Functions of dreaming during REM sleep
Historical Context and Evolution of Dream Research
For centuries, the phenomenon of dreaming was relegated to the domains of mysticism, philosophy, and psychoanalysis. Early twentieth-century frameworks, most notably those of Sigmund Freud and Carl Jung, posited that dreams were heavily symbolic expressions of suppressed wishes or compensatory psychological manifestations 122. Freud's seminal 1900 publication, The Interpretation of Dreams, popularized the notion that the bizarre content of nocturnal experiences originated in complex psychological mechanisms fundamentally dissimilar from those governing normal waking cognition 12. Conversely, Jung proposed a compensatory theory of dreaming, suggesting that the "self-regulation of the psychic organism" drove dream narratives, a concept that conceptually aligns more closely with modern physiological views of emotional homeostasis 2.
Despite their historical significance, psychoanalytic approaches lacked empirical neurobiological grounding. The discovery of rapid eye movement (REM) sleep by Eugene Aserinsky and Nathaniel Kleitman in 1953 catalyzed a paradigm shift, transitioning dream research into the realm of quantitative neuroscience 36. Initially, the prevailing neurobiological view of the late twentieth century was the "activation-synthesis hypothesis," proposed by J. Allan Hobson and Robert McCarley in 1977 45. This model suggested that dreaming was merely an epiphenomenon - the forebrain's attempt to impose a semblance of narrative structure upon random, chaotic electrical signals (pontogeniculooccipital waves) originating in the brainstem during sleep 56. Under this framework, the content of dreams was considered biologically meaningless "noise" 17.
Modern neuroimaging, high-density electroencephalography (EEG), and computational cognitive psychology have since dismantled the assertion that dreams are random epiphenomena. Current empirical evidence indicates that dreaming is a highly structured, state-dependent cognitive process 18. The scientific consensus has coalesced around several functional theories: memory consolidation (the reorganization and integration of newly acquired episodic and semantic information), emotional regulation (the recalibration of affective autonomic responses), and evolutionary simulation (the nocturnal rehearsal of threat avoidance and social interactions). Rather than a passive byproduct of sleep, dreaming is now understood as an active neurophysiological state essential for psychological homeostasis, cognitive plasticity, and adaptive waking behavior 14910.
Neuroanatomical and Neurochemical Architecture of Sleep
To evaluate the functional hypotheses of dreaming, it is necessary to examine the physical state of the sleeping brain. The subjective phenomenology of a dream - characterized by vivid visual imagery, intense emotionality, disrupted temporal awareness, and profound logical incongruities - corresponds directly to highly specific patterns of regional brain activation and neurochemical modulation during REM sleep.
Regional Brain Metabolism in REM Sleep
