Neurobiology of uncertainty and ambiguity processing
The human brain is fundamentally a prediction engine, evolved to detect patterns, anticipate environmental demands, and deploy biological resources accordingly. Under this paradigm, sensory information is not merely processed from the bottom up; it is continuously compared against internal, generative models of the world. When sensory input aligns with predictions, the organism maintains metabolic efficiency. When input deviates from predictions, the brain detects an error and must expend energy to update its models or alter its behavior. Within this framework, ambiguity - a state of profound uncertainty regarding future outcomes - represents a persistent, unresolvable prediction error.
The paradox of human psychology, wherein an individual often experiences greater physiological and psychological distress while waiting for a diagnosis than upon receiving a definitive negative diagnosis, is rooted in this computational architecture. Bad news, while emotionally painful, resolves a prediction error. It allows the brain to update its model of the world and deploy a specific action policy. Ambiguity, conversely, prevents the formulation of a reliable action policy, locking the nervous system in a chronic, metabolically expensive state of high alert.
Taxonomy of Environmental Uncertainty
Uncertainty is not a monolithic construct in cognitive neuroscience. To understand how the brain processes the unknown, it is necessary to divide uncertainty into discrete categories based on the epistemological status of the underlying probabilities 1.
Expected Uncertainty and Risk
Expected uncertainty, often formalized as "risk" in neuroeconomic literature, describes scenarios where outcomes are probabilistic, but the distribution of those probabilities is known or learnable 12. For example, the roll of a fair six-sided die represents risk; the outcome of any single roll is uncertain, but the probability of rolling a specific number is known with absolute certainty. In natural environments, risky decisions involve outcomes drawn from a stable, established probability distribution 2. Because the parameters of the environment are understood, the brain can compute an expected value for different choices, allowing for the deployment of optimal action policies to maximize reward or minimize harm. Under risk, uncertainty signals are primarily localized in the orbitofrontal cortex, rostral portions of the anterior cingulate cortex, and the striatum 3.
Unexpected Uncertainty and Ambiguity
Unexpected uncertainty, or "ambiguity," arises when both the outcomes and the probabilities of those outcomes are entirely unknown 12. The environment is volatile, and the probability distribution governing events is either hidden, actively shifting, or completely unlearnable. During a state of ambiguity, the brain cannot compute an expected value. Because theoretical limits on learning cannot be established, the brain is forced to continuously sample the environment in a futile attempt to resolve a probability matrix that does not exist or is unavailable 2.
Behaviorally, human beings demonstrate a profound aversion to ambiguity, frequently choosing a guaranteed negative outcome or a highly risky outcome over an ambiguous one 45. This phenomenon, termed ambiguity aversion, indicates that the absence of information is processed differently - and often more negatively - than the presence of adverse information.
| Dimension | Expected Uncertainty (Risk) | Unexpected Uncertainty (Ambiguity) |
|---|---|---|
| Epistemological Status | Probabilities of outcomes are known or discoverable. | Probabilities of outcomes are unknown, volatile, or hidden. |
| Computational State | Solvable probability distribution. | Unsolvable or non-existent probability distribution. |
| Behavioral Goal | Maximize expected reward based on known odds. | Information seeking; model building; immediate uncertainty reduction. |
| Primary Neural Correlates | Orbitofrontal cortex, rostral anterior cingulate cortex, striatum. | Anterior insula, dorsolateral prefrontal cortex, dorsal anterior cingulate cortex. |
| Psychological Experience | Calculated anticipation. | Aversive disorientation, anxiety, hesitation. |
The Free Energy Principle and Predictive Coding
To understand why ambiguity is physiologically aversive, one must view the brain through the lens of predictive coding and the free energy principle. Originating from Helmholtz's concept of unconscious inference, predictive coding postulates that the brain does not passively receive sensory input. Instead, it actively generates top-down probabilistic predictions about the causes of sensory signals 67.
