How Your Brain Makes Decisions Under Uncertainty
When the human brain faces uncertainty, it acts as a biological statistician, accumulating sensory and memory evidence in a rapid race toward a critical decision threshold. This complex calculation relies on a highly synchronized, brain-wide neural network - spanning the prefrontal cortex, insula, and basal ganglia - that continuously measures the gap between expected and actual outcomes. Ultimately, decision-making is not an isolated event in a single "choice center," but a dynamic cascade of electrical signals that updates internal models and drives behavioral adaptation.
The Illusion of Immediate Choice: Breaking Down the Process
Every day, humans make thousands of decisions without complete information. Whether choosing a career path or guessing which lane of traffic will move faster, the outcomes of these choices are inherently unpredictable. For decades, the internal computation behind these choices was treated by neuroscientists as an impenetrable "black box." The subjective experience of making a decision feels instantaneous, but beneath the surface of conscious awareness, a complex and measurable biological algorithm is executing.
Recent breakthroughs at the intersection of mathematical modeling and cognitive neuroscience have illuminated this process, transforming abstract psychological concepts into observable physiological events. Researchers have discovered that the brain does not simply flip a coin when faced with ambiguity. Instead, it engages in a rigorous, time-pressured process of evidence accumulation, weighing probabilities, estimating risks, and preparing motor responses simultaneously. This process is governed by specific neurotransmitters, precise electrical brain waves, and anatomical structures that have evolved to balance speed against accuracy.
The Drift Diffusion Model: A Biological Race for Evidence
To understand how the brain resolves ambiguity, neuroscientists frequently rely on the Drift Diffusion Model (DDM). The DDM is a mathematical blueprint that conceptualizes every decision under uncertainty as a literal race for evidence 1.
In this model, every available option acts as a runner at a starting line. As the brain gathers evidence from the external environment, internal memories, and sensory inputs, it pushes one of the "runners" forward. The speed at which this evidence accumulates is referred to as the "drift rate" 1. Simultaneously, the brain establishes a "decision threshold," which acts as the finish line. The first option to accumulate enough evidence to cross that threshold dictates the final choice 1.
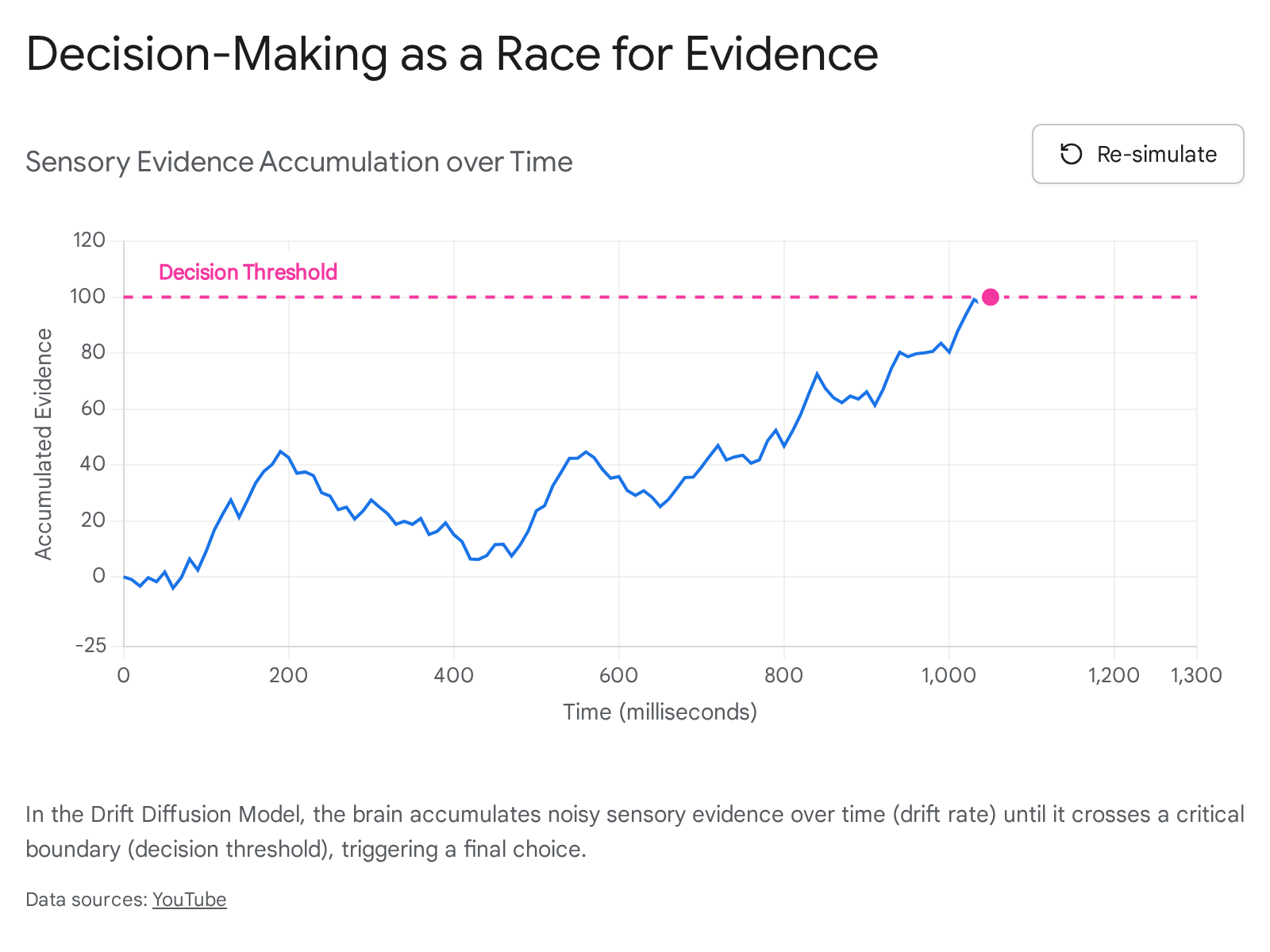
