How Chronic Poor Sleep Affects the Brain Week by Week
Chronic poor sleep systematically degrades the brain's architecture, shifting it from a state of nightly repair to sustained neurological stress. Week by week, this lack of rest severs emotional regulatory circuits, halts the birth of new neurons, triggers severe neuroinflammation, and ultimately leads to irreversible cell death and structural brain atrophy.
When we think of sleep deprivation, we often visualize the acute effects of pulling an all-nighter: the heavy eyelids, the reliance on caffeine, and the undeniable brain fog that follows a full twenty-four hours awake. However, the modern epidemic of sleep loss rarely involves total sleep deprivation. Instead, it is characterized by chronic sleep restriction - sleeping five or six hours a night, week after week, month after month. While the human brain is remarkably resilient to a single night of bad sleep, chronic sleep loss triggers a cascade of compounding structural and chemical changes that fundamentally rewire our neurology 123.
Through a combination of advanced neuroimaging, behavioral studies, and molecular biology, researchers have mapped exactly what happens to the human brain as sleep debt accumulates. The timeline of this neurological degradation moves from initial chemical imbalances and impaired waste clearance in the first few days, to profound structural atrophy and permanent cell loss observed after months and years of restricted rest 456. This report details the week-by-week deterioration of the brain under chronic sleep restriction, exposing the biological mechanisms that drive cognitive decline, emotional volatility, and accelerated brain aging.
Days 1 to 3: Chemical Imbalances and Waste Accumulation
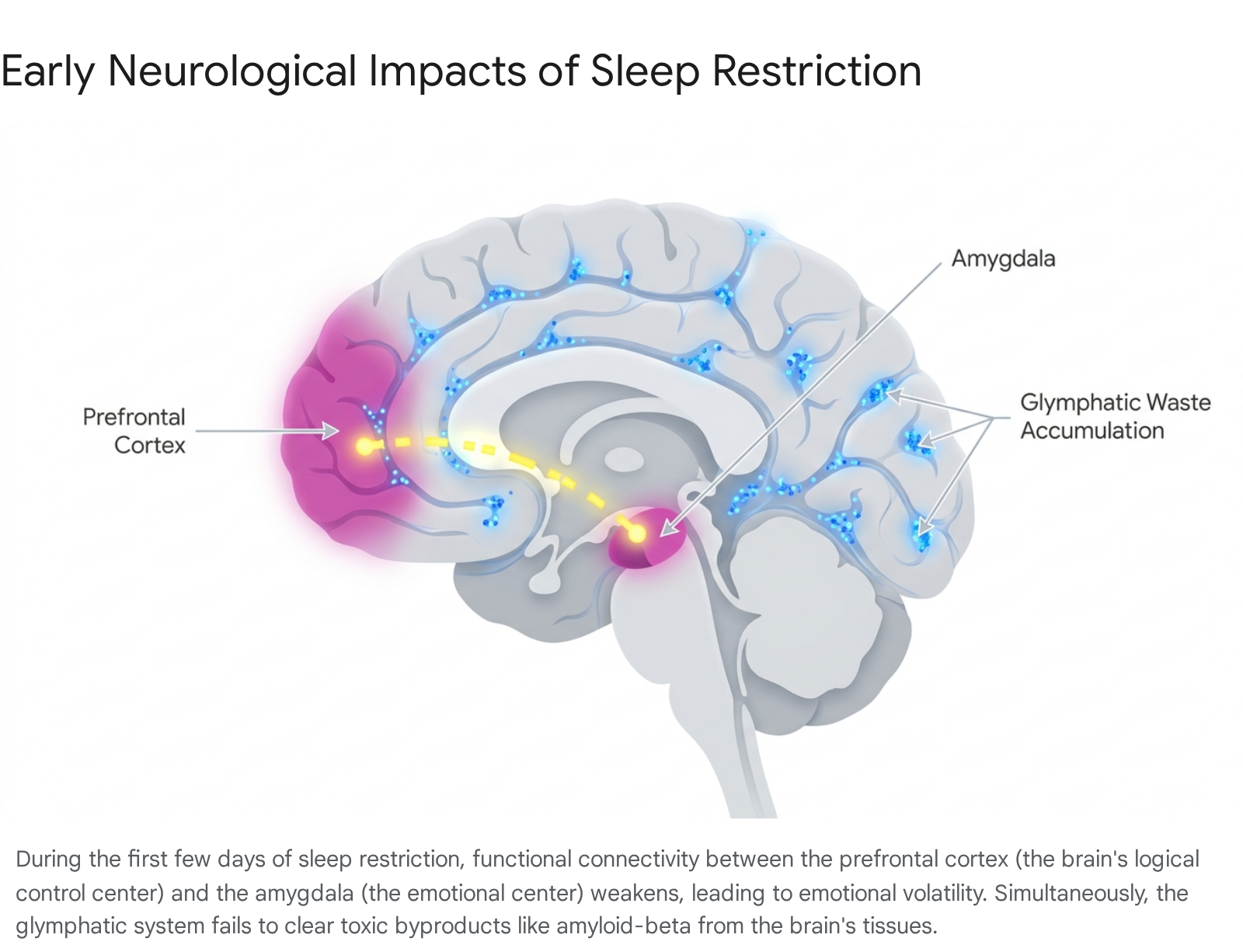
The brain is a highly metabolically active organ. During waking hours, it consumes vast amounts of energy to process sensory information, execute physical movements, and maintain consciousness. This relentless activity produces a significant amount of metabolic byproducts. The first few days of sleep restriction are defined by the brain's sudden inability to effectively clear this waste, alongside a rapid fracturing of its chemical communication networks.
The Failure of the Glymphatic System
For decades, scientists did not fully understand how the brain, which lacks a traditional lymphatic system, cleared away its cellular trash. It is now known that the brain possesses a unique, fluid-based waste disposal mechanism known as the glymphatic system 478. This system is driven by aquaporin-4 (AQP4) water channels located on the endfeet of specialized support cells called astrocytes 8. These channels facilitate the convective exchange between cerebrospinal fluid and the brain's interstitial fluid, effectively pressure-washing the brain's tissues 78.