Functional magnetic resonance imaging (fMRI) and positron emission tomography (PET) reveal that REM sleep is not a state of global neural quiescence; rather, its overall glucose metabolism and oxygen consumption rival, and in specific regions exceed, those of the waking brain 311. However, this activation is anatomically selective.
The functional neuroanatomy of REM sleep features a distinct dichotomy between hyperactive limbic and visual networks and hypoactive executive networks.
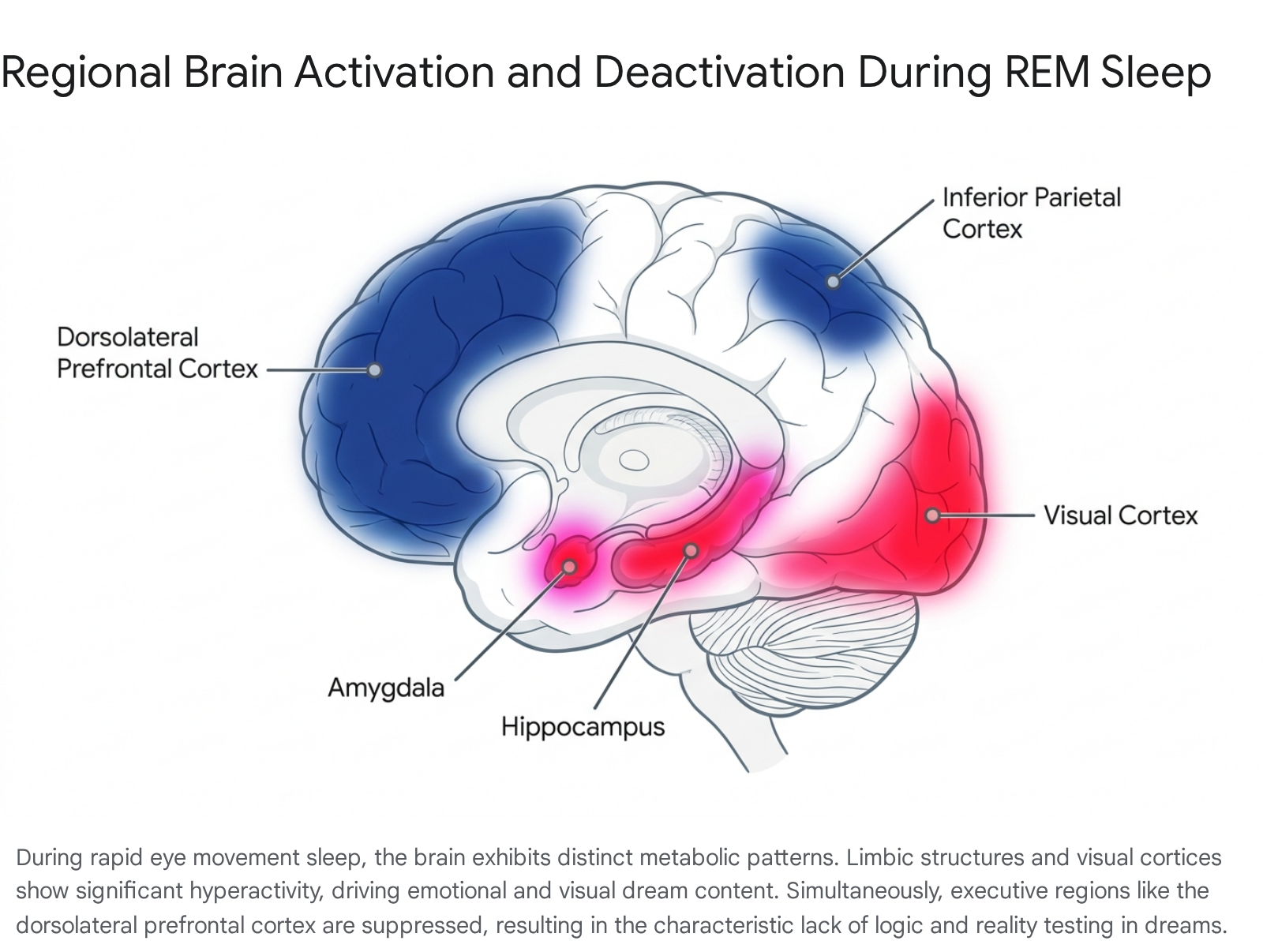
Specifically, the amygdala, hippocampus, anterior cingulate cortex (ACC), and pontine tegmentum demonstrate marked increases in activation compared to waking states 111213. The amygdala and ACC are central to emotional processing, threat detection, and affective regulation, providing a direct physiological explanation for the high emotional intensity and anxiety frequently reported in REM dreams 121415. Visual association areas, such as the temporo-occipital cortices and the parahippocampal gyrus, also surge with activity, generating the immersive, hallucinatory perceptual qualities of the dream state 111216.
Conversely, REM sleep is characterized by significant metabolic deactivation in the dorsolateral prefrontal cortex (dlPFC), orbitofrontal cortex, precuneus, and posterior cingulate gyrus 311131517. The prefrontal cortex serves as the brain's executive control center during wakefulness, mediating logic, working memory, impulse control, volition, and reality testing 1216. The suppression of these frontal regions accounts for the characteristic cognitive deficiencies of the dream state: disorganized temporal sequencing, amnesia upon waking, delusional acceptance of bizarre narratives, and a profound lack of insight 14151617.
The following table summarizes the metabolic activation states of key neural structures during REM sleep and their corresponding cognitive manifestations within the dream environment:
| Neural Structure | Activation State in REM | Primary Waking Function | Dream State Manifestation |
|---|---|---|---|
| Amygdala | Hyperactive | Threat detection, emotional arousal | Intense affective themes, fear, anxiety 1214. |
| Hippocampus | Hyperactive | Episodic memory retrieval | Incorporation of waking memory fragments 1214. |
| Visual Association Cortex | Hyperactive | Complex visual processing | Immersive, vivid hallucinatory imagery 1216. |
| Dorsolateral Prefrontal Cortex | Hypoactive | Executive function, logic, volition | Delusional acceptance of bizarre narratives, lack of insight 1516. |
| Primary Motor Cortex | Hyperactive | Motor command generation | Perceived physical movement (though muscle atonia prevents actual execution) 1216. |
The Neuromodulatory Environment
The distinct cognitive state of dreaming is further governed by a radical shift in the brain's neurochemical environment. Waking consciousness relies on an orchestrated balance of neurotransmitters, including acetylcholine, noradrenaline (norepinephrine), and serotonin 18. During transitions into non-rapid eye movement (NREM) and subsequently REM sleep, this aminergic-cholinergic balance alters drastically.
