Neuroscience of Dreaming and Memory
The phenomenon of human dreaming has transitioned from a subject of purely philosophical and psychoanalytic inquiry into a rigorous domain of cognitive neuroscience. Historically constrained by the subjective nature of self-reporting and the limitations of early neuroimaging, the study of the sleeping brain now leverages high-density electroencephalography (hd-EEG), functional magnetic resonance imaging (fMRI), simultaneous multimodal imaging, and advanced artificial intelligence. Current research indicates that dreaming is not merely an epiphenomenon of sleep but a highly structured neurological process that supports memory consolidation, emotional regulation, and cognitive simulation. By mapping the contours of sleep consciousness, researchers are uncovering the distinct cortical networks and metabolic states responsible for our nightly immersion into virtual sensory realities.
Neurobiological Architectures of the Dreaming State
The foundational paradigm of twentieth-century sleep research heavily conflated dreaming exclusively with rapid eye movement (REM) sleep. This assumption emerged largely due to the wake-like, high-frequency "activated" EEG patterns characteristic of the REM state, combined with the profound motor atonia that prevents the acting out of dreams 12. However, subsequent empirical investigations have dismantled this unified-state theory, demonstrating that vivid, narrative dreaming occurs throughout various sleep stages, including non-rapid eye movement (NREM) sleep 13. Equating REM sleep strictly with dreaming is now considered a foundational error in neuroscientific logic; while the two phenomena frequently overlap, they operate via dissociable physiological mechanisms 34.
Cortical Activation and State Disassociation
The neurobiological signature of dreaming is highly localized rather than globally uniform. During REM sleep, brain activity significantly diverges from both wakefulness and NREM states. Modern neuroimaging via positron emission tomography (PET) and fMRI reveals a distinct pattern of regional activation and deactivation. During REM dreaming, there is pronounced hyperactivation in the limbic system - particularly the amygdala and hippocampal formation - alongside heightened activity in the anterior cingulate cortex, the basal forebrain, and the temporo-occipital cortices 256. This deep limbic and temporal engagement correlates strongly with the emotional intensity, episodic memory retrieval, and visual vividness that characterize dream content.
Conversely, specific regions undergo profound suppression. The dorsolateral prefrontal cortex (DLPFC), the precuneus, and the inferior parietal cortex exhibit marked deactivation during REM sleep 277. The suppression of the DLPFC - a region integral to executive function, working memory, and logical reasoning - provides a structural explanation for the hallmark features of dream phenomenology. Because executive logic is muted, the dreaming mind exhibits hyper-associative logic, diminished self-reflective awareness, and an unquestioning acceptance of bizarre or impossible events 57. Visualizing the spatial topography of the sleeping brain highlights this dichotomy: the posterior sensory regions and deep limbic structures "heat up" to drive the visual and emotional intensity of the dream, while the anterior frontal networks "cool down," effectively removing the brain's reality-testing filters.
To illustrate the contrasting metabolic and hemodynamic landscapes of the brain across different states of arousal, the following table summarizes the relative regional cerebral metabolic rates for glucose (rCMRglc) and blood flow dynamics:
| Brain Region | Primary Functional Role | Activity in REM Sleep | Activity in NREM Sleep | Activity in Wakefulness |
|---|---|---|---|---|
| Amygdalo-Hippocampal Complex | Emotion processing, episodic memory consolidation | Highly Active (Hyperactivated) | Moderately Active (Bursts) | Baseline Active |
| Dorsolateral Prefrontal Cortex (DLPFC) | Executive function, logic, working memory | Suppressed (Deactivated) | Suppressed | Highly Active |
| Anterior Cingulate Cortex | Affect regulation, conflict monitoring | Highly Active | Variable | Baseline Active |
| Thalamus | Sensory relay, cortical arousal | Active | Suppressed | Highly Active |
| Temporo-Occipital Cortices | Visual processing, spatial environment construction | Highly Active | Decreased (Global) | Active |
| Brainstem (Pontine Tegmentum) | REM generation, motor atonia | Highly Active | Active | Baseline Active |
The Posterior Hot Zone and Conscious Mentation
The discovery that robust dreaming occurs during NREM sleep - a state dominated by low-frequency slow-wave activity (SWA) - initially presented a paradox for sleep neuroscience 134. Resolving this paradox required shifting the analytical focus from global brain states to highly localized cortical networks.