Hierarchical Generative Models
According to the free energy principle, formalized by Karl Friston, all self-organizing biological systems are driven by a single imperative: to resist entropy by minimizing "surprise" or variational free energy 68. The brain maintains a hierarchical generative model of the world 910. Higher cortical levels represent abstract, long-term conceptual models, which generate predictions that cascade down to lower sensory areas via deep pyramidal neurons 6.
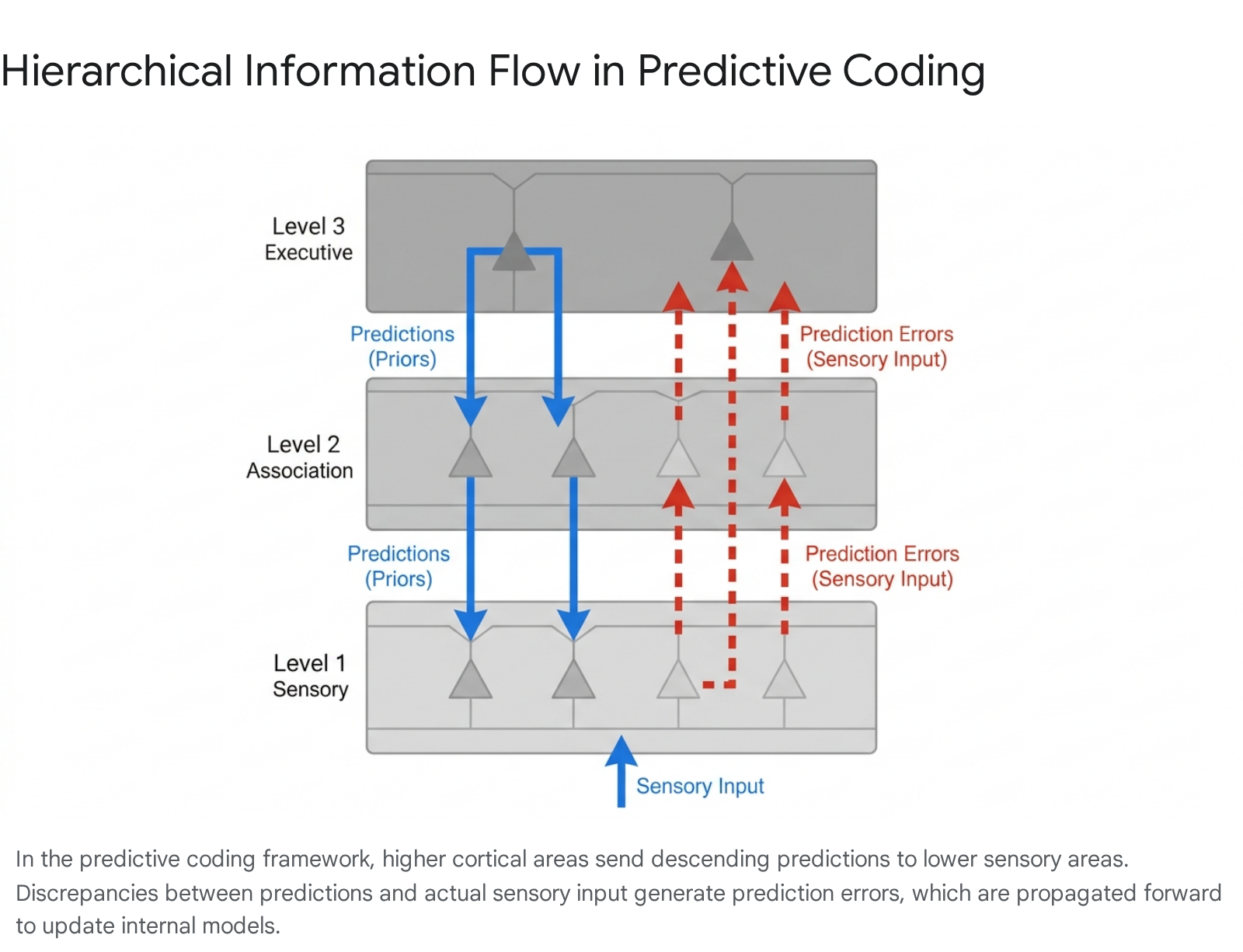
Sensory organs collect data from the environment and the body. If the sensory data matches the top-down prediction, the data is effectively "explained away," and no further cognitive processing is required 1112. The brain remains in a state of low free energy, operating with high metabolic efficiency.