Neuroscientists have mapped this abstract mathematical model directly onto specific neural hardware. The lateral intraparietal area (LIP) functions as the scorekeeper, constantly tallying the incoming evidence and driving the drift rate 1. Concurrently, the basal ganglia and the subthalamic nucleus (STN) define the threshold, determining exactly how much evidence is required to commit to an action 1. Once this threshold is crossed, a "go" signal is transmitted to the sensory-motor cortex, which executes the physical action 1.
Reading the Brain's Electrical Ticker
By monitoring the brain's electrical chatter using electroencephalography (EEG), researchers can observe the mechanisms of the Drift Diffusion Model in real-time. Specific brain waves map perfectly onto the model's theoretical components 1. A signal known as the N200 directly reflects the drift rate - the speed and volume of evidence coming in 1. Following this, another signal called the P300 tracks the approach to the decision threshold, effectively measuring how close the brain is to finalizing the choice 1.
Why "Noise" is a Feature, Not a Bug
Interestingly, the electrical signals involved in this process are not perfectly smooth. They exhibit a natural wobble or randomness in neural firing, commonly referred to as "noise" 1. Rather than being a flaw in the biological system, this noise is a critical feature. It explains why a person might take slightly longer to make the exact same choice on two different days, even under identical conditions 1. This natural variability allows the nervous system to remain flexible, preventing the brain from falling into overly rigid, deterministic loops. In fact, this same algorithm - accumulating noisy evidence up to a threshold - governs not just abstract choices, but also how the nervous system processes pain, integrates sensory signals, and perceives the passage of time 1.
Mapping the Uncertainty Network: Cortical and Subcortical Players
When information is sparse or contradictory, the brain cannot rely on automatic habits. Instead, it must engage an extensive, specialized neural network encompassing both cortical and subcortical structures 1. A comprehensive meta-analysis synthesizing 76 functional magnetic resonance imaging (fMRI) studies, involving over 4,186 participants, has identified nine distinct activation clusters dedicated exclusively to processing uncertainty 12.
The prefrontal cortex (PFC), located at the front of the brain, acts as the primary executive center. It is responsible for higher-order cognitive functions such as planning, assessing risks and rewards, and formulating complex strategies 34. Within this region, the dorsolateral prefrontal cortex (dlPFC) evaluates options based on past experiences and future goals, serving as a critical hub for working memory 14.