Using high-density EEG paired with serial awakening paradigms, researchers identified a specific neural correlate of dreaming that operates independently of the broader sleep stage: the "posterior hot zone" 198. Encompassing the medial and lateral occipital lobe, the precuneus, and the posterior cingulate gyrus, this parieto-occipital region demonstrates a localized decrease in low-frequency power (specifically the 1 - 4 Hz delta band) whenever a subject reports having experienced a dream 111. In neurophysiological terms, this localized decrease in low-frequency SWA signifies heightened localized cortical activation amidst a globally resting brain.
Crucially, monitoring the activation of the posterior hot zone allows researchers to accurately predict the presence of a conscious dream experience in real-time, whether the subject is currently in REM or NREM sleep 18. Furthermore, high-frequency activity (beta and gamma bands) within this hot zone correlates with highly specific dream contents. For instance, high-frequency activity in face-processing or spatial-processing cortical areas directly maps onto the dreamer's subsequent reports of seeing faces or moving through specific spatial environments 8.
The isolation of this hot zone also clarifies the phenomenon of "white dreaming" - instances where an individual awakens with the absolute certainty of having dreamed but possesses no recall of the specific narrative or visual content 11. Neuroimaging indicates that the posterior hot zone remains highly activated during white dreaming, confirming it as a fundamental marker of the conscious experience itself, rather than merely the memory encoding or retrieval of that experience 11.
The Default Mode Network and Spontaneous Thought
The cortical regions comprising the posterior hot zone overlap significantly with the brain's Default Mode Network (DMN), a large-scale interconnected network that includes the medial prefrontal cortex, posterior cingulate cortex, and angular gyrus 910. During wakefulness, the DMN is primarily active during states of rest, mind-wandering, autobiographical memory recall, and self-referential simulation 10.
During sleep, the DMN becomes largely uncoupled from external sensory input systems (such as the dorsal attentional network) and executive control pathways, allowing it to autonomously generate the spontaneous, internally directed simulations characteristic of dreams 1011. Contemporary neuroscience increasingly suggests that dreaming may be a specialized operational mode of the DMN, operating untethered from waking constraints. Clinical lesion studies corroborate this deep connection; damage to key DMN hubs, particularly the ventromedial prefrontal cortex (vmPFC), can result in the complete and permanent cessation of dreaming - a neurological condition known as Charcot-Wilbrand syndrome 1011. Furthermore, the hyper-associative state of the DMN during dreaming shares neurochemical and functional similarities with the DMN activity observed during deep meditation, psychedelic experiences, and even the source-monitoring failures typical of psychosis 911.
Hemodynamic and Metabolic Transitions
Recent methodological advances utilizing simultaneous EEG, functional PET (fPET), and fMRI have provided an unprecedented look at the complex transition dynamics of the sleeping brain. As the human brain descends from wakefulness into deep NREM sleep, there is a tightly coupled temporal progression of global hemodynamics and glucose metabolism 1213. Energy use and overall metabolic consumption decline markedly alongside a drop in EEG arousal dynamics.
However, despite the overall reduction in metabolic demand, blood flow remains highly dynamic and spatially structured 1314. Specifically, researchers have identified a ~0.02-Hz oscillating sensorimotor network that remains highly active and metabolically demanding throughout NREM sleep. This network allows the sleeping brain to retain baseline environmental reactivity and process physical positioning, even as higher-order cognitive networks and the core components of the DMN undergo profound functional suppression and reorganization 1213. Concurrently, this metabolic quieting facilitates an increase in cerebrospinal fluid flow, underscoring sleep's parallel role in neuro-metabolic waste clearance 13.
Memory Consolidation and the Processing of Emotion
A primary, evolutionarily conserved biological function of sleep is the consolidation of memory - the intricate process by which fragile, newly encoded memory traces are stabilized, integrated into pre-existing long-term cortical networks, and protected from waking interference. Dreaming is increasingly understood as the conscious, phenomenological manifestation of this underlying neuro-computational process.