Crucially, the glymphatic system is primarily active during deep, slow-wave sleep. When we sleep, the brain's parenchymal resistance drops - meaning the cellular structure of the brain physically relaxes and expands, allowing fluid to flow more freely 4. A landmark 2026 clinical trial published in Nature Communications directly demonstrated this mechanism in humans. By utilizing continuous overnight monitoring and advanced plasma biomarker analysis in a randomized crossover trial of 39 participants, researchers showed that a normal night of sleep significantly increases the morning plasma levels of Alzheimer's-linked proteins, such as amyloid-beta (Aβ40 and Aβ42) and tau (both phosphorylated and non-phosphorylated) 479. These elevated blood levels are not a sign of disease; rather, they are definitive evidence that the glymphatic system successfully flushed these toxic, aggregation-prone proteins out of the brain tissue and into the bloodstream for disposal 479.
When sleep is restricted to just a few hours a night, the brain experiences a sharp reduction in non-rapid eye movement (NREM) EEG delta power, which is the electrical signature of deep, restorative sleep 4. Without this deep sleep, brain parenchymal resistance remains high, and cerebrovascular compliance drops 49. The glymphatic system's plumbing effectively slows to a trickle. By the second or third day of poor sleep, amyloid-beta, tau, lactate, and other metabolic byproducts begin to accumulate in the brain's interstitial spaces, setting the physiological stage for long-term neuroinflammation and cognitive decline 8910.
Emotional Dysregulation: The Amygdala-PFC Disconnect
As physical waste builds up in the background, the brain's emotional control centers are the first behavioral circuits to fracture. In a well-rested brain, the prefrontal cortex (PFC) - the highly evolved region responsible for executive function, logic, and behavioral control - exerts strong top-down regulatory control over the amygdala, the brain's primitive fear and emotion center 1110. This neural pathway ensures that we react to stressors with measured, logical responses rather than raw emotional outbursts.
Resting-state functional magnetic resonance imaging (fMRI) studies reveal that even acute, short-term sleep loss severely disrupts this pathway. Functional connectivity between the emotion-regulating regions of the ventromedial prefrontal cortex (vmPFC) and the right amygdala is significantly reduced after sleep deprivation 1011. Specific sub-regions, such as the basolateral amygdala (BLA), show decreased connectivity with executive control areas like the dorsolateral prefrontal cortex (DLPFC) and the dorsal anterior cingulate cortex (ACC) 11.