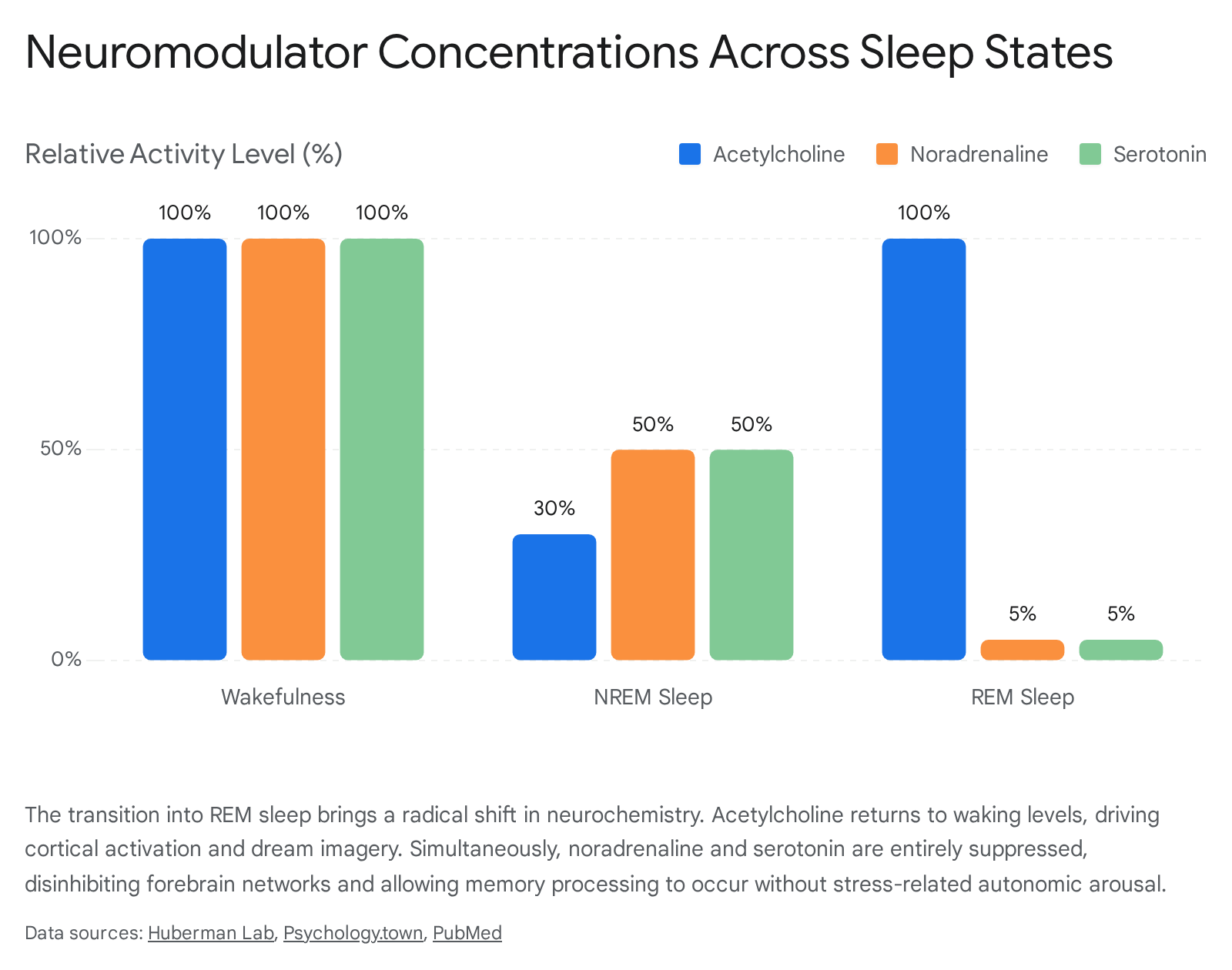
Acetylcholine, released by cholinergic neurons in the laterodorsal and pedunculopontine tegmental nuclei, rises to waking or super-waking levels during REM sleep 1819. This cholinergic dominance is responsible for generating the fast, desynchronized cortical EEG waves (such as theta and gamma rhythms) that make REM sleep resemble wakefulness neurologically. This activity promotes internal sensory processing and memory consolidation independent of external environmental input 51820.
