Neuroscience of glossophobia and the threat response
Social evaluative threat represents a pervasive psychological phenomenon in which an individual perceives an acute risk of being negatively judged, scrutinized, or rejected by their peers. Glossophobia, clinically recognized as the severe fear of public speaking, serves as one of the most reliable and acute manifestations of this social evaluative threat 12. From an evolutionary neuroscience perspective, human survival in ancestral environments relied entirely on group cohesion, interpersonal cooperation, and social inclusion. In such contexts, ostracism, rejection, or loss of social standing equated to a loss of resources and physical protection, effectively functioning as a death sentence 13. Consequently, the human nervous system evolved to process acute social evaluation through the exact same primary neural and endocrine survival circuits that are responsible for detecting and responding to immediate physical predators 145. The neuroscience of glossophobia is therefore not merely the study of a specific situational anxiety, but a broader inquiry into how the brain interprets interpersonal scrutiny as an existential threat to biological survival.
Neuroanatomical Foundations of Public Speaking Anxiety
The neurobiology of public speaking anxiety is characterized by a profound network imbalance: the hyperactivation of subcortical threat-detection structures combined with diminished regulatory, top-down control from higher-order cortical networks. This specific structural and functional dynamic produces the intense physiological, affective, and cognitive symptoms experienced by individuals before and during a public speaking event.
Amygdala Reactivity and Threat Detection Pathways
The amygdala serves as the central neuroanatomical hub for threat detection, fear conditioning, and the initiation of the biological stress response. In instances of public speaking anxiety, complex visual and auditory stimuli - such as a crowd of staring faces, specific facial micro-expressions of judgment, or the silence of an audience - are processed initially by the sensory organs. These signals are routed rapidly to the thalamus, which functions as the brain's primary sensory relay station 6. From the thalamus, the threat signal bifurcates into two distinct neural pathways. The first is a direct subcortical pathway, frequently termed the "low road," which bypasses conscious cortical processing and travels directly to the amygdala 6. This pathway allows for near-instantaneous autonomic reactions before any conscious appraisal of the situation can occur, effectively prioritizing speed over accuracy in threat detection 564. The second pathway, or "high road," routes the sensory information through the prefrontal cortex for a slower, more deliberate logical assessment before projecting back to the amygdala 6.
