stimulus while awake closely match the neural patterns generated when they dream of that same stimulus 111556.
In a pioneering methodology, researchers map the fMRI time series of participants onto a common surface map (fsLR32K space), isolating early and higher visual cortical regions (such as V1, V2, V4, and the fusiform face area) 56. By training machine learning algorithms on a subject's fMRI data during waking visual perception, scientists can decode the semantic content of the visual imagery occurring in the posterior hot zone during REM sleep 5657.
In a recent 2025 study, participants were exposed to different audiobooks before sleep to prime memory integration. Using multivariate pattern analyses, researchers demonstrated that the specific content of the audiobooks was actively reprocessed in the sleeping brain, identifiable via distinct beta band power patterns during REM sleep. Blind raters analyzing the algorithmic output were able to successfully determine which narrative the subjects had been exposed to based purely on their neural reinstatement during dreaming 5758.
Natural Language Processing of Dream Reports
Simultaneously, Natural Language Processing (NLP) and large language models are being deployed to analyze vast databases of transcribed dream reports. These algorithms evaluate the semantic density, emotional valence, and narrative structure of dreams far faster and more consistently than human coders utilizing traditional scales like the Hall and Van de Castle system 424459.
A recent study evaluating text "perplexity" - a metric indicating how surprising or chaotic a sequence of words is to an AI model - yielded highly counterintuitive results. When analyzed by language models, dream reports exhibited significantly lower perplexity scores than standardized waking texts, such as Wikipedia articles 59. This indicates that despite the subjective bizarreness of dreams, the subconscious mind generates narratives that follow highly predictable, mathematically structured semantic and syntactic patterns 4459. These methodological advances are definitively shifting dream research from qualitative psychoanalysis to quantifiable cognitive neuroscience, proving that the subconscious operates as a highly organized, rule-bound system of information processing.