Slow Oscillation and Spindle Coupling
During NREM sleep, the mechanical transfer and stabilization of declarative and spatial memories are driven by the precise phase-amplitude coupling of neocortical slow oscillations (SOs) and thalamic sleep spindles 1516. Spindles - brief, rapid bursts of 11 - 15 Hz electrical activity - nest within the up-state trough of the slow oscillations. This synchronization facilitates the highly efficient transfer of declarative memory representations from their temporary storage in the hippocampus to distributed, long-term networks across the neocortex 1617. The strength and precision of this SO-spindle coupling directly predict an individual's next-day memory retention and represent a critical mechanism for integrating new information into established schemas 16.
Theoretical Models of REM Memory Processing
While NREM sleep primarily stabilizes factual, declarative, and spatial memories, REM sleep is uniquely implicated in the processing of emotionally arousing and traumatic experiences. Two prominent, complementary theoretical frameworks attempt to explain how REM sleep achieves this: the "Sleep to Forget, Sleep to Remember" (SFSR) hypothesis and the "REM Refine and Rescue" (RnR) hypothesis.
The SFSR hypothesis posits that REM sleep serves a dual, seemingly paradoxical role in memory transformation. It suggests that during REM, the brain consolidates the core episodic and declarative details of an emotional memory (the "remember" function) while actively stripping away the visceral, autonomic, and stress-inducing affective charge originally associated with the event (the "forget" function) 151819. This affective decoupling is neurobiologically facilitated by the unique neurochemical environment of REM sleep. The state is characterized by a profound reduction in central aminergic activity - specifically the suppression of locus coeruleus-derived norepinephrine - combined with sustained hippocampal theta oscillations 19. In this low-adrenergic, low-stress environment, emotional memories are reactivated and re-encoded without triggering the corresponding autonomic stress response. Over successive sleep cycles, this leads to decreased amygdala reactivity and enhanced top-down prefrontal connectivity upon waking, effectively neutralizing the trauma of the memory 1920. Comparing the memory pre-sleep to post-sleep, the original memory trace enters sleep tightly bound to a high emotional charge; through the low-noradrenaline processing of REM, the memory emerges preserved in detail but significantly reduced in emotional intensity.
The RnR hypothesis builds upon this concept by focusing on the signal-to-noise ratio within memory representations 19. Rather than viewing forgetting merely as affective decoupling, the RnR model suggests that REM sleep actively "refines" memories by enhancing the essential, core nodes of a representation while actively inhibiting or pruning superfluous, neutral, or noisy details. Simultaneously, it "rescues" weak or degraded memory traces that have suffered from retroactive interference during waking hours, preventing them from being permanently forgotten 19. Under the RnR framework, an initially weak or noisy memory enters the REM cycle, undergoes targeted signal enhancement and noise elimination, and emerges as a strengthened, core memory trace.
Targeted Memory Reactivation Dynamics
Experimental validation of sleep's active role in memory transformation relies heavily on Targeted Memory Reactivation (TMR). In TMR experimental paradigms, sensory cues - most commonly unique odors or auditory tones - are explicitly paired with specific learning tasks or emotional stimuli while the subject is awake. Later, while the subject sleeps, these identical cues are covertly re-played during specific polysomnographically verified sleep stages. This external cueing artificially triggers the reactivation of the targeted memory trace in the sleeping brain 1821.
Meta-analyses demonstrate that TMR applied during NREM sleep (specifically Stage 2 and slow-wave sleep) reliably yields moderate-to-strong enhancements in declarative memory, motor skill acquisition, and spatial discrimination 212223. NREM TMR triggers localized increases in theta and spindle oscillations, directly facilitating cortical integration and improving post-sleep test performance for the cued items 22.
The application of TMR during REM sleep, however, yields fundamentally different outcomes. While REM TMR does not consistently improve the basic recall or retention of declarative facts, it profoundly modulates emotional regulation and physiological arousal 2024. Studies indicate that when odors associated with highly negative or distressing images are replayed specifically during REM sleep, subjects awaken with significantly reduced subjective arousal ratings when re-exposed to those images 2024.
Furthermore, EEG and fMRI measurements validate this subjective habituation. REM TMR selective enhances the Late Positive Potential (LPP) amplitude during post-sleep emotional processing, and significantly decreases blood-oxygen-level-dependent (BOLD) activity in the brain's salience network - including the anterior insula, dorsal anterior cingulate cortex (dACC), and the amygdala 202224. These physiological alterations provide robust empirical support for the SFSR hypothesis, indicating that REM sleep is specifically optimized for the disarming of emotional trauma and the recalibration of the brain's threat-response systems 1820.