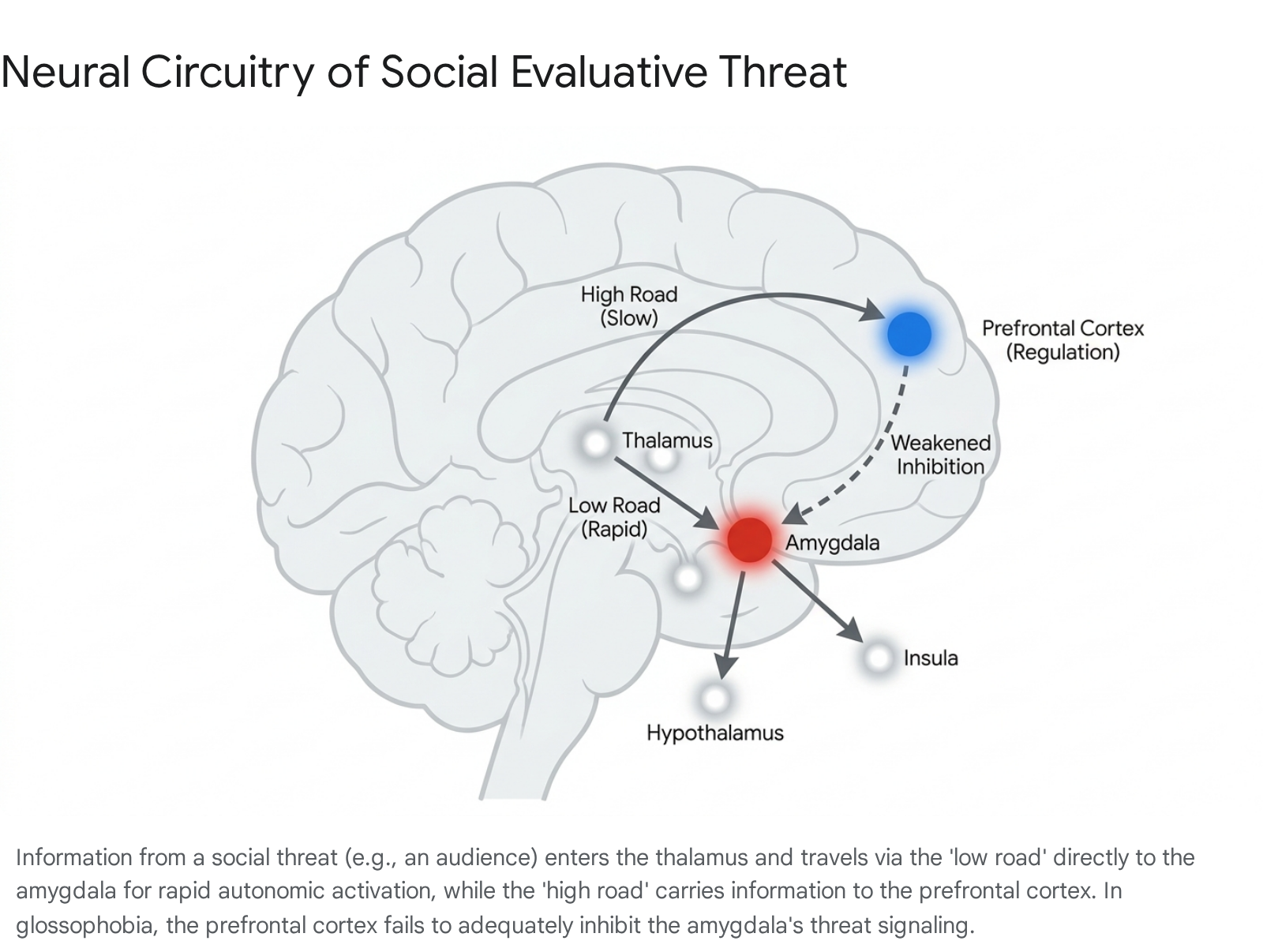
Functional magnetic resonance imaging (fMRI) studies consistently demonstrate exaggerated amygdala reactivity in individuals with social anxiety disorder (SAD) and severe glossophobia when they are exposed to disorder-related stimuli, such as harsh, rejecting, or evaluating facial expressions 567. This subcortical hyperactivity directly triggers the hypothalamus, which acts as the command center for the systemic autonomic stress cascade 54. Furthermore, resting-state and task-based connectivity analyses reveal hyperconnectivity between the amygdala and the globus pallidus during socially anxious threat processing 6. The involvement of the globus pallidus - a major component of the basal ganglia - highlights the critical relevance of motor-regulatory structures in the physical and affective expressions of fear, such as trembling or freezing behaviors often observed in highly anxious speakers 6.
Prefrontal Cortex Dysfunction and Top-Down Regulation
The prefrontal cortex (PFC), specifically the ventromedial prefrontal cortex (vmPFC) and the dorsolateral prefrontal cortex (dlPFC), is the primary cortical region responsible for executive function, emotional regulation, and cognitive reappraisal 589. In individuals without public speaking anxiety, the PFC effectively evaluates the perceived threat of an audience through the "high road" and successfully transmits inhibitory signals down to the amygdala to dampen the acute fear response 6810. This specialized primate prefrontal expansion underlies the human capacity to engage high-level regulatory strategies to cope with and modify the subjective experience of anxiety 11.
In highly anxious individuals facing public speaking, there is a documented failure of this top-down regulatory mechanism. Resting-state fMRI meta-analyses reveal pronounced hypo-connectivity between the amygdala and several regulatory cortical nodes, including the medial frontal gyrus, the anterior cingulate cortex (ACC), and the vmPFC in patients with clinical anxiety 71112. When confronted with social evaluative threat in real time, individuals with glossophobia exhibit significantly diminished recruitment of these cognitive control regions compared to healthy controls 511. Because the neural pathways that interface with distinct inhibitory systems within the cortex and thalamus fail to regulate the amygdala's output, the "thinking brain" is functionally bypassed or overridden 1511. This lack of top-down inhibition directly accounts for the phenomenological experience of a "blank mind," wherein the individual is unable to recall rehearsed speech material because subcortical survival mechanisms have entirely superseded working memory retrieval and logical cognitive planning 1.