Without the prefrontal cortex applying its inhibitory brakes, the amygdala becomes hyperactive and begins to couple with other regions, such as the posterior cingulate cortex and parahippocampal gyrus, which are involved in generating internal emotional states 11. This physical disconnect explains why, after just a few days of poor sleep, individuals become highly reactive to negative emotional stimuli, experience increased subjective psychological distress, and suffer measurable declines in emotional intelligence 1112. The brain essentially reverts to a more primitive, reactive state, prioritizing immediate emotional survival responses over reasoned appraisal 1110.
The Breakdown of Social Memory Circuits
Cognitive deficits also begin to manifest within the first few days, particularly in highly specialized memory circuits. One such circuit resides in the CA2 region of the hippocampus. The hippocampus is universally recognized for its role in memory and learning, but recent discoveries have pinpointed the CA2 area as a critical hub specifically linking the sleep cycle to the formation of social memories - the ability to recognize, differentiate, and remember familiar individuals 151314.
Just five hours of sleep deprivation is enough to disrupt the maintenance of synaptic plasticity in this specific CA2 region 151418. Electrophysiological recordings show that sleep loss markedly impairs long-term potentiation (LTP), the primary cellular mechanism behind learning, leading to weakened communication between neurons 151419. This biological failure results in noticeable deficits in social recognition; the brain literally loses its capacity to form new social memories or distinguish new acquaintances from familiar ones 141819.
Research published in Neuropsychopharmacology in 2026 revealed that this specific memory deficit is mediated by the upregulation of adenosine A1 receptors and a signaling molecule called PDE4A5, which accumulate during wakefulness and actively dampen brain activity 151319. Remarkably, administering caffeine - a known adenosine receptor antagonist - specifically targets the CA2 region to stabilize synaptic plasticity and restore these broken neural connections, effectively reversing the social memory deficits caused by short-term sleep loss without overstimulating the rest of the brain 151314.
Days 4 to 7: The Illusion of Adaptation and Network Reorganization
As poor sleep persists across a full week, the brain enters a dangerous transitional phase. The initial shock of exhaustion begins to fade, replaced by a psychological phenomenon where the brain loses its ability to accurately judge its own level of impairment. This is often referred to in sleep science as the "illusion of adaptation."
The Divergence of Sleepiness and Performance
One of the most insidious aspects of chronic sleep restriction is that it tricks individuals into believing they are functioning normally. This phenomenon was definitively mapped in a seminal 2003 dose-response study by Hans Van Dongen, David Dinges, and colleagues 151617. In this highly controlled laboratory experiment, researchers restricted the sleep of healthy adults to either 4, 6, or 8 hours a night for 14 consecutive days 1617. They measured both the participants' subjective sleepiness (how tired they felt, using standardized rating scales) and their objective cognitive performance using a Psychomotor Vigilance Task (PVT), which measures sustained attention, reaction times, and lapses in behavioral alertness 161723.
The results revealed a startling disconnect between perception and biological reality. Subjective sleepiness ratings showed an acute spike in the first two to three days of sleep restriction, but then they largely plateaued 161825. The participants essentially stopped feeling increasingly tired; they reported that they had adapted to their new, restricted sleep schedule and felt they were managing well 1623.
However, their actual objective cognitive performance told an entirely different story. Deficits in cognitive throughput, working memory, and behavioral alertness did not plateau; they accumulated linearly across every single day of the sleep restriction period 172325. The brain's processing speed slowed, and the number of times participants completely failed to respond to a stimulus (a cognitive lapse) skyrocketed 1517.
Data indicates a notable and alarming trajectory: by the end of the 14 days, the participants who had been sleeping six hours a night - a schedule exceedingly common in modern working society - exhibited cognitive performance deficits that were completely indistinguishable from individuals who had been kept awake for two full days and nights (up to 48 hours of total sleep deprivation) 1617. A statistical model derived from the study revealed that these lapses in behavioral alertness were near-linearly related to the cumulative duration of wakefulness 1723.