Evolutionary Functions and Simulation Hypotheses
Beyond physiological maintenance, the question of why the brain constructs elaborate, immersive, and often bizarre virtual realities during sleep remains heavily debated. Evolutionary psychology and cognitive neuroscience have proposed several simulation theories, suggesting that the virtual environments of dreams conferred distinct adaptive and survival advantages to early humans.
Threat Simulation Theory (TST)
Formulated by cognitive neuroscientist Antti Revonsuo, the Threat Simulation Theory posits that dream consciousness is an ancient biological defense mechanism, evolutionarily selected for its unique capacity to repeatedly simulate threatening events 282526. According to TST, the brain utilizes the safe, physically paralyzed state of REM sleep to rehearse threat perception and evasive motor maneuvers. By generating simulated predatory or environmental threats, the brain sharpens waking survival instincts, reflex speeds, and threat-recognition heuristics without exposing the organism to genuine physical risk 2728.
Evidence supporting TST is mixed but compelling in contexts of severe environmental stress. A major predictive tenet of TST is that real-world trauma and danger should hyper-activate the threat simulation system. A landmark 2005 study supported this hypothesis, demonstrating that severely traumatized Kurdish children living in highly dangerous, conflict-ridden environments reported significantly more dreams containing severe, realistic threats compared to a control group of non-traumatized Finnish children 2829. Furthermore, a large-scale quantitative analysis of 212 recurrent adult dreams found that 66% contained one or more explicit threats, with the dreamer frequently taking logical, defensive actions to evade the danger 252830.
However, contradictory data complicates the universal applicability of the theory. A 2008 epidemiological study comparing populations in a high-crime, dangerous area of South Africa with those in a low-crime area in Wales found that the South African cohort, despite significantly higher real-world threat exposure, actually reported fewer threat-based dreams 28. Additionally, in the aforementioned recurrent dream study, fewer than 15% of the simulated threats depicted realistic survival scenarios that would actually improve evolutionary fitness. Most critically, dreamers rarely succeeded in escaping the threat - fleeing successfully in fewer than 2% of reports - suggesting that if dreaming is a training simulator, it is remarkably prone to failure 2528.
Social Simulation Theory (SST)
Parallel to TST, the Social Simulation Theory suggests that the primary evolutionary function of dreaming is the rehearsal, management, and resolution of complex social challenges 283531. Humans are a hyper-social species; evolutionary fitness and reproductive success rely heavily on social cohesion, bond maintenance, empathy, and the successful navigation of complex tribal hierarchies. SST argues that the universal prevalence of social interactions in dreams reflects a specialized virtual-reality mechanism designed explicitly to train social and emotional skills 31.
Proponents of SST critique TST on several grounds, primarily for failing to account for the 25% to 35% of dreams that contain absolutely no physical threats, and for downplaying the overwhelming abundance of social perception, interpersonal conflict, and emotional bonding that dominates routine sleep mentation 35. By framing the dream space as a "social simulator," SST accounts for the highly interpersonal nature of most dreams. This aligns seamlessly with neurobiological findings that the regions activated during dreaming strongly overlap with the waking "social brain" network, and supports the clinical observation that REM sleep actively downregulates emotional reactivity to waking social stressors 2835.
Reevaluation of Psychoanalytic Frameworks
For decades, the neurobiological consensus - led by J. Allan Hobson and Robert McCarley's Activation-Synthesis model in the 1970s - dismissed the classical psychoanalytic models of Sigmund Freud and Carl Jung as prescientific and fundamentally incorrect 33732. The Activation-Synthesis model posited that dreams were essentially "garbage" - meaningless, hallucinatory epiphenomena resulting from the forebrain's desperate attempt to synthesize and make narrative sense of random, chaotic pontine signals firing from the brainstem during REM sleep 3732. Under this model, the content of a dream held no intrinsic psychological meaning.