Default Mode Network and Self-Referential Processing
The psychological terror of glossophobia is deeply tied to an individual's perception of how they are being viewed, judged, and evaluated by an audience. This heightened, hyper-vigilant self-focus is intrinsically linked to altered functional connectivity within the Default Mode Network (DMN), a distributed brain network highly active during self-referential thought, internal reflection, and mental time travel 613. The posterior cingulate cortex (PCC) and the precuneus, which serve as primary nodes of the DMN, exhibit severe functional anomalies in socially anxious individuals during public speaking simulations 613.
During states of reflective self-appraisal - when individuals evaluate their own performance or anticipate the audience's judgment - individuals with social anxiety display greater excitatory connectivity between the PCC and the medial prefrontal cortex (mPFC), indicating an intense, anxiety-inducing preoccupation with the views others hold of them 13. Simultaneously, there is greater inhibitory connectivity from the left inferior parietal lobule (IPL) to the mPFC 13. This altered interplay between parieto-occipital areas and paralimbic structures generates a hypersensitivity to interoceptive cues. Consequently, when an individual notices their own racing heart, dry mouth, or flushed face, the DMN hyper-focuses on these physiological markers, which the brain subsequently misinterprets as further undeniable evidence of imminent social failure, thereby compounding the anxiety 614.
Salience Network and Interoceptive Sensitivity
In addition to the DMN, the Salience Network (SN) - comprising primarily the anterior insula and the dorsal anterior cingulate cortex (dACC) - plays a critical role in glossophobia. The SN is tasked with detecting and filtering salient, relevant stimuli from the environment and coordinating appropriate physiological responses 61516. Functional connectivity within the SN is notably altered even in subclinical, high-trait anxiety populations. Reduced connectivity between the right supramarginal gyrus and the anterior cingulate cortex has been observed, a decoupling that impairs the network's ability to appropriately value emotional stimuli 1516. Because the ACC is essential for optimizing behavior in response to negative emotional stimuli, reduced connectivity within the SN reflects the persistence of negative value judgments and an impaired capacity for emotion regulation 16.
Comparative Processing of Social and Physical Threats
Historically, the neural basis of threat perception was studied almost exclusively through the lens of physical danger, such as evasion of predators, response to physical pain, or avoidance of immediate bodily harm 2617. However, contemporary neuroimaging and high-resolution electrophysiological research confirms that social evaluative threats - like public speaking - command overlapping but highly distinct neural networks 51718. The brain does not merely equate a staring audience with a charging predator; rather, it attributes a highly sophisticated and specific processing architecture to social evaluation.
Evolutionary Origins of Social Evaluative Threat
The profound overlap between physical and social threat circuitry stems from the fact that both social exclusion and physical injury precipitate similar cognitive, affective, and behavioral consequences in human evolutionary history 23. Anthropological and social psychology models suggest that the social and physical pain systems co-evolved; ostracism from the group led to a lack of shelter, shared food, and protection, meaning social isolation was a proximate cause of physical death 23. Thus, when a speaker steps onto a stage and perceives an unresponsive or judgmental audience, the brain interprets this not as a minor social inconvenience, but as an acute threat to biological survival, triggering the exact same defensive survival behaviors necessary for predator evasion 126.
Distinctive Neural Signatures of Social Evaluation
While both physical and social threats elicit equivalent subjective ratings of negative emotion and rely fundamentally on the amygdala and basic autonomic arousal circuitry 25, profound neurological differences exist in how the human cortex handles these stimuli. During social threat processing, the brain exhibits an early, specialized processing advantage. Human sensory cortices begin assessing social threat features (such as facial micro-expressions in an audience) at roughly 160 milliseconds, a phenomenon evidenced by significantly increased amplitudes of the N170 component and the Early Posterior Negativity (EPN) in electroencephalographic (EEG) recordings 171920. This indicates that the brain prioritizes the visual processing of perceived social threats long before conscious cognitive evaluation takes place.