To illustrate this divergence between how sleep-deprived individuals feel and how they actually perform, consider the following breakdown of the phenomena observed during chronic sleep restriction:
| Metric Tracked | Initial Reaction (Days 1-3) | Prolonged Reaction (Days 4-14) | Neurological Implication |
|---|---|---|---|
| Subjective Sleepiness | Sharp increase in feelings of fatigue. | Plateaus; individual feels they have "adapted." | Loss of self-awareness regarding cognitive deficits. |
| Objective Cognitive Lapses | Moderate increase in errors. | Linear, continuous accumulation of severe errors. | Deficits equivalent to 48 hours of total sleep deprivation. |
| Working Memory | Noticeable decline in recall. | Cumulative deterioration of cognitive throughput. | Inability to hold and process complex information. |
| Physiological Sleep Drive | Acute changes in sleep EEG markers. | Negligible further changes; drive remains constantly high. | The brain maintains a chronic state of unmet homeostatic need. |
The danger of this illusion is profound. Because people do not feel increasingly sleepy after the first few days, they assume their cognitive accuracy, processing speed, and judgment are intact, making them prone to critical errors in professional and safety-critical environments 1626.
Global Brain Network Reorganization
During this first week, the brain actively attempts to compensate for its exhaustion by reorganizing its functional networks. Modern neuroimaging demonstrates that the brain responds differently to a single night of total sleep deprivation (TSD) versus a week of chronic sleep restriction (CSR) 327.
Using Covariate-Constrained Manifold Learning (CCML) and graph-theoretical analyses on resting-state fMRI data, researchers have shown that after five days of chronic sleep restriction, the brain exhibits widespread and distinct changes in its intrinsic functional architecture 32719. Network integration is severely reduced, meaning different regions of the brain struggle to communicate with one another 27. Regional topological changes occur specifically within the subsystems of the default mode network (DMN) - which governs self-reflection and mind-wandering - as well as the frontoparietal network (involved in sustained attention) and the cerebellum 327.
The brain attempts to stabilize its performance by shifting the cognitive load to alternative neural networks, but it does so at a severely reduced operational capacity 26202122. This topological reorganization explains why complex decision-making and sustained vigilance become overwhelmingly difficult as the week progresses; the brain's internal communication highways are essentially experiencing rolling blackouts 1526.
Weeks Two to Three: Neuroinflammation and Suppressed Neurogenesis
As chronic sleep restriction extends into a second and third week, the damage shifts from functional connectivity disruptions to deeper cellular and inflammatory pathology. The brain transitions from being merely tired to actively inflamed.
The Neuroinflammatory Cascade
Sleep deprivation acts as an intense physiological stressor on the entire body. The lack of restorative downtime triggers the hypothalamic-pituitary-adrenal (HPA) axis and the sympathetic nervous system, resulting in the sustained, abnormal release of glucocorticoids (such as cortisol) and catecholamines 2324. Normally, cortisol levels are minimal at the beginning of the sleep period, but chronic sleep fragmentation and restriction prevent this necessary dip, keeping the body in a state of high alert 2425.
This systemic stress disrupts immune homeostasis and initiates a severe neuroinflammatory cascade. Within the central nervous system, chronic sleep loss activates microglia and astrocytes - the brain's resident immune and support cells 242627. When primed by the stress of sleep deprivation, these glial cells shift into an aggressive, phagocytic state. They release a storm of pro-inflammatory cytokines, specifically Tumor Necrosis Factor-alpha (TNF-α), Interleukin-1β (IL-1β), and Interleukin-6 (IL-6) 242627. These cytokine shifts are observed in both central and peripheral compartments, indicating a system-wide inflammatory crisis 27.
This inflammatory response is compounded by a dangerous failure in the brain's natural self-regulation. Animal studies have shown that a week of sleep deprivation leads to the significant downregulation of the α7 nicotinic acetylcholine receptor (α7-nAChR) on microglia and astrocytes 26. This receptor normally plays a crucial role in counter-regulating immune responses and activating anti-oxidant pathways (like the PI3K/AKT/GSK-3β pathway) 26. As this receptor's activity drops, the brain's primary anti-oxidant enzymes (such as Nrf-2 and HO-1) are suppressed 26. Consequently, oxidative stress skyrockets, creating a feed-forward loop of inflammation and oxidative damage 2426.
In humans, this neuroinflammation can be actively visualized using Positron Emission Tomography (PET) scans. Researchers utilize specific radiotracers, such as [11C]PK11195 and F-18 DPA-714, which bind to the 18-kDa translocator protein (TSPO) 373828. TSPO is an outer mitochondrial membrane protein that is heavily upregulated in activated microglia during inflammation 372930. Similarly, new PET imaging techniques target Cyclooxygenase-2 (COX-2), an enzyme that is highly induced in microglia and neurons during stress and pathology 293031. These advanced imaging techniques confirm that chronic physiological stressors like sleep loss induce a measurable, brain-wide inflammatory milieu that can persist for long periods, damaging synaptic connections and accelerating cognitive decline 24372943. The brain is essentially mounting an immune response against its own exhaustion.