Conversely, the release of aminergic neurotransmitters - specifically noradrenaline from the locus coeruleus and serotonin from the dorsal and median raphe nuclei - ceases almost entirely during REM sleep 91921. Noradrenaline is the brain's primary stress, alertness, and "fight-or-flight" chemical, while serotonin acts to modulate mood, reality monitoring, and waking behavioral inhibition 18.
This state of "aminergic demodulation" combined with high cholinergic activity creates a microenvironment where forebrain networks are fundamentally disinhibited 21. Theoretical models of neural dynamics suggest that this specific neurochemical profile shifts network outputs toward mathematical "errors," which are subjectively experienced as the bizarre incongruities characteristic of dream plots 21. Sudden bursts of pontogeniculooccipital (PGO) waves introduce abrupt cholinergic signals to the visual pathways of the forebrain, causing unpredictable bifurcations in neural activity that correlate with sudden scene shifts and discontinuities in cinematic dreams 521.
Memory Consolidation and the Neural Replay Phenomenon
One of the most robust empirical findings in cognitive neuroscience is that sleep facilitates memory consolidation - the process by which fragile, newly encoded memory traces are stabilized, reorganized, and integrated into long-term cortical storage 102223.
Evidence for Memory Replay
Animal models have repeatedly demonstrated that learning-related neural activity is re-expressed during post-training sleep. Specifically, hippocampal place cells, which fire in precise sequences as a rodent navigates a spatial environment while awake, spontaneously reactivate in the identical temporal sequence during subsequent sleep 222425. This phenomenon, known as "neural replay" or memory reactivation, occurs via high-frequency electrical bursts called sharp-wave ripples 222526.
If neural replay biologically consolidates memories, it is a logical hypothesis that the psychological phenomenon of dreaming represents the subjective, conscious experience of this cellular replay. Numerous behavioral studies in humans support this connection. Researchers have found that learning a novel, engaging task prior to sleep influences subsequent dream content 12728. In a landmark study utilizing a virtual 3D maze navigation task, participants who napped and subsequently reported dreams incorporating maze-related imagery demonstrated a tenfold greater improvement in post-sleep task performance compared to those who napped but did not dream of the maze 72829. Notably, the participants who incorporated the maze into their dreams performed more poorly during the initial pre-sleep baseline testing; the profound overnight improvement brought them up to par with the non-incorporating group, indicating that the sleeping brain may actively prioritize the consolidation of tasks that were found to be exceptionally challenging during wakefulness 2829.
Discrepancies Between Replay and Conscious Dreaming
Despite the correlation between task-related dreaming and performance enhancement, neuroscientists caution against equating cellular "replay" with the narrative "replay" of dreams. Evidence suggests that episodic memories are rarely, if ever, replayed in dreams exactly as they occurred in waking life 1822. In comprehensive analyses matching hundreds of dream reports to waking diaries, over 50% of dreams contained at least one feature related to a recent waking experience, but fewer than 2% of dreams replicated a specific waking episode with exact situational context, characters, and actions 122.
Furthermore, the mechanisms and timescales of these phenomena are vastly different. Neural replay occurs in the range of hundreds of milliseconds, frequently on a faster timescale than the original experience, and is predominantly observed during early-night NREM sleep 2225. Conversely, complex, immersive, cinematic dreaming unfolds over minutes and occurs most vividly during late-night REM sleep 625. Studies also show that replay systematically deviates from direct recapitulation; replay can activate never-experienced novel trajectories, unroll in reverse order after a reward, and over-represent salient events rather than strictly echoing the temporal structure of the waking episode 30. This suggests that the brain engages in active reconstruction and abstraction rather than passive recording playback 3031.
Meta-Analyses of Sleep Stages and Task Performance
Recent meta-analytic data further complicates the relationship between specific sleep stages, dreaming, and memory. A 2023 meta-analysis by Hudachek and Wamsley aggregated 45 effects across 16 qualifying studies to determine the strength of the association between learning-related dreams and post-sleep memory improvement 242732. Integrating across all effects, the authors reported a strong, statistically significant positive association (Standardized Mean Difference = 0.51) between task-related dreaming and memory performance 2427.