Furthermore, processing social evaluative threat relies heavily on midline cortical structures. Viewing social threats (e.g., harsh, rejecting faces typical of an unfriendly audience) compared to physical threats results in significantly greater blood-oxygen-level-dependent (BOLD) responses in the medial orbitofrontal cortex (mOFC), subgenual anterior cingulate cortex (sgACC), and bilateral parahippocampal gyrus 5. These regions are heavily implicated in complex emotion regulation, interpersonal cognition, and theory of mind.
Differences in Cognitive Regulation Strategies
The mechanisms utilized to regulate the fear response also diverge depending on the nature of the threat. When individuals attempt to cognitively regulate their fear in response to a physical threat, they rely on more fundamental cognitive control areas, specifically demonstrating heightened blood-oxygen-level-dependent response in the right dorsolateral prefrontal cortex (dlPFC) 5. Conversely, regulating a social threat demands greater integration of cognitive and attention regulation-related brain activation, relying less on the dlPFC and more heavily on medial prefrontal structures to process the complex interpersonal dynamics 5.
| Neurological Dimension | Physical Threat Processing (e.g., Predator, Bodily Harm) | Social Evaluative Threat Processing (e.g., Public Speaking) |
|---|---|---|
| Evolutionary Driver | Avoidance of immediate bodily injury or predation 16. | Avoidance of group exclusion and loss of social standing 123. |
| Core Subcortical Node | Amygdala, Thalamus 6. | Amygdala, Fusiform Face Area (FFA), Superior Temporal Gyrus 517. |
| Distinctive Cortical Activation | Dorsolateral Prefrontal Cortex (dlPFC) dominant during cognitive regulation attempts 5. | Medial Prefrontal Cortex (mPFC), Default Mode Network (PCC, Precuneus) heavily engaged 56. |
| Early EEG Signatures | Generalized arousal and fast wave activity. | Enhanced N170, N190, and EPN components, indicating preferential, rapid visual processing of faces 1719. |
| Endocrine Recovery Profile | Cortisol generally normalizes swiftly once physical danger physically exits the environment 421. | Cortisol recovery can be significantly delayed due to post-event rumination, shame, and cognitive replay of the evaluation 222. |
Temporal Dynamics of the Biological Stress Response
The biological sequence that occurs upon stepping onto a stage - or even anticipating doing so in the hours prior - unfolds across two highly coordinated, yet temporally distinct, neuroendocrine stages. This carefully orchestrated cascade serves to mobilize bodily resources, altering cardiovascular, respiratory, and cognitive functions to address the perceived threat 4232425.
Immediate Sympathetic-Adrenomedullary Activation
The initial, acute phase of the public speaking stress response is mediated by the sympathetic-adrenomedullary (SAM) system, which functions as the physiological "gas pedal" and initiates within milliseconds to seconds of the stressor's onset 5423. The moment the amygdala identifies the audience as a threat, it transmits distress signals to the hypothalamus, which immediately activates the sympathetic nervous system 54. Autonomic nerves transmit electrical signals directly to the adrenal medulla - the inner core of the adrenal glands located atop the kidneys - prompting the rapid secretion of catecholamines, primarily epinephrine (adrenaline) and norepinephrine (noradrenaline), into the bloodstream 5426.
The concentration of adrenaline spikes sharply at the onset of the stressor and peaks within one to five minutes, precipitating the immediate physical symptoms characteristic of severe stage fright 2125. This surge produces tachycardia (rapid heartbeat) and elevated arterial blood pressure to aggressively pump blood to major muscle groups, preparing the body for physical evasion 142627. Simultaneously, the bronchioles dilate, causing respiration to become shallow and rapid to maximize oxygen intake, while the pupils dilate to allow more light into the retina for enhanced threat detection 5427. To conserve energy for the impending physical exertion, non-essential systems like digestion and salivation are immediately halted, leading to the sensation of a dry mouth and gastrointestinal distress often described as "butterflies" 127. Furthermore, vasodilation in the skin causes facial flushing, and sweat glands activate to cool the body for anticipated action 12728.