Prediction Errors and Precision Weighting
When sensory data violates the brain's internal model, a prediction error occurs. These errors are encoded by superficial pyramidal neurons and ascend the cortical hierarchy, forcing the brain to either update its generative model (perceptual inference) to accommodate the new reality, or engage in actions that change the sensory input to align with the existing model 6913.
Crucially, the brain must determine which prediction errors are meaningful and which are merely environmental noise. It does this via "precision weighting," a neuromodulatory gain control process that assigns confidence to specific signals, often mediated by NMDA receptors and dopaminergic pathways 6111314. In a state of ambiguity, precision weighting fails. The brain cannot determine if the incoming sensory data is reliable, nor can it trust its own internal priors. This results in an inability to resolve the prediction error, leading to a relentless, unquenchable cycle of cognitive updating and elevated variational free energy.
Trauma and Predictive Rigidity
In populations with trauma histories or severe anxiety, predictive coding goes awry. Rather than maintaining flexible priors that update based on sensory evidence, traumatic experiences can establish highly rigid, over-weighted priors - "hyper-priors" - that interpret neutral or ambiguous stimuli as dangerous 121516. According to recent neurocomputational models, Post-Traumatic Stress Disorder (PTSD) is fundamentally a disorder of prediction rather than a disorder of memory storage 15. The brain assigns excessive precision to threat predictions, causing it to anticipate danger everywhere. Consequently, the brain enters a state of chronic hypervigilance, desperately attempting to minimize prediction errors by avoiding ambiguous stimuli altogether 1516.
Active Inference and Allostatic Regulation
The brain minimizes free energy through a process known as active inference 68917. Active inference posits that the brain does not merely passively observe the world to update its models; it acts upon the world to generate sensory data that confirms its prior predictions 6912.
Action Policies and Expected Surprise
Cognitive agents commit to action policies based on an estimation of how effectively those actions will minimize expected surprise 9. Under conditions of expected uncertainty (risk), the brain can map out an action policy because the probability distribution is known.
Under conditions of unexpected uncertainty (ambiguity), the brain cannot map an action policy because the outcome states are entirely unpredictable. The inability to formulate an action policy means the organism cannot exert control over its environment 9. When active inference is paralyzed, the brain cannot guide behavior to minimize free energy, leading to intense psychological frustration.
The Metabolic Tax of Prediction Error
The distress associated with ambiguity is quantifiable as a drain on the body's energy reserves. This is best understood by viewing predictive coding through the physiological framework of allostasis. Homeostasis is the reactive maintenance of physiological set points, whereas allostasis is the predictive regulation of the body's internal environment in anticipation of future demands 171819.
The brain's generative models are intrinsically tied to the body's energy budget. The human brain accounts for roughly 20 percent of the body's total metabolic consumption; therefore, it cannot afford to process every piece of sensory data from scratch 1920. Predictive coding is an evolutionary mechanism designed to conserve energy - it is metabolically cheaper to predict sensory input than to process it continuously 19.
When a prediction error occurs, the brain cannot rely on its energy-efficient, automated models. It is forced to recruit expensive, higher-order neural resources to process the novelty and update the model 20. Unresolved prediction error - such as waiting for a medical diagnosis or corporate restructuring - imposes a severe "metabolic tax" on the organism 2021.
Efficiency Topologies and Systemic Exhaustion
Because the brain cannot resolve the uncertainty cognitively, it triggers systemic allostatic adjustments to prepare the body for any potential outcome 1819. The brain essentially concludes that if the environment is unpredictable, a threat may be imminent, and thus mobilizes physical energy reserves to ensure survival 1821. This explains why the physiological state of waiting for a diagnosis feels physically exhausting: the brain is forcing the body to burn metabolic fuel to manage an unrelenting stream of prediction errors.