However, a critical nuance emerged when the researchers analyzed studies utilizing polysomnography to isolate sleep stages. The relationship between task-related dreaming and memory enhancement was statistically significant for dreams collected from NREM sleep (across 10 studies), but not for dreams collected from REM sleep (across 12 studies) 2427. This divergence suggests that explicit memory reactivation tied to immediate performance gains may be heavily reliant on NREM processes, whereas REM dreaming serves a different cognitive function related to broader associative integration rather than strict declarative or procedural consolidation 2732.
The NEXTUP Model of Network Exploration
To account for the bizarre, fragmented nature of REM dream content and the lack of straightforward memory replay, sleep researchers Robert Stickgold and Antonio Zadra proposed the NEXTUP model (Network Exploration to Understand Possibilities) 333435. NEXTUP posits that dreaming is a unique form of sleep-dependent memory processing that does not seek to replay memories, but rather to extract overarching rules and discover novel knowledge from existing information 63335.
Theoretical Mechanisms of Weak Associations
Under the NEXTUP framework, the sleeping brain actively searches for and strengthens "weak associations" between recent episodic memories and vast networks of remote semantic memories 333536. Because the prefrontal cortex is suppressed and the focusing chemical noradrenaline is absent, the brain is liberated to drift into loose, associative networks without the strict constraints of waking logic 37.
During REM sleep, the brain stitches these disparate memory fragments together into a simulated narrative 8. By evaluating the emotional resonance of these novel associations as the dream unfolds, the brain determines whether the newly formed connections contain useful predictive information for future survival or problem-solving 6. Therefore, the bizarreness of dreams is not an error resulting from random neural noise, but a deliberate feature of an algorithm searching for abstract connections and divergent problem-solving pathways 3337. The theory also suggests that the conscious recall of dreams is not necessary for them to fulfill their biological function; the cognitive work of network exploration is completed during the sleep cycle regardless of morning amnesia 37.
Empirical Tests and Limitations
While NEXTUP offers a compelling explanatory framework for dream bizarreness, establishing direct causality remains scientifically challenging. A 2024 study published in the context of a Festschrift for Dr. Stickgold attempted to test the NEXTUP hypothesis by artificially introducing new information about a participant's "favorite celebrity" (a well-established semantic network) to see if it would preferentially incorporate into dreams across three consecutive nights 363839. While the specific celebrity manipulation failed to statistically alter dream content, the resulting dataset provided rich descriptive evidence of how the brain interweaves recent waking fragments with deeply remote memories to construct imaginative scenarios, supporting the broader concept of associative network activation 3639.
Critiques of the NEXTUP model point out that while discovering weak associations may be beneficial, the theory must more clearly articulate the specific, periodic survival benefit that this process conferred during evolutionary history to become a target of natural selection . Furthermore, NEXTUP struggles to explain certain pathologies, such as "epic dreaming" - a condition where individuals experience relentless, exhausting, highly narrative dreams that are seemingly devoid of emotional content, for which psychological and pharmacological treatments have proven largely ineffective 33.
Emotional Regulation and the Sleep to Forget, Sleep to Remember Hypothesis
If the primary function of memory consolidation in NREM sleep is the stabilization of facts, procedural skills, and spatial navigation, researchers posit that the primary function of REM sleep is the processing of the emotional tone attached to those events. This mechanism is most prominently articulated in the "Sleep to Forget, Sleep to Remember" (SFSR) hypothesis developed by Matthew Walker and colleagues 2640.
Decoupling Affect from Memory
According to the SFSR model, REM sleep provides a unique "overnight therapy" by divorcing the visceral emotional charge of an experience from the informational core of the memory 91940. When an individual experiences a traumatic or highly stressful event during the day, the memory is encoded alongside a massive surge of autonomic arousal, driven by sympathetic nervous system activation, noradrenaline, and amygdala hyper-responsivity 941.
During subsequent REM sleep, these emotional memories are reactivated. However, because REM sleep is the only period in the 24-hour circadian cycle during which the brain is completely devoid of noradrenaline, the memory is replayed in a neurochemically calm, stress-free environment 791219. The active amygdala and hippocampus process the informational memory trace, while the absence of noradrenaline systematically depotentiates the autonomic distress previously associated with the event 4041.