While catecholamines cannot directly cross the blood-brain barrier, they exert profound cognitive effects via peripheral pathways that stimulate the brainstem's locus coeruleus. This structure subsequently triggers the release of noradrenaline and dopamine into the prefrontal cortex 25. This neurochemical flood temporarily heightens alertness, sharpens focus, and enhances short-term working memory - evidenced by an increased recency effect - during the immediate acute phase of the speech 425.
Sustained Hypothalamic-Pituitary-Adrenal Axis Engagement
Following the initial, instantaneous SAM activation, the body deploys a slower, sustained response via the hypothalamic-pituitary-adrenal (HPA) axis. This neuroendocrine network begins its activation roughly ten seconds post-threat but operates on a much slower timescale, resulting in a delayed hormonal curve that takes twenty to thirty minutes to reach peak concentrations in the bloodstream 232529.
In response to ongoing stress, the paraventricular nucleus of the hypothalamus releases corticotropin-releasing hormone (CRH) 2330. CRH travels via the hypophyseal portal system to the anterior pituitary gland, which responds by synthesizing and secreting adrenocorticotropic hormone (ACTH) into the systemic circulation 52330. ACTH ultimately reaches the adrenal glands, where it stimulates the zona fasciculata of the adrenal cortex to synthesize and release cortisol, the primary human glucocorticoid 52330.
| Response Parameter | Sympathetic-Adrenomedullary (SAM) System | Hypothalamic-Pituitary-Adrenal (HPA) Axis |
|---|---|---|
| Primary Physiological Function | Immediate "fight-or-flight" physical mobilization and arousal 423. | Sustained energy mobilization, metabolic regulation, and stress adaptation 52330. |
| Neurological Initiation Pathway | Amygdala → Hypothalamus → Autonomic Nerves 54. | Hypothalamus → Pituitary Gland 523. |
| Endocrine Effector | Adrenal Medulla 5. | Adrenal Cortex 523. |
| Primary Hormonal Output | Epinephrine (Adrenaline), Norepinephrine 426. | Glucocorticoids (primarily Cortisol) 2330. |
| Initial Time to Activation | Milliseconds to seconds 523. | Roughly 10 seconds to initiate chemical cascade 23. |
| Time to Peak Concentration | 1 to 5 minutes post-stressor 25. | 20 to 30 minutes post-stressor 2529. |
| Cognitive Consequences | Enhanced alertness, hypervigilance, rapid attention capture, increased working memory 425. | Altered memory consolidation, impaired long-term memory retrieval 25. |
Interaction Between Stress Axes and Memory Consolidation
The sustained neuroendocrine cascade initiated by the HPA axis profoundly alters cognitive processing, particularly within the hippocampal memory systems. Unlike adrenaline, cortisol is highly lipid-soluble and easily crosses the blood-brain barrier 25. Once in the brain, cortisol binds to two distinct receptor types: high-affinity mineralocorticoid receptors (MR) and lower-affinity glucocorticoid receptors (GR), both of which are densely populated in the hippocampus and prefrontal cortex 25.
During a severe episode of glossophobia, the massive influx of cortisol floods the lower-affinity GR receptors in the hippocampus. While minor, baseline elevations of cortisol can promote focus, the high concentrations elicited by intense social evaluative threat actively disrupt long-term memory retrieval 2531. This glucocorticoid-induced interference in the hippocampus explains the ubiquitous phenomenon wherein an anxious speaker suddenly forgets a well-practiced speech once on stage. The primacy effect of memory is suppressed by the cortisol peak approximately twenty minutes after the onset of the stressor 25. If the perceived social threat is not resolved through cognitive reappraisal or removal from the stage, the HPA axis continues to idle high, leading to persistent cortisol elevation, delayed physiological recovery, and potential allostatic overload 410.