The Halting of Hippocampal Neurogenesis
Simultaneously, the chronic stress of sleep loss begins to shut down the brain's ability to regenerate. The adult hippocampus, specifically a sub-region called the dentate gyrus (DG), is one of the few areas in the mammalian brain capable of adult neurogenesis - the birth of new neurons 3233. This ongoing cellular renewal is vital for learning, memory consolidation, and maintaining emotional resilience.
While a single night of sleep loss might temporarily increase brain-derived neurotrophic factor (BDNF) and spur a brief burst of cell proliferation in an attempt to protect the brain, chronic sleep restriction rapidly and aggressively suppresses this process 132. Animal models utilizing intermittent treadmill systems to induce sleep fragmentation - a highly accurate analog to human sleep disturbances like sleep apnea or chronic insomnia - demonstrate devastating effects on the hippocampus 253233.
After just four to seven days of fragmented sleep, the number of newly proliferating cells in the dentate gyrus (measured by markers like BrdU and Ki67) plummets by approximately 70% 2532. Furthermore, sleep deprivation fundamentally alters cell fate determination. Of the new cells that do manage to survive the initial proliferation phase, significantly fewer mature into functional, mature neurons (measured by the expression of the neuronal marker NeuN) 3233.
The sustained elevation of glucocorticoids due to the stress of sleep loss is a primary driver of this suppressed neurogenesis. When researchers experimentally prevent the elevation of these stress hormones in sleep-deprived subjects, the decrease in cell proliferation is largely eliminated, confirming that the physiological stress response to sleep loss is killing the new cells 2332. Consequently, chronic sleep restriction drastically impairs hippocampal-dependent cognitive functions. Subjects tested after weeks of sleep restriction exhibit severe spatial memory impairments and altered search strategies, as the new neurons required for these tasks were never allowed to reach functional maturity 25. Microscopic examination further reveals that the apical and basal dendritic arborization of CA1 hippocampal neurons is significantly reduced, indicating a physical shriveling of the memory circuits 2.
Months to Years: Structural Atrophy and Permanent Cell Loss
When poor sleep habits span months or years, the temporary functional connectivity changes and chemical imbalances calcify into permanent structural damage. Chronic insomnia and long-term sleep restriction are no longer viewed merely as lifestyle issues; they are recognized as powerful catalysts for accelerated brain aging and neurodegeneration.
Locus Coeruleus Neuron Degeneration and Cell Death
Perhaps the most alarming biological discovery regarding chronic sleep loss is its lethal impact on a specific cluster of cells called the locus coeruleus (LC). The LC is a tiny but immensely powerful nucleus located in the brainstem. It serves as the primary source of noradrenaline (norepinephrine) for the entire brain, projecting its neural pathways throughout the cerebral cortex 3435.
LC neurons are entirely wake-active. They fire continuously at a low tonic rate to maintain our baseline alertness while we are awake, and they fire in rapid bursts when we need to pay sudden attention to a stimulus 3435. Crucially, these neurons require sleep to survive. They decrease their firing during NREM sleep and go nearly silent during REM sleep, allowing them time to rest and clear metabolic waste 3435.
When the brain is subjected to extended, chronic periods of wakefulness, these LC neurons are forced to fire continuously without a break. This relentless activity places profound mitochondrial oxidant stress on the cells 535. Chronic sleep loss causes a dramatic reduction in essential metabolic enzymes, specifically Sirtuins 1 and 3 (SirT1 and SirT3), inside these neurons 536. Without these protective deacetylase enzymes, the neurons begin to manifest a premature senescence (aging) phenotype 5. They accumulate massive amounts of lipofuscin - a toxic aging pigment - and develop vacuolated (empty, damaged) dendrites 536.
Eventually, this intense metabolic demand leads to actual cell death. Groundbreaking animal models replicating human shift-worker sleep patterns have shown that chronic short sleep can result in the permanent loss of up to 25% to 30% of locus coeruleus neurons 537. This cell death is irreversible. Because the locus coeruleus is one of the very first regions to degenerate in the early stages of Alzheimer's and Parkinson's disease, the permanent loss of these neurons due to chronic sleep deprivation has profound, terrifying implications for long-term brain health and the acceleration of neurodegenerative disorders 353638.
