Neurobiological mechanisms of memory consolidation during sleep
Sleep is a fundamental biological imperative governing central nervous system homeostasis. Far from being a passive state of rest, sleep is a highly dynamic and structured neurophysiological process essential for cognitive function, memory consolidation, and metabolic clearance. Modern neuroscientific investigations have delineated how precise electrical oscillations during sleep transform transient experiences into durable memory traces, while concurrent mechanical fluid dynamics protect the brain against the accumulation of neurotoxic proteins. Alterations in these dual processes dictate the trajectory of learning, performance, and cognitive aging.
This report provides an exhaustive analysis of the neuroscientific mechanisms driving sleep-dependent memory consolidation, the competing models of cerebral extracellular waste clearance, the cognitive consequences of acute and chronic sleep restriction, and the evolutionary baselines of human sleep architecture.
Sleep Architecture and Memory Processing
Sleep is cyclically organized into Rapid Eye Movement (REM) sleep and Non-Rapid Eye Movement (NREM) sleep, which alternate in approximately 90-to-120-minute intervals 11. Healthy adult sleep architecture comprises progressively deeper stages of NREM sleep (N1, N2, and N3), followed by episodes of REM sleep that typically lengthen as the sleep period progresses 11. Each stage facilitates distinct mechanisms of memory processing and physiological restoration.
NREM sleep, accounting for 75% to 80% of total sleep time, is critical for declarative and spatial memory consolidation 12. Stage N2 is characterized by the presence of sleep spindles and K-complexes, while Stage N3 - often referred to as slow-wave sleep (SWS) - is dominated by high-amplitude, low-frequency delta waves 11. Conversely, REM sleep comprises 20% to 25% of total sleep and is characterized by rapid eye movements, muscle atonia, and fast, desynchronized electroencephalographic (EEG) activity resembling wakefulness 12. REM sleep is heavily implicated in emotional memory processing, procedural skill consolidation, and the integration of novel memory traces into existing cognitive schemas 34.
Mechanisms of NREM Sleep Consolidation
The transformation of labile, newly encoded information into stable, long-term memory traces is best understood through the active systems consolidation theory. This framework posits that memories temporarily stored in the hippocampus are repeatedly reactivated during NREM sleep and gradually redistributed to neocortical networks for long-term storage 56. This process is entirely dependent on the precise hierarchical coupling of three distinct neural oscillations.
Thalamocortical Slow Oscillations and Sleep Spindles
The deepest stage of NREM sleep is driven by neocortical slow oscillations (SOs). These SOs are high-amplitude, low-frequency (< 1 Hz) fluctuations in membrane potentials that travel as waves primarily from the prefrontal cortex toward posterior regions 57. The SO represents a global synchronization of neural networks, toggling between hyperpolarized down-states (neuronal silence) and depolarized up-states (intense neuronal firing) 578.
During the depolarized up-state of the SO, corticothalamic projections stimulate the thalamic reticular nucleus. This stimulation triggers the generation of sleep spindles - waxing and waning bursts of oscillatory activity in the 11 to 16 Hz range. These spindles propagate widely back to the neocortex through synchronized thalamocortical projections 5910. Fast sleep spindles (12.5 - 16 Hz) are particularly crucial, as they synchronize cortical activity and establish a correlated activation between the hippocampus and frontal brain regions 11.
Hippocampal Sharp-Wave Ripples and Neural Replay
The actual content of the memory being consolidated is carried by hippocampal sharp-wave ripples (SWRs). SWRs are brief, high-frequency oscillatory bursts (typically 80 - 140 Hz) generated within the hippocampus 1112. These ripples represent the compressed, high-speed neural replay of spatial and episodic firing patterns that initially occurred during prior waking experiences 1213.
Recent intracranial electroencephalography (iEEG) studies involving neurosurgical epilepsy patients performing serial recall tasks have provided high-resolution, direct evidence of this phenomenon in the human brain. These recordings demonstrate that the temporal coupling of ripples between the hippocampus and extrahippocampal areas, such as the orbitofrontal cortex and posterior cingulate cortex, is significantly greater than chance during successful memory encoding and retrieval 1215.
Phase-Amplitude Coupling and Synaptic Plasticity
The success of active systems consolidation relies entirely on phase-amplitude coupling. For the memory trace to be successfully transferred and permanently stored, the hippocampal SWR must nest precisely within the excitable trough of the thalamic sleep spindle, which must in turn be locked to the peak of the neocortical slow oscillation up-state 5711.