When bad news arrives, the ambiguity collapses. The probability distribution shifts from unknown to certain. While the news triggers acute emotional stress, the computational prediction error is resolved 920. The brain updates its internal model and immediately calculates new action policies. The chronic metabolic hemorrhage caused by the prediction error ceases, which accounts for the paradoxical sense of relief individuals report when long-feared bad news is confirmed.
Neuroanatomical Correlates of Ambiguity
The differentiation between risk, ambiguity, and threat processing is supported by highly specific functional topographies within the human brain. Recent Activation Likelihood Estimation (ALE) meta-analyses encompassing 76 fMRI studies confirm a distributed neural network devoted to uncertainty processing 222324.
The Insular Cortex and Interoception
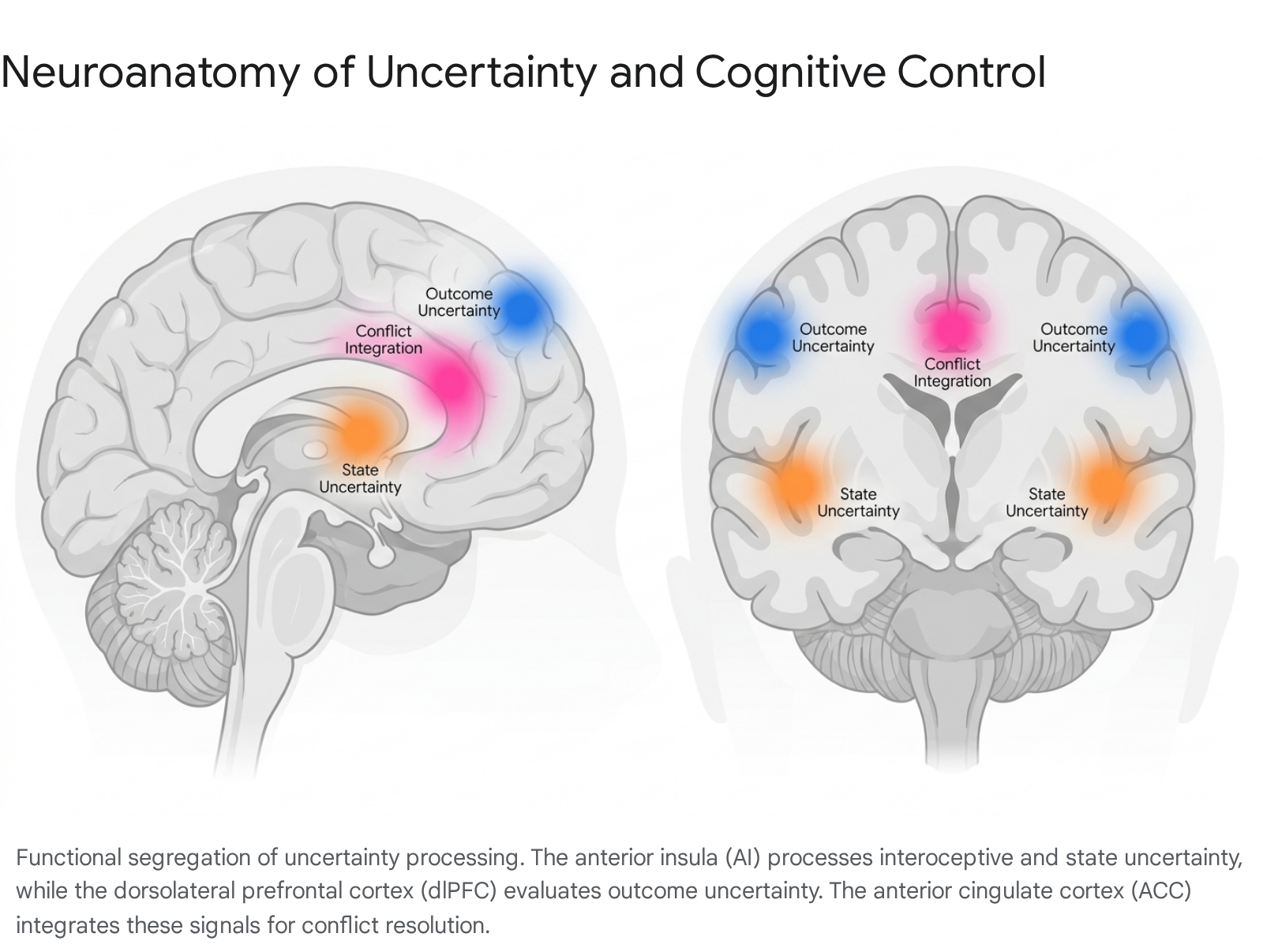
The insular cortex is one of the most frequently activated regions in functional neuroimaging studies involving uncertainty 12225. The insula serves as an integrative hub that bridges internal physiological states (interoception) with external environmental stimuli 25. Within the predictive coding framework, the posterior insula receives initial interoceptive inputs, which are then relayed to the anterior insula (AI) and integrated with motivational and cognitive signals 25.