The Proliferation of White Matter Hyperintensities (WMH)
Long-term sleep disturbances also leave visible, physical scars on the brain's white matter - the heavily myelinated neural cables that connect different regions of the brain and facilitate rapid communication. Large-scale longitudinal imaging analyses have tracked tens of thousands of adults to correlate their self-reported sleep duration over years with their physical brain structure 3940.
These studies rely on Magnetic Resonance Imaging (MRI) to detect White Matter Hyperintensities (WMHs), which appear as bright, abnormal spots on T2-weighted or FLAIR scans 414243. WMHs indicate areas of microstructural injury, demyelination, and cerebral small vessel disease 404144. They are widely recognized as a proxy for brain aging and are a strong predictor of future stroke, cognitive decline, and vascular dementia 414344.
Research reveals a distinct "U-shaped" relationship between sleep duration and the presence of these white matter lesions. Sleeping approximately 7.7 hours a night is consistently associated with the lowest burden of WMHs and the healthiest white matter microstructure (measured by high fractional anisotropy and low mean diffusivity) 4045. Conversely, individuals who habitually sleep less than 6 hours or more than 9 hours a night exhibit significantly increased volumes of white matter hyperintensities, particularly in the frontal, parietal, and temporal regions 40444546.
These mid-life differences in white matter integrity suggest that chronic sleep deprivation fundamentally compromises the physical infrastructure of the brain's communication networks 46. Furthermore, studies focusing on older adults on the Alzheimer's trajectory have found that sleep disturbances are positively associated with a greater accumulation of WMHs, which in turn acts as a direct mediator for future cognitive deficits 4143.
Accelerated Macroscopic Brain Atrophy
Over years, the combined, compounding effects of severe neuroinflammation, halted neurogenesis, irreversible locus coeruleus cell death, and white matter degradation result in macroscopic brain shrinkage. The brain literally begins to lose physical volume at an accelerated rate.
Longitudinal MRI tracking studies that follow cohorts of humans over five to ten years have shown that individuals who experience persistent poor sleep in midlife suffer from accelerated brain atrophy 647. The lack of restorative slow-wave sleep and REM sleep is specifically associated with reduced brain volumes in cortical regions highly vulnerable to Alzheimer's disease. These include the inferior parietal region, the posterior cingulate cortex, and the hippocampus 64849.
The visual cortex and parieto-occipital regions also show significant cortical thinning following long-term sleep deprivation 195051. Interestingly, these structural changes are highly age-dependent, with younger adults showing different vulnerabilities in the anterior insula and medial temporal lobe network compared to older adults 505253.
The total cumulative impact of this chronic sleep loss is startling. Advanced machine learning models, trained on neuroimaging data to estimate the biological "brain age" of a participant based on their degree of brain shrinkage, have quantified this damage. They found that individuals who suffer from chronic insomnia and severe difficulty sleeping possess brains that appear, structurally, up to 2.6 to 3.5 years older than the brains of well-rested peers of the exact same chronological age 47.
Summary of Neurological Degradation Over Time
| Duration of Poor Sleep | Primary Neurological Impact | Biological Mechanism of Action | Observable Cognitive/Behavioral Consequence |
|---|---|---|---|
| Days 1 to 3 | Glymphatic failure & Amygdala Disconnection | Reduced AQP4 channel fluid clearance; disrupted vmPFC-amygdala functional connectivity. | Accumulation of Aβ/tau in plasma; emotional volatility; impaired social memory (CA2 region). |
| 1 to 3 Weeks | Neuroinflammation & Suppressed Neurogenesis | Microglial activation (IL-6, TNF-α); glucocorticoid-driven suppression of dentate gyrus cell proliferation. | Subjective sleepiness plateaus while objective reaction times and working memory plummet. |
| Months to Years | Permanent Cell Death & Structural Atrophy | Mitochondrial stress in Locus Coeruleus; demyelination causing White Matter Hyperintensities (WMH). | Accelerated brain aging (by 2.6-3.5 years); permanent cognitive decline; increased dementia risk. |
The Recovery Myth: Why the Weekend Cannot Save You
A pervasive and deeply flawed habit in modern society is the concept of the "weekend catch-up" - sleeping five hours a night from Monday to Friday to meet work demands, and then attempting to sleep ten hours on Saturday and Sunday to repay the accumulated "sleep debt." Neurological research unequivocally proves that this strategy is biologically insufficient to reverse the damage 1.