Experimental evidence heavily supports this mechanism. In fMRI studies comparing subjects who slept between exposure to emotionally disturbing images versus those who remained awake, the sleep group demonstrated significantly reduced subjective emotional distress and a palliative decrease in amygdala reactivity upon re-exposure 7940. Furthermore, there was a reengagement of the rational prefrontal cortex in the sleep group, providing a top-down dampening brake on emotional reactivity 79. EEG recordings confirmed that the degree of overnight emotional depotentiation was directly correlated with a reduction in specific high-frequency gamma brain waves during REM sleep, which serve as a proxy for adrenergic stress chemistry 7917.
Recent research reinforces this framework. A 2024 study conducted at the UC Irvine Sleep and Cognition Lab examined 125 women completing emotional picture tasks. The findings demonstrated that participants who reported dreaming had better recall of negative images but were significantly less emotionally reactive to them the following day, providing empirical support for dreaming's active involvement in transforming emotional responses 42.
Pathologies of the Emotional Processing System
The critical nature of REM sleep in emotional regulation is starkly evident when the process fails. Insomnia is highly correlated with emotional dysregulation, daytime hyperarousal, and impaired threat extinction 4344. In chronic insomnia, fragmented REM sleep prevents the prolonged silencing of the locus coeruleus, meaning central noradrenaline levels do not drop sufficiently 44. Consequently, the brain attempts to process emotional memories while still bathed in stress chemistry, reinforcing rather than resolving the affective distress and creating a feedback loop of anxiety 44.
Similarly, Post-Traumatic Stress Disorder (PTSD) is characterized by repetitive, highly distressing nightmares. In healthy populations, the dreaming brain connects recent threats to remote, non-threatening memories to dilute the fear 3645. In PTSD, chronically elevated noradrenaline during sleep prevents this contextualization, causing the brain to endlessly replay the trauma in its original, terrifying form without ever achieving emotional depotentiation 72046. The administration of prazosin, an alpha-1 blocker that reduces central noradrenaline activity, has shown efficacy in reducing PTSD nightmares by artificially lowering the adrenergic tone during sleep, thus mimicking healthy REM conditions and allowing the SFSR mechanism to function 579.
Nuances in Generalization and Threat Extinction
While the SFSR theory is foundational, recent neurobiological research has introduced crucial nuances regarding how fear memories are altered. A 2021 fear-conditioning study utilizing sleep deprivation protocols and fMRI found that REM sleep had dual, opposing effects. While REM sleep impaired the precise recall of the original threat memory, it significantly improved the subject's ability to generalize the threat memory to novel, related situations that required discrimination between threat and safety signals 47. This suggests REM sleep calibrates the balance between specific memory retention and generalized adaptive survival skills 47.
Additionally, emerging literature utilizing high-density continuous recordings in animal models suggests that the "forgetting" aspect of sleep might actually be driven by the representational drift of memories during non-REM sleep, where newly recruited neurons update the memory to fit broader contexts, followed by REM sleep preserving and pruning the modified connections 48. Therefore, emotion regulation is likely a synergistic process relying on the cyclical interplay between NREM slow-wave sleep and REM theta rhythms, rather than the isolated function of REM alone 2649.
Evolutionary Perspectives and Simulation Theories
Parallel to memory and emotion theories are evolutionary frameworks that view dreaming not merely as a byproduct of maintenance processes, but as a specialized biological mechanism selected for its direct survival value. These theories, situated under the umbrella of "simulation theories," argue that dreaming creates an immersive virtual-reality environment to safely rehearse critical behaviors without physical risk 5051.
Threat Simulation Theory and its Critiques
Proposed by philosopher and cognitive scientist Antti Revonsuo in 2000, the Threat Simulation Theory (TST) posits that the dream generation system evolved to repeatedly simulate threatening events, effectively serving as a nocturnal "fire drill" 452. Ancestral human populations lived in environments fraught with severe physical, environmental, and predatory threats. According to TST, the brain utilizes the motor paralysis and sensory isolation of REM sleep to activate the amygdala and fear-conditioning networks, generating highly immersive threatening scenarios 45253.
By practicing threat perception and evasion in this virtual environment, early humans increased the probability of executing appropriate fight-or-flight responses in waking life, thereby conferring a direct reproductive advantage 454. Empirical support for TST rests on content analyses of dream reports across diverse populations, which consistently reveal a disproportionately high incidence of negative emotions - principally fear and anger - and aggressive encounters compared to waking life 45255.