However, modern neuropsychoanalysis, championed by Mark Solms, has utilized advanced clinical lesion studies and fMRI to challenge Activation-Synthesis and partially vindicate early psychoanalytic concepts 334034. Solms discovered a critical double dissociation between REM sleep and dreaming. Patients with extensive lesions in the brainstem can lose the capacity for REM sleep entirely, yet they continue to experience vivid, narrative dreams 4034. Conversely, patients with damage to the ventromedial forebrain and its underlying white matter - specifically the dopaminergic pathways associated with motivation, emotion, and instinctual "wanting" - lose the ability to dream entirely, while their physiological REM sleep cycles remain perfectly intact and cyclical 40.
This neuroanatomical separation irrevocably indicates that dreaming is driven not by random brainstem activation, but by the forebrain's motivational and emotional command centers. This provides a modern, biologically grounded analog to Freud's highly contested theory that dreams are fundamentally driven by desire, emotion, and instinctual drives (wish fulfillment) seeking resolution while executive censorship is offline 374034.
At the outermost theoretical edges of evolutionary psychology, frameworks like Epic Cognition Theory (ECT) propose even more radical interpretations, suggesting that the "schizoid" withdrawal of sleep allows the deep, symbolic processing required for major evolutionary leaps. While highly speculative, such frameworks view dreaming not as a passive byproduct, but as an active, symbolic engine for biological and cognitive adaptation 3536.
| Theory / Model | Primary Function of Dreaming | Neurological / Evolutionary Basis | Status in Modern Neuroscience |
|---|---|---|---|
| Activation-Synthesis | None (Epiphenomenon) | Random brainstem firing synthesized by the forebrain | Largely discredited due to double-dissociation of REM/Dreaming |
| Threat Simulation Theory | Rehearsal of survival skills | Rehearsal of ancestral physical threats in a paralyzed state | Partially supported, but challenged by lack of escape success |
| Social Simulation Theory | Rehearsal of social interactions | Evolution of complex social hierarchies and tribal bonding | Strongly supported by DMN and "social brain" overlap |
| Neuropsychoanalytic Model | Processing of motivational drives | Forebrain dopaminergic pathways seeking emotional resolution | Gaining empirical support via lesion and fMRI studies |
Artificial Intelligence in Dream Analysis and Decoding
The intersection of artificial intelligence, machine learning, and neuroimaging is precipitating a paradigm shift in dream research. Large language models (LLMs), diffusion models, and massive foundation models are moving the field away from speculative psychological interpretation and toward the quantitative, algorithmic decoding of physiological data and visual sleep mentation.
Foundational Models for Sleep Physiology
In January 2026, researchers at Stanford University introduced SleepFM, a pioneering multimodal foundation model designed to extract predictive health insights directly from the architecture of sleep 4437383940. Trained on nearly 600,000 hours of dense polysomnography data collected from 65,000 participants, SleepFM acts as an advanced pattern-recognition system for the sleeping body's general physiology 38.
Rather than assessing isolated metrics in a vacuum, SleepFM utilizes a novel "leave-one-out contrastive learning" technique. It ingests simultaneous, complex data streams - EEG (brainwaves), ECG (heart activity), EMG (muscle tension), and respiratory airflow - hides one modality, and trains the AI to reconstruct the missing data based on the remaining signals 3840. This rigorous training forces the model to learn the deep, interconnected physiological language of sleep.
Remarkably, by compressing these massive data matrices into compact numerical forms, SleepFM successfully identifies physiological sleep anomalies that predate clinical waking symptoms by years. The model accurately predicts over 130 health conditions, achieving exceptionally high concordance indices (C-index) for severe pathologies, including Parkinson's disease (0.89), prostate cancer (0.89), breast cancer (0.87), dementia (0.85), and heart attacks (0.81) 3840. SleepFM demonstrates that sleep mentation and autonomic physiology serve as a highly dense, largely untapped biomarker for long-term systemic health 39.
Functional MRI Decoding and Visual Reconstruction
Historically, the concept of directly decoding the visual content of a dream from brain activity was deemed impossible due to the abstract, noisy nature of neurophysiological signals. However, the advent of generative AI - particularly Stable Diffusion models integrated seamlessly with fMRI data - has made "visual brain decoding" a tangible reality 41425143.
In July 2025, an international consortium of researchers introduced DreamConnect, a breakthrough brain-to-image AI system capable of translating fMRI signals directly into editable visual imagery 424445. The system utilizes a dual-stream diffusion framework that bridges the gap between raw neural blood flow and semantic image generation.