Electrophysiological Biomarkers of Threat Perception
Recent advancements in biomedical engineering and clinical neuroscience have identified highly specific electroencephalogram (EEG) biomarkers associated with public speaking anxiety. These biomarkers capture the exact temporal and frequency dynamics of attentional bias - the involuntary tendency for socially anxious individuals to unconsciously fixate on threatening stimuli, such as a frowning audience member, rather than neutral cues.
Frontal Alpha Asymmetry and Approach-Avoidance Motivation
Social anxiety is reliably associated with heightened right Frontal Alpha Asymmetry (FAA). In neurophysiological terms, relatively greater right frontal brain activity correlates with withdrawal, avoidance motivation, and negative affect, whereas left frontal activity aligns with approach behavior and positive affect 323334. During public speaking anxiety assessments, individuals with high social anxiety display significantly higher right FAA compared to low-anxiety controls, serving as a reliable neural marker of their internal drive to flee the social evaluative context 323334. Furthermore, negative correlations have been identified between alpha power in the left cerebral region and attentional bias parameters, underscoring a suppression of approach-oriented cognitive control 3234.
Delta-Beta Correlation and Excitation-Inhibition Ratios
Heightened coupling between slow-wave (delta) and fast-wave (beta) activity in the frontal and central brain regions, known as Delta-Beta Correlation (DBC), has been documented in subjects with high public speaking anxiety 323334. This cross-frequency spectral coupling is hypothesized to reflect the intense, ongoing interaction between subcortical emotional processing (represented by delta oscillations) and cortical cognitive control efforts (represented by beta oscillations) 3234. Similarly, resting-state EEG studies analyzing the fractional Excitation-Inhibition (fEI) ratio reveal that individuals with moderate to severe social anxiety exhibit significantly higher fEI ratios in the alpha and low-beta bands, specifically within the prefrontal cortex 35. This dysregulation implies a state of cortical hyperexcitability, leaving the brain highly susceptible to minor social threat cues.
Event-Related Potentials and Ex-Gaussian Modeling
High social anxiety significantly modulates the early-stage perceptual encoding of social threat. Event-Related Potential (ERP) analyses indicate that socially anxious individuals show enhanced N170 amplitudes - reflecting intense attentional selection and discrimination in favor of threatening faces - and significantly more negative Early Posterior Negativity (EPN) compared to controls 1920. Furthermore, increased P200 amplitudes indicate a generalized early attentional sensitivity to threat orienting 2034. Because these potentials occur within 200 milliseconds of stimulus onset, it demonstrates that attentional biases in glossophobia emerge primarily during early perceptual encoding, meaning the brain prioritizes the visual processing of perceived social threats long before conscious evaluation is possible 20.
To quantify this attentional interference, researchers employ Ex-Gaussian modeling of reaction time data during emotional Stroop tasks. Individuals with public speaking anxiety demonstrate a significantly higher ex-Gaussian sigma value (indicating higher variability in the exponential tail of their reaction times) when processing emotional conditions 3234. This mathematical parameter acts as a robust performance biomarker, confirming that social threat words or imagery severely hijack the cognitive resources of an anxious speaker, causing pronounced delays in executive processing 3234.
Neuroendocrinological and Physiological Modulators
The magnitude of the stress response to public speaking is not static; it is heavily modulated by endogenous endocrinological factors, neurochemical baselines, and targeted pharmacological interventions.
Influence of Biological Sex and Ovarian Hormones
The neural connectivity and physiological reactivity governing social evaluative threat fluctuate significantly in response to biological sex and circulating ovarian hormones. During the mid-luteal phase of the menstrual cycle, when both progesterone and estradiol levels are elevated, the amygdala generally exhibits heightened reactivity to stress and negative social stimuli 3637. Furthermore, high luteal levels of ovarian hormones augment the overall sympathetic nervous system response, resulting in increased heart rate, diminished heart rate variability, and elevated noradrenergic activity during social stress compared to the follicular phase 37.