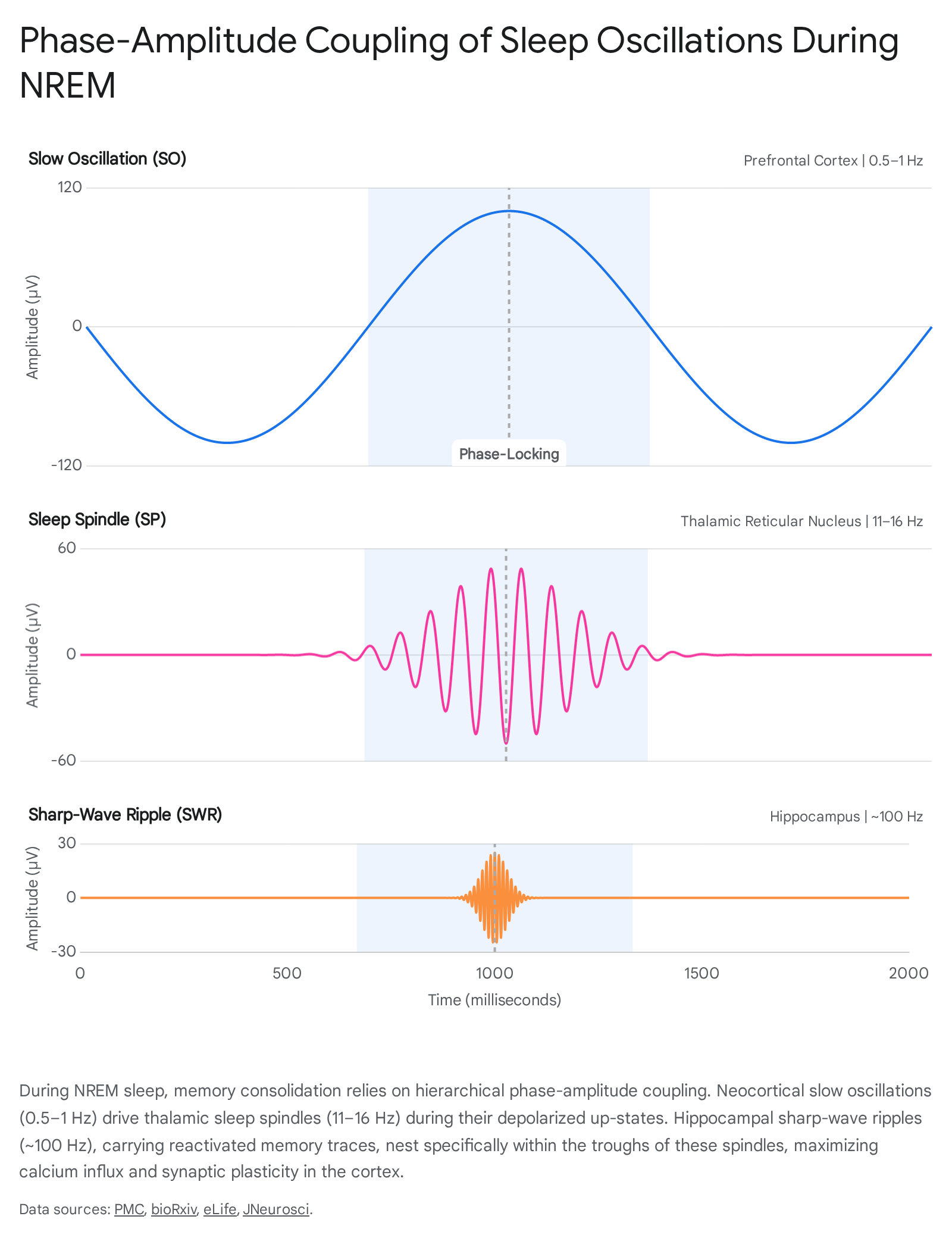
This tripartite temporal alignment establishes an optimal physiological environment for spike-timing-dependent plasticity (STDP). When these rhythms align, spindle-driven spike discharges induce a massive influx of calcium (Ca2+) into excitatory neocortical synapses 5911. The sequential coupling of SOs, spindles, and ripples leads to a stepwise increase in neuronal firing rates and short-latency cross-correlations among local neuronal assemblies, maximizing the efficiency of synaptic potentiation 710.
The Role of REM Sleep in Memory Processing
While NREM sleep facilitates the systemic consolidation of declarative memories, REM sleep presents an entirely different neurochemical and oscillatory profile. REM sleep is characterized by prominent theta activity (4 - 7 Hz), which describes low-frequency oscillations in the local field potentials within the hippocampus, amygdala, and neocortex 34.