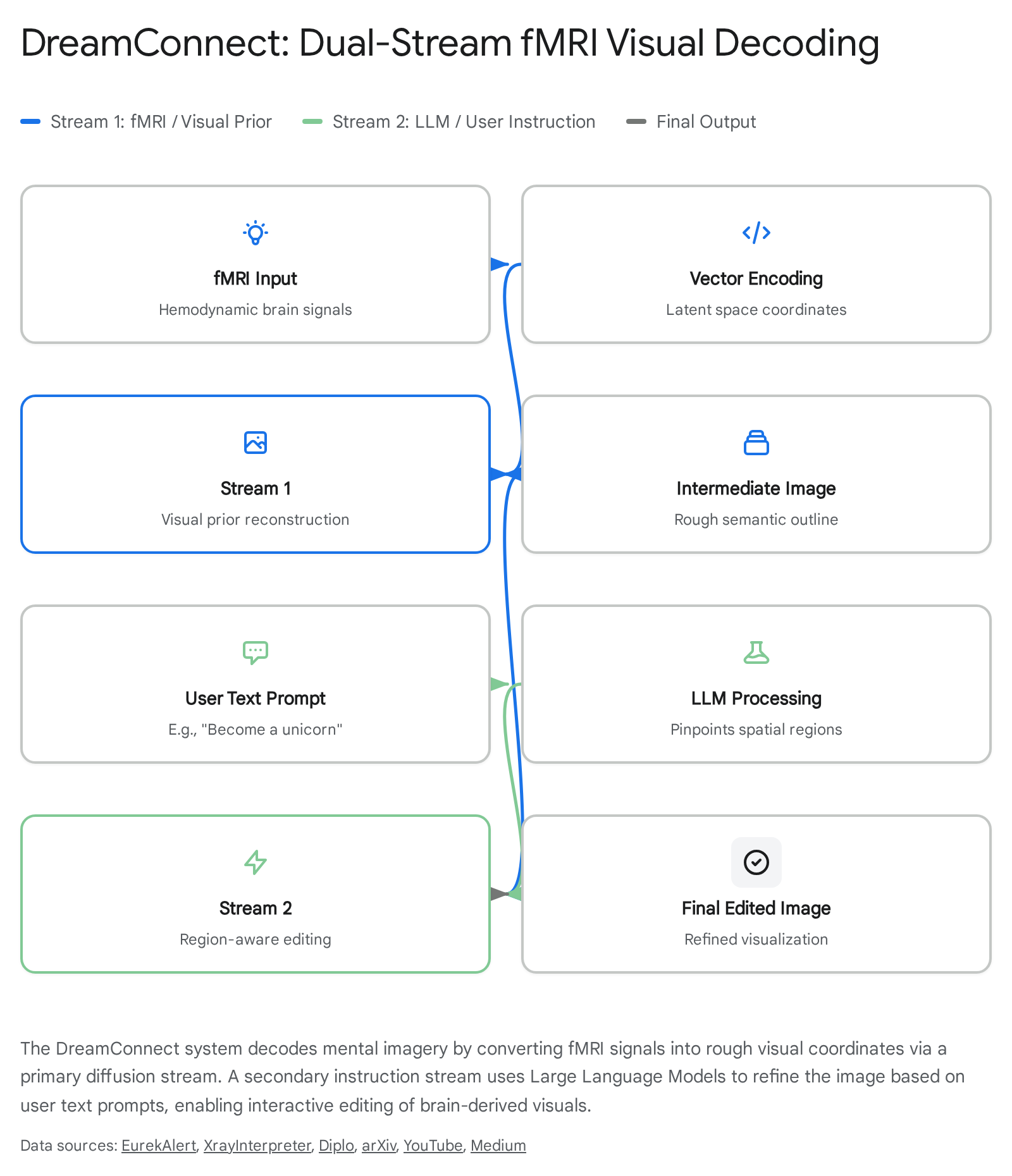
When a subject visualizes a concept in an MRI scanner, the first stream of DreamConnect translates the compressed vector of their visual cortex activity into rough latent-space coordinates, generating an intermediate semantic outline of the image 4251. The true breakthrough of the system lies in its interactive, instruction-based editing. A parallel stream integrates natural language prompts via LLMs to perform region-aware editing on the decoded brain signals. If a subject visualizes a horse, but the decoded fMRI signal is ambiguous, the system can accept the verbal prompt "make it a unicorn." The AI will then selectively alter only the relevant spatial features of the mental image while meticulously preserving the raw neural outline provided by the brain 424445. By aligning OpenAI's Contrastive Language-Image Pre-training (CLIP) embeddings with fMRI time-series data, systems like DreamConnect represent a fundamental transition from passive brain monitoring to active, real-time collaboration between human imagination and machine rendering 41454647.