The Asymmetric Timeline of True Recovery
Recovery from chronic sleep deprivation is a highly asymmetric process. The brain requires exponentially more time to heal its functional networks and clear inflammatory markers than it took to incur the damage in the first place. While research varies slightly on the exact mathematical ratio based on individual genetics and age, several robust sleep science frameworks and aviation fatigue management models suggest a sobering reality: complete recovery from chronic sleep deprivation requires roughly 4 to 9 days of consistent, quality rest for every single hour of accumulated sleep debt 2568.
When researchers rigorously test cognitive function after a period of chronic restriction, they find that different brain domains recover at vastly different rates 2569:
- Reaction Time and Basic Alertness: Basic psychomotor speed and physical energy are usually the first to rebound. These metrics can sometimes return to baseline after 2 or 3 days of adequate, extended sleep 6954.
- Sustained Vigilance, Complex Memory, and Mood: These higher-order executive functions are highly resistant to short-term recovery. In controlled studies where participants were restricted to 4 or 5 hours of sleep for a week, a single 10-hour recovery sleep failed entirely to correct deficits in vigilant attention or subjective mood 21855.
- Complete EEG Restoration: Even after a full 7 days of unrestricted recovery sleep following a 10-day period of sleep restriction, participants' underlying EEG brain activity patterns and complex cognitive metrics (like Stroop task accuracy for executive function) often fail to fully return to their pre-deprivation baseline 54.
The deeply troubling nature of incomplete recovery was highlighted in a classic 2003 study by Gregory Belenky and colleagues. Participants were restricted to 3, 5, or 7 hours of sleep for seven days, and then given three full days of 8-hour recovery sleep 202122. During the recovery phase, the cognitive performance of the 3-hour group improved slightly, but only to match the already-impaired level of the 5-hour and 7-hour groups 1820. Most alarmingly, the performance of the 5-hour and 7-hour groups never improved during the three recovery days 202122. Their brains had adapted to a restricted operational capacity and remained physically stuck at that lower, impaired level of functioning despite returning to 8 hours of sleep 20212256. This suggests that the brain alters its baseline operational capacity during chronic restriction, and un-doing that alteration takes immense time.
Is the Damage Reversible?
The reversibility of long-term structural changes remains a highly contested area of active study. While acute functional connectivity changes (like the prefrontal-amygdala disconnect) are largely reversible with sufficient rest 19, the actual death of Locus Coeruleus neurons cannot be undone; once those cells undergo apoptosis due to metabolic stress, they are lost forever 537.
However, there is hope found in the mechanisms of neuroplasticity. Recent studies in animal models demonstrate that placing sleep-deprived subjects in an "Enriched Environment" (environments offering high intellectual, social, and physical stimulation) can actually reverse chronic sleep deprivation-induced damage to cellular plasticity in the hippocampus 157. The enriched stimulation promotes neuronal repair pathways and can rescue the halted process of neurogenesis in the dentate gyrus 57.
Furthermore, targeted interventions, such as ensuring adequate time in deep Slow-Wave Sleep (which maximizes the slow-wave energy needed to flush the glymphatic system), are essential for physiological repair 1855. But ultimately, preventing permanent damage requires completely abandoning the concept of "catching up." Once sleep debt triggers the neuroinflammatory cascades, microglial activation, and structural demyelination characteristic of chronic restriction, the brain requires weeks - if not months - of consistent, high-quality, 7-to-8 hour sleep schedules to truly recalibrate and clear the physiological stress 2569.
Bottom line
Chronic sleep restriction systematically degrades the brain, moving rapidly from functional connectivity disruptions in the first few days to severe, brain-wide neuroinflammation and suppressed neurogenesis within weeks. Over months and years, this sustained biological stress leads to irreversible cell death in critical attention centers and accelerates macroscopic structural brain atrophy, effectively aging the brain by several years. While a weekend of extra rest may temporarily alleviate subjective feelings of tiredness, true cognitive and cellular recovery requires weeks of consistent, restorative sleep, underscoring the stark reality that neurological sleep debt cannot simply be "paid off" in a matter of days.