However, the PFC does not act alone. Deeper limbic structures, particularly the amygdala, process the emotional weight of a decision. When a choice carries significant risk, the amygdala triggers emotional responses - such as fear or anxiety - that strongly influence the PFC to prioritize safety and risk aversion 34.
Neuroimaging reveals a profound functional specialization across the two hemispheres of the brain when navigating uncertainty, with different regions handling distinct aspects of the cognitive and emotional workload.
| Brain Region | Hemispheric Specialization | Primary Role in Decision-Making |
|---|---|---|
| Anterior Insula | Left: Reward evaluation Right: Learning and cognitive control |
Appraises risks and modulates emotional responses to unpredictable environments 12. |
| Inferior Frontal Gyrus | Left: Motor planning Right: Impulse control |
Governs the execution or suppression of physical actions once a decision is formulated 2. |
| Dorsal Anterior Cingulate Cortex (dACC) | Central / Medial | Evaluates the ongoing reliability of the current decision-making strategy 1. |
| Striatum | Subcortical (Bilateral) | Integrates cognitive and emotional input to guide decisions based on anticipated rewards 3. |
The Prefrontal Cortex as the Executive Architect
The division of labor within the frontal lobes highlights the complexity of decision-making. The meta-analysis shows that cognitive processes are largely handled by the inferior parietal lobule and the inferior frontal gyrus, while emotional and motivational appraisals are heavily tied to the anterior insula 2. By functionally splitting the workload, the brain can simultaneously calculate the statistical probability of a positive outcome while actively suppressing the impulse to make a hasty, fear-driven choice.
Dopamine's True Role: The Reward Prediction Error
A pervasive misconception in popular science is that dopamine is simply the brain's "pleasure chemical," released exclusively to make humans feel happy when they receive a reward 5. In reality, the neuroscience of decision-making reveals a much more sophisticated computational mechanism. Most dopaminergic neurons in the midbrain are not triggered by the reward itself. Instead, they are triggered by the mathematical difference between the expected reward and the actual reward - a concept known in computational neuroscience and reinforcement learning as the "reward prediction error" (RPE) 56.
Beyond the Pleasure Myth
When an individual makes a decision under uncertainty, the brain creates a subconscious expectation of the outcome. If the result of the decision is better or more valuable than expected, the individual experiences a positive prediction error. This results in a sharp spike of dopamine 56. This dopamine release does not merely create a fleeting sense of joy; it serves as a powerful, permanent teaching signal. The phasic firing of dopamine neurons endows the preceding cues with subjective value, updating the brain's internal models so that the individual is more likely to repeat that behavior in the future 78.
Positive vs. Negative Prediction Errors
The reward prediction error system is bidirectional, allowing the brain to learn flexibly from both success and failure 7. If a person perfectly predicts a reward, dopamine neurons remain at their baseline activity level 6. No matter how objectively great the reward is, if there is zero uncertainty and zero prediction error, there is no extra dopamine release 5.
Conversely, if a choice is made and the outcome is worse than expected, it generates a negative prediction error. This suppresses the firing of dopaminergic neurons in the midbrain 57. This suppression acts as an inhibitory signal, instructing the brain to reduce the value associated with the cue that led to that poor decision, thereby preventing the individual from making the same mistake again 79.
The Boredom of Certainty
The reliance on prediction errors highlights why uncertainty is fundamentally necessary for sustained motivation. In a hypothetical environment entirely devoid of uncertainty - where an individual could predict every outcome flawlessly without any subjective bias - the brain would be starved of positive reward prediction errors 5. Without these dopamine spikes, life and learning would become incredibly monotonous. The joy of winning a game, for instance, largely stems from the fact that winning is not guaranteed; the resolution of that uncertainty triggers the dopamine response that makes the activity inherently rewarding 5.
From Abstract Goals to Concrete Actions: The Cascade Model
Deciding what to do is only half the battle; the brain must also determine how to execute the decision. The "cascade model" of prefrontal executive function provides a compelling theoretical framework for how the brain translates an uncertain, abstract goal into a concrete sequence of physical actions 110.