Under conditions of ambiguity, the AI generates a prediction signal indicating the possibility of future aversive interoceptive states - broadcasting a warning that the body's homeostasis may soon be disrupted 2627. Meta-analyses reveal distinct hemispheric asymmetries: the left anterior insula is strongly associated with reward evaluation under uncertainty, while the right anterior insula is heavily involved in cognitive control, learning, and error detection 222324. In top-down processes, the AI connects with the dorsolateral prefrontal cortex (dlPFC) and superior marginal gyrus to represent cognitive control of emotion, whereas bottom-up interoception involves AI connectivity to the nucleus tractus solitarius (NTS) 27.
Prefrontal and Cingulate Integration
Operating in tandem with the insula is the anterior cingulate cortex (ACC), a major node in the salience network. The ACC tracks the volatility of the environment, estimates error likelihood, and signals decision conflict 15. When the environment is ambiguous, the ACC identifies the conflict between multiple competing action policies and signals the need for increased cognitive control 122.
The prefrontal cortex, specifically the dlPFC and the ventromedial prefrontal cortex (vmPFC), handles the calculation of specific outcome uncertainties 1522. While the anterior insula responds to the general context of state ambiguity, the dlPFC attempts to process distinct outcomes 1.
Interestingly, functional brain mapping demonstrates that the states of absolute belief and disbelief activate the caudate within the basal ganglia, providing a route to influence motor action 28. In contrast, pure uncertainty lacks this basal ganglia activation, resulting in the physiological inability to translate thought into definitive action 28.
Amygdala Threat Circuitry and Top-Down Failure
The amygdala serves as the brain's primary threat detector. In predictable environments, the prefrontal cortex exerts top-down inhibitory control over the amygdala, regulating fear and anxiety 4. However, ambiguity suppresses the prefrontal cortex's executive functioning capabilities. Without precise action policies provided by the PFC, top-down inhibition is diminished 4. The amygdala interprets the prolonged ambiguity as an ongoing, unresolved threat, keeping the organism's survival circuitry continuously activated 42930.
Physiological Cascades of Anticipatory Stress
The brain's inability to resolve the prediction errors of ambiguity results in severe downstream systemic effects, mediated by the autonomic nervous system and the endocrine system.
The HPA Axis and Systemic Cortisol Saturation
When the amygdala detects the unresolved threat of ambiguity, it signals the hypothalamus. The hypothalamus releases corticotropin-releasing hormone (CRH), which prompts the pituitary gland to release adrenocorticotropic hormone (ACTH) 3031. ACTH travels via the bloodstream to the adrenal glands, stimulating the release of cortisol and adrenaline 303132.
This cascade - the Hypothalamic-Pituitary-Adrenal (HPA) axis - is designed for acute fight-or-flight responses 3031. It triggers gluconeogenesis to flood the muscles with glucose, inhibits insulin secretion, elevates heart rate, and shifts fluids from blood vessels to tissues, causing hemoconcentration 31. While highly adaptive for fleeing a predator, this cascade is entirely maladaptive when an individual is sitting in a clinical waiting room anticipating a medical diagnosis - a phenomenon that induces "White Coat Syndrome" and can falsely elevate cholesterol, LDL, and blood glucose readings 31.