At the network level, ovarian hormones enhance functional connectivity between the salience network and the default mode network. This increased inter-network connectivity, combined with heightened stress responding, facilitates enhanced memory for unpleasant material, making negative social evaluations easier to recall and potentially increasing vulnerability to prolonged mood disturbances 37. Interestingly, long-term exposure to endogenous hormones (e.g., longer reproductive lifespan) appears to offer neuroprotective benefits regarding cerebral small vessel disease, though exogenous synthetic hormones (oral contraceptives) tend to suppress endogenous secretion and dramatically alter resting-state whole-brain connectivity in complex, cyclical patterns 3738.
Pharmacological Interventions and Autonomic Regulation
The autonomic severity of glossophobia is frequently mitigated through pharmacological interventions, most notably beta-adrenergic blocking agents such as propranolol. Propranolol competitively inhibits beta-adrenergic receptors, thereby blunting the peripheral effects of the sympathetic nervous system's catecholamine surge 39. In rigorous experimental paradigms utilizing psychosocial stressors like the Trier Social Stress Test, administration of propranolol successfully blunts pre-ejection period reactivity (a definitive marker of SNS activity on cardiac contractility) and reduces salivary alpha-amylase concentrations 39.
Crucially, while beta-blockade effectively attenuates the negative, high-arousal emotions (e.g., panic, trembling, extreme tachycardia) associated with the physical stress response, research demonstrates that cognitive appraisals of the threat remain entirely intact 39. The speaker still cognitively recognizes the social evaluative threat of the audience, but their physiological feedback loop is severed, preventing the physical symptoms of anxiety from spiraling into a debilitating panic attack 39.
Oxytocin and Social Learning
The neuropeptide oxytocin, widely recognized for its role in social bonding and affiliation, also acts as a powerful modulator of the neural circuits processing social threat. Intranasal administration of oxytocin during social learning tasks yields profound behavioral and neural changes that are heavily dependent on an individual's baseline social anxiety and biological sex 4041. In high-anxiety individuals, oxytocin administration increases subjective sympathy ratings for faces and alters objective physiological arousal (measured via pupil dilation) 40. Furthermore, oxytocin dynamically modulates neural activation in the insula - a key region of the salience network - decreasing activation in low-anxious individuals while paradoxically increasing it in highly anxious individuals 40. This reveals a highly nuanced, context-dependent neurochemical landscape where the brain's response to social evaluation is continuously mediated by the interplay of baseline trait anxiety, sex, and neuropeptide concentrations.
Cross-Cultural Variances in Social Evaluative Threat
The neurobiological baseline and intensity of social evaluative threat are not uniform across all global populations; they are significantly modulated by cultural frameworks. Specifically, the socio-cognitive dimension of individualism versus collectivism dictates how the brain structurally represents the self and consequently determines the physiological magnitude of the public speaking stress response 42.
Individualism, Collectivism, and the Structural Self
Individualistic cultures (e.g., the United States, Germany, Australia) heavily emphasize uniqueness, personal autonomy, and an independent view of the self 34643. In contrast, collectivistic cultures (e.g., Japan, China, South Korea) prioritize group harmony, interdependence, modesty, and a relational view of the self 34643. Cross-cultural fMRI neuroimaging studies reveal that these values fundamentally alter self-representation in the brain. Individuals from highly individualistic backgrounds show significantly greater medial prefrontal cortex (MPFC) activation during general, context-free self-judgments (e.g., "I am honest"). Conversely, individuals from collectivistic backgrounds exhibit greater MPFC activation when processing contextual self-judgments (e.g., "When talking to my mother, I am honest"), indicating that the collectivistic neural architecture inherently defines the self in relation to social surroundings 4244.
Differences in Autonomic and Glucocorticoid Reactivity
These deep structural differences in self-representation profoundly impact physiological stress reactivity. Studies measuring cardiovascular and endocrine responses to social exclusion paradigms (e.g., Cyberball tasks) reveal that individuals from individualistic cultures suffer significantly greater physiological distress than those from collectivistic cultures 3. Because the individualistic self is rooted in autonomy, explicit social exclusion or negative evaluation registers as a severe, direct threat to self-integrity. This precipitates an immediate autonomic reaction, evidenced by significant increases in heart rate and a robust HPA axis response 3.