Hierarchical Control in the Prefrontal Cortex
According to the cascade model, control processes within the prefrontal cortex are organized hierarchically based on their level of abstraction 10. Information transfer is asymmetric. High-level, abstract policies (for example, "I need to prioritize my long-term financial health") exert strong top-down control over lower-level, concrete policies (such as, "I will not buy this expensive item today") 10.
When faced with an uncertain scenario, medial structures like the dorsal anterior cingulate cortex (dACC) evaluate whether the current behavioral strategy is reliable 1. If the dACC determines that the current approach is failing or that the environment is too unpredictable, lateral prefrontal regions - including the frontopolar cortex - step in to generate and maintain alternative strategies 1.
Hippocampal Integration
This hierarchical planning requires intense communication between the prefrontal cortex, which acts as the brain's working memory, and the hippocampus, which stores long-term episodic knowledge 11. Together, they form a highly efficient, closed-loop neural circuit. The prefrontal cortex approximates the overarching goal, retrieves necessary complementary information from the hippocampus to evaluate past precedents, predicts the potential results of various actions, and issues control commands to correct course as the decision unfolds in real-time 11.
Catching Mistakes in Milliseconds: The Error-Related Negativity (ERN)
Because decisions under uncertainty are inherently risky, they inevitably lead to mistakes. Fortunately, the human brain is equipped with a highly sensitive internal alarm system designed to monitor performance and catch errors almost instantaneously. Within roughly 50 milliseconds of executing a flawed behavioral response - often before the individual is even consciously aware that a mistake has occurred - the brain generates a sharp electrical signature 1213.
The Brain's Immediate Alarm System
This specific neural signal, known as the Error-Related Negativity (ERN), is a response-locked event-related potential (ERP) that peaks over the medial frontal cortex 13141516. First identified in the early 1990s during rapid decision-making tasks, the ERN reflects a highly efficient, early error-monitoring process 1213. Its primary function is to facilitate immediate behavioral adaptation. When the ERN fires, it essentially commands the brain's executive networks to slow down the next response, increase focused attention, and reduce the likelihood of repeating the error 131617.
A related, though typically smaller, waveform called the Correct Response Negativity (CRN) occurs after correct responses. An enlarged CRN can sometimes indicate an increase in general uncertainty or a misclassification of a correct trial as an error, reflecting heightened overall performance monitoring 18.
Oscillations and Theta Waves
Electroencephalography (EEG) research shows that error monitoring is closely tied to specific frequency bands of brain activity. The ERN is heavily associated with theta oscillations in the medial frontal cortex 15. By analyzing both phase-locked neural activity (the traditional ERN) and non-phase-locked activity (total theta and delta power), researchers can gain a more comprehensive picture of how different neural populations synchronize to flag an error and update decision-making strategies 1516.
When the System Misfires: Uncertainty, Decision-Making, and Mental Health
Because the ERN indicates a person's underlying sensitivity to internal threats and poor performance, it has become a major focus in clinical neuroscience and psychopathology 1417. Individual differences in how strongly the brain reacts to its own mistakes are deeply intertwined with mental health vulnerabilities 1317.
Hyperactive Error Monitoring in Anxiety and OCD
An abnormally enlarged, or potentiated, ERN is frequently observed in individuals diagnosed with anxiety disorders and obsessive-compulsive disorder (OCD) 121317. For these individuals, the brain's decision-making architecture overvalues errors. The neural system reacts to minor mistakes or ambiguous uncertainties as if they were major, catastrophic threats 1217.
This hyper-vigilance is not just a symptom; it can be a predictor of pathology. Studies involving children and adolescents demonstrate that an elevated ERN can serve as a neural risk marker, prospectively predicting the first onset of generalized anxiety disorder later in life 12. Even after treatments like Cognitive Behavioral Therapy (CBT), if an individual's ERN remains elevated, they are highly vulnerable to future relapses of anxiety, suggesting that treating the behavioral symptoms does not always normalize the underlying neural uncertainty response 13.