Because ambiguity prevents the brain from signaling that the threat has passed, the parasympathetic nervous system is not engaged 3032. The HPA axis remains active, resulting in a chronic saturation of cortisol. Prolonged cortisol exposure disrupts reproductive and thyroid feedback loops, suppressing Luteinizing Hormone (LH), Follicle-Stimulating Hormone (FSH), and Thyroid Stimulating Hormone (TSH) 31.
Ocular and Autonomic Dysregulation
The persistent release of adrenaline in ambiguous states also causes chronic pupil dilation to increase visual light intake for threat detection 33. Constant dilation and autonomic imbalance lead to continuous eye strain, photophobia, and elevated intraocular pressure. Prolonged mental stress has been robustly linked to the exacerbation of optic neuropathy and glaucoma, creating a vicious cycle where vision loss induces further psychological uncertainty 33.
Heart Rate Variability as a Predictive Marker
Heart rate variability (HRV) - the variation in time between successive heartbeats - is a primary metric of autonomic nervous system balance and emotion regulation capacity 343536. High HRV indicates healthy parasympathetic vagal tone; low HRV indicates sympathetic dominance 3637.
Research indicates that it is not necessarily the stressful event itself that determines the cortisol response, but the physiological state during the anticipation of the event 3537. Studies tracking participants waiting for a stressful task show that a larger decrease in HRV during the ambiguous anticipation phase strongly predicts a higher cortisol spike when the stressor ultimately occurs 353738. Conversely, changes in HRV during the actual stress task itself are often uncorrelated with cortisol recovery 3537. This suggests that the physiological cost of a traumatic event is heavily dictated by how poorly the nervous system tolerates the ambiguity preceding it.
| Physiological Metric | Response to Acute Ambiguity | Consequence of Prolonged Ambiguity (Chronic Stress) |
|---|---|---|
| Cortisol | Rapid release via HPA axis activation. | Baseline elevation; immune suppression; TSH/FSH disruption. |
| Blood Glucose | Temporary spike via gluconeogenesis. | Potential hyperinsulinemia; metabolic syndrome risks. |
| Heart Rate Variability | Sharp anticipatory decrease. | Chronically low HRV; compromised emotional regulation. |
| Pupillary Response | Dilation for enhanced threat detection. | Photophobia; intraocular pressure; glaucoma risk. |
| Blood Volume | Fluid shifts to tissues (hemoconcentration). | Falsely elevated lipid and cholesterol laboratory readings. |
Electrophysiological Signatures of Uncertainty
The computational struggle to resolve prediction errors can be measured in real-time using electroencephalography (EEG), specifically through event-related potentials (ERPs) that act as biomarkers for cognitive processing 1439.
Mismatch Negativity (MMN) and Early Detection
The mismatch negativity (MMN) is a negative deflection in the fronto-central EEG amplitude occurring roughly 100 to 250 milliseconds after an unexpected stimulus is presented 1014. In the predictive coding framework, the MMN is universally recognized as the primary neural marker of a precision-weighted prediction error 1014. When the brain successfully predicts a sequence of events, MMN amplitude is low. When an ambiguous or unexpected event violates the brain's top-down generative model, the MMN spikes, reflecting the sudden mismatch between top-down expectations and bottom-up sensory realities 1014.
The P300 Component and Cognitive Integration
Following the MMN, the P300 component - a positive deflection occurring around 300 milliseconds post-stimulus - reflects the higher-order cognitive processing of the prediction error 144041. While the MMN represents the initial preattentive detection of the discrepancy, the P300 represents the conscious allocation of attention and the brain's attempt to update its working memory to accommodate the ambiguous information 1442.