Semantic Analysis and the Debunking of Dream Symbolism
Beyond visual decoding, advanced Natural Language Processing (NLP) is being applied to massive databases of written dream reports, such as the DreamBank repository. These computational analyses are systematically debunking the utility of traditional "dream dictionaries" - popular books and websites claiming that specific symbols (e.g., losing teeth, seeing a snake) yield absolute, universal meanings 485849.
NLP analysis proves conclusively that dream semantics are highly idiosyncratic, deeply personal, and lack absolute universal symbolism 4950. When analyzing the perplexity scores - a quantitative metric of text predictability and sequence entropy - of dream reports against standard texts like Wikipedia, LLMs actually find dream reports surprisingly easier to model and predict 51. The semantic structure of dreams is not chaotic or random; it follows distinct architectural patterns heavily influenced by the dreamer's waking life, personality, and environment.
For instance, semantic mapping shows that dreams are heavily enriched with perceptual and spatial language compared to waking reports, which heavily favor metacognitive and thought-based vocabulary 52. Furthermore, dream content is demonstrably altered by macro-societal events. NLP analysis of dreams collected during the COVID-19 pandemic revealed heightened emotional intensity and statistically significant increases in references to physical constraints, lockdowns, and spatial limitations 505253. As psychological adaptation to the pandemic progressed over subsequent years, these constraints faded proportionally from the semantic dream space. This confirms that dreaming is an active, dynamic process of memory and emotional synthesis, heavily tethered to both individual psychological traits (such as a propensity for waking mind-wandering) and undeniable environmental realities 5053.
Subjective Sleep Quality and the Phenomenology of Dreaming
The integration of advanced neuroimaging and detailed self-reporting has also begun to clarify the relationship between the subjective experience of dreaming and our perception of sleep quality. Historically, deep, restorative sleep was thought to correlate exclusively with dreamless, slow-wave NREM sleep, while the wake-like brain activity of REM sleep was viewed as a potential disruptor of perceived restfulness.
Perceptions of Sleep Depth
Recent studies have challenged this dichotomy. Research analyzing EEG recordings alongside serial awakenings has shown a counterintuitive relationship between vivid dreaming and the subjective feeling of having had a "good night's sleep" 5455. When individuals experience perceptually immersive, emotionally intense, and vivid dreams, they are significantly more likely to report feeling that their sleep was deep and restorative upon waking 5455.
Conversely, dreams that are abstract, highly reflective, or contain metacognitive awareness (resembling waking thought) are associated with feelings of shallow, fragmented sleep 54. Furthermore, sleep misperception - the phenomenon where individuals feel they were awake even when EEG confirms they were asleep - occurs most frequently during early, dreamless stages of NREM sleep 55. This suggests that the immersive virtual reality of vivid dreaming plays a functional role in sleep architecture: by deeply engaging the brain's internal DMN and sensory networks, dreaming effectively disconnects the mind from the external environment, thereby securing the depth and continuity of sleep 54.
Conclusion
The neuroscience of dreaming has moved conclusively past the reductionist view that sleep mentation is mere biological noise or random hallucination. Facilitated by precise regional brain activation - such as the hyperactive limbic system and the posterior hot zone - and guided by the intricate coupling of slow oscillations and sleep spindles, dreaming serves as an essential cognitive and emotional utility. Whether refining fragile memories, stripping traumatic experiences of their autonomic physiological charge, or simulating social interactions to prepare the waking mind, the dreaming brain performs vital maintenance on human consciousness. As multimodal foundation models and fMRI-to-image decoding technologies mature, the scientific community is gaining unprecedented, quantitative access to this internal theater. This technological evolution transforms our understanding of sleep from a passive state of physiological rest into a highly active, computational process required for psychological resilience and cognitive survival.