However, TST faces substantial scholarly critique. Critics argue that TST focuses too heavily on the "chasing nightmare" as the paradigmatic dream, failing to account for the vast majority of dreams that are entirely mundane or completely lack threatening stimuli 54. Furthermore, empirical tests of TST's propositions have yielded contradictory results. A study tracking approach versus avoidant behavior in dreams compared participants from a high-crime environment (South Africa) to those from a low-crime environment (Wales) using the Most Recent Dream (MRD) method 53. The results indicated that dreamers exhibited significantly more approach behavior than avoidant behavior (χ2(1) = 43.89, p < 0.001), and that the prevalence of real-world environmental threat did not dictate the frequency of avoidant nightmares, directly contradicting TST's core prediction 53.
Social Simulation Theory and the Sociality Bias
In response to the limitations of TST, Revonsuo and colleagues subsequently developed the Social Simulation Theory (SST). SST proposes that the primary evolutionary function of dreaming is the rehearsal of complex social interactions and the maintenance of social bonds 505156. Human evolutionary success is deeply rooted in sophisticated social cooperation, alliance building, and the development of "Theory of Mind" - the cognitive capacity to attribute mental states, beliefs, and intentions to others 565758.
SST is supported by a robust statistical phenomenon known as the "Sociality Bias." While waking life experiences contain social interactions roughly 64% of the time, self-reported dream diaries feature social interactions over 80% to 83% of the time 5658. To test whether dreams merely reflect daily residue or possess an intrinsic drive to simulate sociality, researchers conducted an experiment where participants were subjected to prolonged social isolation. During the period of total social seclusion, their waking social interactions dropped to zero, yet their dream content remained highly social, showing almost no decrease in simulated interpersonal interactions 565859. This suggests that the brain's default state during sleep is to persistently generate social networks and practice interpersonal navigation regardless of immediate waking input 58.
Cross-Cultural Studies in Non-WEIRD Populations
Historically, dream research has been heavily skewed toward WEIRD (Western, Educated, Industrialized, Rich, and Democratic) populations, specifically North American and European university students 6061. To rigorously test evolutionary simulation theories, researchers must examine populations whose environments and social structures more closely approximate those of ancestral hunter-gatherers.
A landmark 2023 study published in Scientific Reports addressed this gap by analyzing the dream content of two forager communities - the BaYaka in the Republic of Congo and the Hadza in Tanzania - against Western control groups 45626364. Using natural language processing and structural dream analysis on 896 dreams from 234 individuals, researchers found profound cross-cultural differences in dream function 45626365.
The dreams of the BaYaka and Hadza exhibited significantly higher levels of physical threat compared to Western samples 456263. Crucially, however, these indigenous dreams frequently began with a life-threatening danger but ended with a resolution involving intense social support, community rescue, and coping mechanisms 6264. Despite the high threat content, the Hadza reported remarkably low levels of negative emotion in their dreams 4563. In contrast, Western dreams focused heavily on individualistic anxieties (e.g., the Canadian student sample during the COVID-19 pandemic displayed the highest anxiety dream content) and rarely featured community-based conflict resolution 456364.
These findings suggest an integrated model combining TST, SST, and emotional regulation: in highly cooperative forager societies, dreams effectively regulate emotion by simulating threats and immediately pairing them with prosocial resolutions, thus reducing anxiety through simulated communal catharsis 4562.
The following table summarizes the primary modern theories of dream function, their proposed mechanisms, and their primary empirical evidence:
| Dream Theory | Core Proposed Function | Key Neural/Cognitive Mechanism | Primary Empirical Evidence |
|---|---|---|---|
| Memory Consolidation (Replay) | Stabilization and integration of newly acquired information. | Hippocampal sharp-wave ripples replaying spatial/temporal firing sequences. | Task-related dreaming correlates with improved post-sleep performance (primarily NREM) 272832. |
| NEXTUP Model | Extraction of new knowledge and abstract rules. | Suppression of prefrontal logic allows exploration of weak semantic associations. | Dream bizarreness; failure of dreams to exactly replicate waking episodic memories 13335. |
| Sleep to Forget, Sleep to Remember (SFSR) | Depotentiation of autonomic distress associated with memories. | High limbic activity coupled with zero noradrenaline during REM sleep. | Decreased amygdala reactivity to negative stimuli post-sleep; efficacy of prazosin in PTSD 57940. |
| Threat Simulation Theory (TST) | Rehearsal of threat perception and evasion tactics. | Activation of fear-conditioning networks in a safe, paralyzed state. | High prevalence of fear, anger, and physical aggression in normative dream reports 452. |
| Social Simulation Theory (SST) | Maintenance of social bonds and practice of Theory of Mind. | Intrinsic drive to generate social networks and interpersonal interactions. | The "Sociality Bias"; persistence of social dreams during waking social isolation 5658. |
Objective Measurement Through Artificial Intelligence Decoding
The historical barrier to the scientific study of dreaming has been its total reliance on subjective, post-hoc verbal reports 66. Because the prefrontal cortex is suppressed during REM sleep, dream recall upon waking is notoriously fragile, highly susceptible to waking-life narrative restructuring, cognitive biases, and amnesia 1816. However, recent advancements at the intersection of neuroscience and artificial intelligence (AI) are making the objective decoding of visual dream content a reality, moving the field from theoretical speculation to empirical validation 66.