Diagnostic Utility in Neuropsychiatric Disorders
In psychiatric populations heavily burdened by uncertainty and cognitive decline, such as individuals with schizophrenia, major depression, or Alzheimer's disease, the latency of the P300 is prolonged, and its amplitude is often reduced or topographically shifted 144142. Source localization techniques (e.g., sLORETA) reveal that these electrophysiological blunting effects occur in identical Brodmann Areas across both Mild Cognitive Impairment (MCI) and Alzheimer's disease, highlighting how the inability to effectively process ambiguous information serves as an early biomarker of cognitive overload and neurodegeneration 41. Further analysis of EEG time-series data using Lempel-Ziv complexity and information theory entropy confirms that spontaneous neural stochasticity is highly sensitive to the presence of chronic psychological stress and depression 42.
Intolerance of Uncertainty as a Vulnerability Factor
The degree to which the brain views prediction errors as aversive varies significantly across individuals. This variance is formalized clinically as Intolerance of Uncertainty (IU), defined as a dispositional incapacity to endure the aversive response triggered by the perceived absence of salient information 4344.
Prospective versus Inhibitory Sub-dimensions
IU is a multidimensional construct, generally divided into two sub-factors: 1. Prospective IU: The desire for predictability and the tendency to engage in anticipatory worry to prevent unexpected outcomes 444546. 2. Inhibitory IU: The experience of behavioral paralysis, hesitation, and cognitive freezing when confronted with ambiguity 444546.
Transdiagnostic Mechanisms and Psychopathology
IU is universally recognized as a transdiagnostic vulnerability factor, meaning it underlies the pathology of numerous psychiatric conditions 434748. High levels of IU are heavily implicated in Generalized Anxiety Disorder (GAD), Obsessive-Compulsive Disorder (OCD), depression, and Post-Traumatic Stress Disorder (PTSD) 464950.
Individuals with high IU experience an exaggerated allostatic stress response under ambiguous conditions. Because they find it exceptionally difficult to incorporate updated security signals into their generative models, an ambiguous context is automatically catastrophized into a threat 1243. A meta-analysis of 161 effect sizes found a strong positive relationship between high IU and maladaptive emotion regulation strategies, most notably cognitive avoidance 47. Furthermore, high IU drives behaviors like doomscrolling and rumination, which act as joint mediators linking uncertainty to severe psychological distress 46. Meta-analyses of clinical trials indicate that evidence-based therapies (like CBT) exhibit a large therapeutic effect (g = 0.89) on reducing IU, which corresponds directly with improved overall anxiety symptom severity 48.
Cross-Cultural Mediations of Uncertainty Processing
While the neurobiological architecture of uncertainty aversion is biologically innate, the cognitive interpretation and tolerance of ambiguity are heavily mediated by cultural context 455152. The Affective Neuroscience Personality Theory (ANPT) posits that universal subcortical affective systems are regulated differently across cultures via parenting styles and emotional socialization 52.
Recent cross-cultural neuroscientific studies reveal that societal norms regarding collectivism, individualism, and cultural tightness influence how IU manifests 4550. For instance, a latent profile analysis using Schwartz's theory of basic human values compared university students in the United States and South Korea 455354. In both populations, high inhibitory IU (paralysis in the face of the unknown) was a strong predictor of membership in tradition-oriented or security-focused value profiles, leading to lower intercultural sensitivity 4554. The brain's attempt to minimize free energy in a complex social environment leads individuals with high IU to retreat into tightly controlled, homogenous social networks where prediction errors are minimized.
Furthermore, studies across the United States, Mexico, and China reveal significant variances in how IU impacts overall life satisfaction 55. In the US and Mexico, elevated IU is strongly and inversely associated with life satisfaction. However, in Chinese samples, this direct inverse relationship with life satisfaction is not observed to the same degree, suggesting that highly collectivist or tight cultures may offer buffering mechanisms against the individual distress of ambiguity 55.