Blunted Signals in Depression
Conversely, a significantly reduced ERN is strongly associated with depressive disorders 16. In addition to a blunted ERN, individuals with a history of depression often show blunted delta wave activity during error monitoring tasks 16. Rather than reflecting hyper-vigilance, this muted neural response is thought to represent a general reduction in motivation and task engagement 16. It acts as a physiological signature of the apathy and hopelessness often seen in depression; the brain simply registers less physiological concern when a decision goes wrong.
| Clinical Condition | Neural Signature during Error Monitoring | Behavioral Manifestation |
|---|---|---|
| Anxiety Disorders | Potentiated (enlarged) ERN | Hyper-vigilance, overvaluing minor errors, excessive worry over uncertainty 1213. |
| Obsessive-Compulsive Disorder | Potentiated ERN and increased Pe (Error Positivity) amplitude | Compulsive checking, intolerance of ambiguity, catastrophic response to perceived mistakes 1317. |
| Major Depressive Disorder | Blunted (reduced) ERN and diminished delta wave activity | Apathy, reduced motivation to correct behavior, disengagement from task outcomes 16. |
Standardizing Brain Signatures: The Quest for Reliable Biomarkers
Despite the clear theoretical links between decision-making neurobiology and mental health, translating these neural signatures into actionable clinical diagnostics remains a significant challenge. For a biological measure to be used in a hospital or clinic to guide patient care, it must meet stringent psychometric standards 12.
Within-Person vs. Between-Person Variability
A major hurdle is that clinical neuroscience frequently adopts measurement tasks from basic science. Basic experimental protocols are explicitly designed to minimize between-person variance, maximizing precision for detecting within-person mechanisms (like comparing a person's error trials to their own correct trials) 12. However, to use a tool like the ERN as a biomarker for anxiety, researchers must rely on the exact opposite: robust between-person variance that allows a clinician to distinguish one patient's brain activity from another's 12.
The Path to Clinical Utility
Currently, ERN scores often demonstrate only modest internal consistency and test-retest reliability 12. In clinical biomarker development, a reliability threshold of at least 0.90 is recommended to minimize the risk of misdiagnosis when making high-stakes decisions 12. To bridge this gap, neuroscientists are currently utilizing "location-scale multilevel models" to mathematically separate genuine trait differences in error processing from background measurement noise 12. Until data-processing pipelines are fully standardized to achieve these high reliability thresholds, tools like the ERN will remain primarily in research laboratories rather than clinical psychiatric practice 12.
The Global Brain: Overturning the Localization Myth
For decades, the standard approach in neuroscience was to study decision-making by looking at isolated brain regions, mapping specific functions to highly localized clusters of tissue 19. However, cutting-edge research is radically overturning this modular perspective.
A massive international collaboration known as the International Brain Laboratory (IBL) recently conducted an unprecedented study mapping brain activity in mice performing decision-making tasks under uncertainty 19. To ensure rigorous data reproducibility, 21 different laboratories pooled their resources and standardized their tools, recording from over 650,000 individual neurons across 279 brain areas simultaneously - representing roughly 95% of the mouse brain's volume 19.
A Simultaneous, Brain-Wide Symphony
The IBL study revealed that decision-making signals do not travel down a neat, linear, hierarchical pathway. Instead, when a decision is being made, the brain lights up "like a Christmas tree" 19. Signals are distributed globally. The parts of the brain historically thought to be responsible strictly for basic sensory processing or motor control were found to be intimately involved in anticipating outcomes and managing expectations, firing in tandem with the advanced prefrontal cortex 19.
Furthermore, prior expectations - what the brain thinks is likely to happen based on past encounters with uncertainty - are encoded throughout the entire brain, not just in localized cognitive hubs 19. This demonstrates that navigating uncertainty is not a localized cognitive event, but a simultaneous, brain-wide symphony of coordination.
Bottom line
When making a decision under uncertainty, the brain does not rely on a single, isolated "choice center." Instead, it operates as a vast, highly synchronized network that calculates odds through a drift-diffusion race for evidence, adjusts learning expectations using dopamine-driven prediction errors, and instantly flags behavioral mistakes via electrical signals like the ERN. While researchers have successfully mapped the specific cortical regions and algorithms that drive these choices, understanding how to reliably harness these signals as clinical biomarkers for mental health remains a complex, ongoing challenge.