Methodologies of Functional MRI Dream Reconstruction
In 2024 and 2025, research teams, notably at the ATR Computational Neuroscience Laboratories in Kyoto, Japan, and various global consortiums, successfully utilized fMRI combined with deep learning algorithms to visually reconstruct dream content directly from the sleeping brain 676869.
The methodology relies on capturing the Blood Oxygen Level Dependent (BOLD) signal in the visual cortex while a subject is in REM sleep 6768. Because the visual cortex maps imagined or dreamed images using similar spatial and neural patterns as actual optical input, AI models (such as advanced diffusion models) can be trained on a subject's waking brain activity while they view known images from databases like the Natural Scenes Dataset (NSD) 6769.
When the subject subsequently sleeps in the fMRI scanner, the AI interprets the spontaneous REM BOLD signals and generates visual reconstructions. Current iterations of this technology, such as the "DreamConnect" dual-stream framework, integrate fMRI with natural language prompts to achieve approximately 60% to 70% accuracy in identifying specific categories of dreamed objects, people, and spatial layouts 686970. During these experiments, subjects were awakened over 200 times during REM sleep to provide verbal reports, allowing the researchers to continuously refine the AI's mapping dictionary 6870.
Clinical and Neuroscientific Implications
While the generated images currently remain rudimentary, blurry, and require subject-specific training models, they represent a monumental leap in the field of "oneirography" (dream recording) 676870. By establishing a non-verbal, objective window into the sleeping mind, AI decoding bypasses the bottleneck of human memory recall.
This technology holds profound implications for clinical psychiatry and cognitive enhancement. It may soon allow clinicians to directly observe the emotional and cognitive patterns of patients suffering from severe PTSD or nightmare disorders, providing an objective measure of whether emotional depotentiation is occurring 666870. Furthermore, it offers a potential communication pathway for "locked-in" patients unable to verbally articulate their internal states, and promises to definitively resolve ongoing debates regarding the exact nature of memory replay versus abstract reconstruction during sleep 666870.
Conclusion
The scientific inquiry into dreaming during REM sleep has definitively moved beyond the binary debate of whether dreams are meaningful psychological symbols or random physiological noise. The synthesis of over half a century of neurobiological, cognitive, and evolutionary research indicates that dreaming is a highly orchestrated, biologically adaptive mechanism that sits at the critical intersection of memory architecture and emotional homeostasis.
The bizarreness of dreams, once dismissed as neural error, is now recognized as a functional necessity. Facilitated by a unique neurochemical profile - characterized by high acetylcholine and the total suppression of noradrenaline and serotonin - the brain deliberately suspends logical prefrontal oversight. This allows for the exploration of weak semantic associations and novel memory combinations, driving creative problem solving and knowledge extraction as proposed by the NEXTUP model. Concurrently, this exact neurochemical environment strips episodic memories of their severe autonomic arousal, preserving the factual data while discarding the painful visceral response, fulfilling the regulatory function outlined by the Sleep to Forget, Sleep to Remember hypothesis.
Furthermore, evolutionary data and cross-cultural analyses demonstrate that this simulated environment serves as a vital neurocognitive training ground. By persistently generating complex social interactions and resolvable threatening scenarios, the dreaming brain prepares the individual for waking survival, reinforcing the communal bonds and threat-evasion tactics necessary for human flourishing. As technological breakthroughs in artificial intelligence and fMRI continue to allow for the direct visual decoding of the sleeping brain, our fundamental understanding of consciousness, memory plasticity, and emotional resilience will inevitably expand, cementing the dream state as one of the brain's most sophisticated and essential cognitive functions.