| Cultural Context | Psychological Mediators | Impact of High Intolerance of Uncertainty (IU) |
|---|---|---|
| United States & Mexico | Highly individualistic; looser cultural norms. | Strong inverse relationship with life satisfaction; high reported PTSD symptoms during global crises. |
| China | Collectivist; high societal structure. | Association with depressive symptoms, but no significant inverse impact on overall life satisfaction. |
| South Korea | High cultural tightness; strong tradition values. | High IU correlates with lower intercultural sensitivity and preference for security-focused value profiles. |
| Lebanon | High-stress environmental context. | "Fear of Happiness" serves as a major mediator between IU and severe anxiety symptoms. |
Active Intersubjective Inference and Therapeutic Models
Building upon predictive coding, emerging psychodynamic models propose Active Intersubjective Inference (AISI) to explain how ambiguity affects social identity. AISI posits that the self emerges from recursive inferences about how others perceive us - the "second-order self" 1656.
In psychopathology, maladaptive core beliefs (e.g., "I am not enough") function as high-precision hyper-priors. When a person with high IU receives ambiguous social feedback, their hyper-prior dictates that the ambiguity is evidence of rejection 1256. New therapeutic models, such as the Resonance-Inference Model (RIM) and Active Inference Therapy (AIT), seek to leverage Bayesian updating to help patients relax these rigid priors 1257. By introducing tolerable amounts of uncertainty in a safe clinical environment (such as via EMDR or psychedelics), therapies aim to induce entropy-mediated plasticity, allowing the brain to learn how to process prediction errors without triggering massive allostatic stress cascades 141657.
The Appetitive Transformation of Uncertainty
While the human brain generally treats ambiguity as an aversive state requiring resolution, the nervous system also possesses an intrinsic mechanism to reframe uncertainty as an appetitive, rewarding experience: curiosity 585960.
Information Gap Theory and Curiosity
From a computational perspective, curiosity can be defined via "information gap theory" - the recognition of a discrepancy between what is known and what is desired to be known 5859. While this gap generates an information prediction error, the valence of this error can be flipped.
Neurobiologically, primary rewards (like food or money) and the acquisition of information share strikingly similar behavioral and anatomical properties 58. The resolution of uncertainty is intrinsically rewarding 59. When an individual successfully leverages curiosity to seek out missing information, the brain's reward circuitry is activated, specifically involving dopamine release in the striatum and midbrain 1460.
Dopaminergic Reward Circuitry and Exploration
When the brain encounters ambiguity, it typically activates the amygdala-driven threat circuitry 4. However, when an individual adopts a curious mindset - shifting internal framing from "What if something terrible happens?" to "I wonder what will happen?" - the brain shifts out of the survival-oriented fight-or-flight mode and engages the SEEKING system 460.
This reframing allows the brain to interpret the prediction error not as a threat to survival, but as an opportunity for learning 60. Curiosity primes the hippocampus, enhancing memory formation and learning efficacy 5960. Paradoxically, studies demonstrate that while the state of outcome uncertainty initially decreases subjective happiness (due to the aversive nature of the unknown), it simultaneously drives a massive increase in curiosity 58. This suggests that curiosity operates as an evolutionary, appetitive drive specifically designed to motivate organisms to reduce the unpleasant, metabolically taxing state of uncertainty by actively seeking information 5859.
Conclusion
The human brain is a predictive organ engaged in a perpetual battle against entropy. It seeks to model the world to conserve metabolic energy and deploy adaptive behaviors. In this context, certainty - even certainty of an adverse outcome - allows the brain to finalize its algorithms, accept the state of the world, and act.
Ambiguity interrupts this fundamental biological mandate. It traps the cognitive control networks - spanning the anterior insula, anterior cingulate cortex, and prefrontal cortex - in a continuous loop of unresolved prediction errors. This computational failure prevents the prefrontal cortex from down-regulating the amygdala, resulting in the chronic activation of the HPA axis. The ensuing flood of cortisol, the withdrawal of parasympathetic vagal tone, and the exhausting metabolic tax required to sustain an unresolvable cognitive state explain why waiting in the dark is profoundly more stressful than standing in the light. Understanding these mechanisms demystifies human anxiety and points to curiosity, cognitive flexibility, and active inference as the primary biological antidotes to the inherent unpredictability of the natural world.