Conversely, the collectivistic self, being structurally interdependent and less ego-focused, appears to be less susceptible to sudden, isolated threats to individual belonging. Consequently, collectivists often display blunted physiological stress profiles - lacking immediate cardiovascular reactivity - during similar social exclusion and evaluative tasks 3.
Social Norms and the Etiology of Anxiety
Despite demonstrating less physiological reactivity to isolated exclusion, this does not imply that collectivist societies are immune to social anxiety. In fact, large-scale epidemiological and network-analysis data indicate significantly higher baseline levels of self-reported social anxiety and fear of negative evaluation in collectivistic nations 4345. This paradox is driven by the stringent societal norms regarding appropriate social behavior; the pressure to maintain group harmony creates an environment where deviating from emotion norms or violating social expectations carries immense cultural weight 4546.
The neurochemical manifestations simply map onto different contextual triggers based on the culture's values. For instance, individuals from individualist cultures secrete higher levels of the stress hormone cortisol when forced to talk about the influence others have had on them, as this threatens their deeply held neural concept of autonomy 51. In stark contrast, individuals from collectivist cultures secrete higher levels of cortisol when forced to talk about their own influence on others, as this self-aggrandizement threatens cultural norms of modesty and group harmony 51. Thus, while the underlying neurobiology of the HPA and SAM axes remains universal, the specific social stimuli required to trigger the threat response are entirely dictated by the cultural programming of the prefrontal cortex.
Neurobiological Mechanisms of Treatment and Extinction
The standard, most effective clinical intervention for specific phobias, including severe glossophobia, is exposure therapy - a cognitive-behavioral process involving gradual, repeated confrontation with the feared stimulus, such as simulated public speaking 524748. The precise neurobiological mechanism underlying the efficacy of this treatment is an area of intense, ongoing clinical research, defined largely by the debate between two competing paradigms: the traditional habituation model and the contemporary inhibitory learning model.
Habituation Theory Versus Inhibitory Learning Models
Historically, the habituation model posited that therapeutic success required a literal desensitization of the fear circuitry. Under this framework, effective exposure therapy would manifest as a gradual reduction in amygdala activation over the course of the exposure sessions 5247. Theoretically, successful treatment of public speaking anxiety would mean the amygdala physically ceases to perceive the audience as a threat, resulting in neural silence.
However, advanced fMRI data directly challenges this assumption. Evidence indicates that amygdala habituation - a decrease in BOLD activity during an exposure session - is actually associated with a failure to reduce phobic symptoms, potentially signaling cognitive detachment or dissociation rather than healing 5247. Instead, sustained or even escalating amygdala activation during the exposure process strongly correlates with superior clinical improvement 52.
This data strongly supports the inhibitory learning model. This model theorizes that exposure therapy does not erase, overwrite, or habituate the original fear memory. Instead, it requires the active, high-arousal engagement of the amygdala to forge entirely new, non-fearful associations alongside the old ones 47. Over time, these new safety associations are consolidated by the ventromedial prefrontal cortex (vmPFC) and the hippocampus, which subsequently project powerful inhibitory signals back to the amygdala to regulate future responses during extinction recall 911. Consequently, the ultimate neurobiological goal of exposing individuals to public speaking is not to eradicate the brain's perception of threat, but rather to aggressively strengthen the prefrontal cortex's capacity to tolerate the stimulus and down-regulate the ensuing autonomic storm.
Post-Treatment Functional Connectivity Restructuring
The successful application of these cognitive behavioral interventions results in profound, measurable restructuring of the brain's resting-state networks. Connectome-wide functional connectivity multivariate pattern analyses (fc-MVPA) of individuals before and after one-session exposure treatments reveal that the brain physically rewires how it handles threat 48. Prior to treatment, socially anxious individuals show severe anticorrelations between sensorimotor regions, the frontoparietal networks, and temporal nodes of the default mode network. Following successful exposure therapy, the connectivity strengths between almost all identified nodes return to levels comparable to healthy, non-phobic control groups 48. This large-scale network reintegration indicates improved cortical action control, enabling the individual to accurately predict their environment and establish a stronger, healthier coupling between the brain's perceptual areas and its higher-order prefrontal control regions 48.