Emotional Regulation and Theta Coherence
REM sleep is pivotal for emotional memory consolidation. Theta coherence between the hippocampus and amygdala drives large-scale pontine-geniculo-occipital (PGO) waves, the density of which predicts increases in plasticity-related gene expression 34. Furthermore, the neuromodulatory milieu of REM sleep is unique, featuring high cholinergic (acetylcholine) activity and minimal noradrenergic activity. High acetylcholine levels are thought to reduce hippocampal afferent inputs to the neocortex, functionally disengaging the hippocampus and allowing the neocortex to integrate previously consolidated memory traces without interference 414.
Clinical and experimental data underscore the adaptive role of REM theta activity in processing traumatic experiences. Elevated theta activity during REM sleep following exposure to emotionally negative or traumatic stimuli is highly predictive of reduced subsequent intrusive memories and attenuated negative affect. Conversely, individuals suffering from post-traumatic stress disorder (PTSD) frequently exhibit pronounced deficits in REM sleep architecture, impairing this adaptive emotional processing 1516.
Representational Drift and Memory Integration
Recent neurophysiological studies have illuminated the distinct, often opposing roles of NREM and REM sleep in memory "representational drift." Representational drift refers to the gradual shift in neuronal activity patterns that occurs over time as memories adapt to new contexts or generalize to fit existing schemas 17.
A landmark 2025 study tracking hippocampal CA1 neuronal assemblies over 20-hour periods in rats revealed that reactivated assembly patterns reorganize spontaneously during prolonged sleep 13. Strikingly, NREM and REM sleep exert antagonistic effects on this phenomenon. NREM sleep actively promotes and accelerates representational drift, facilitating the integration and updating of the memory into broader, generalized neocortical networks 1720. Conversely, REM sleep counters this drift, acting to preserve and stabilize the original high-fidelity memory trace 1317. This challenges older hypotheses positing that REM sleep functions primarily to prune or forget unimportant information. Instead, the precise REM-to-SWS ratio dictates the balance between memory transformation (generalization) and preservation (fidelity) 1718.
Functional Signatures of NREM versus REM Sleep
| Parameter | NREM Sleep (Slow-Wave Sleep) | REM Sleep (Paradoxical Sleep) |
|---|---|---|
| Dominant Oscillations | Slow Oscillations (<1 Hz), Spindles (11-16 Hz), Ripples (80-140 Hz) | Theta Activity (4-7 Hz), Beta Activity (15-25 Hz) |
| Primary Neurochemistry | Reduced acetylcholine, moderate noradrenaline | High acetylcholine, minimal noradrenaline |
| Memory Domain | Declarative, spatial, systems consolidation | Emotional, procedural, trace integration |
| Representational Impact | Accelerates representational drift (generalization) | Counters drift (preserves original fidelity) |
| Clinical Relevance | Disrupted in normal aging, impairs declarative recall | Disrupted in PTSD, impairs trauma regulation |
Behavioral Manipulation of Memory Consolidation
The discovery that memories are spontaneously reactivated during sleep has driven the development of targeted bioengineering techniques to augment this process. The most thoroughly validated approach is Targeted Memory Reactivation (TMR).
Targeted Memory Reactivation
Targeted Memory Reactivation is a non-invasive tool designed to manipulate and enhance memory consolidation. In TMR paradigms, specific learning content is paired with sensory cues, such as discrete auditory tones or olfactory stimuli, during waking encoding. During subsequent NREM sleep, the same cues are presented at a sub-awakening intensity threshold to selectively trigger the neural replay of the associated memory trace 192021.
TMR must be distinguished from the concept of hypnopaedia (sleep learning), which attempts to introduce entirely novel information to a sleeping subject. TMR relies strictly on reinforcing preexisting memory networks 2022. When TMR auditory cues are administered, electroencephalography reveals a distinct neurophysiological signature: successful reactivation is associated with an event-related increase in theta oscillations (4 - 7 Hz) occurring 500 to 1200 milliseconds post-cue, immediately followed by an induced burst of sleep spindle oscillations (11 - 16 Hz) 1922. This artificially induced sequence mimics the endogenous SO-spindle-ripple complex, amplifying consolidation.
Clinical and Home-Based Applications
Recent engineering advancements have transitioned TMR from highly controlled polysomnography laboratories to automated, in-home applications. Wearable systems, such as the SleepStim platform, utilize actigraphy and heart rate data processed by machine-learning models to reliably identify deep NREM sleep. The system pairs with a smartphone to emit auditory cues automatically 2324. Studies validating these systems confirm that automated TMR effectively enhances spatial memory, provided the auditory stimulus intensity remains strictly below the arousal threshold; high-intensity cues disrupt sleep architecture and negate memory benefits 2324.
Clinically, TMR is being actively investigated for accelerating vocabulary acquisition, aiding motor skill rehabilitation following stroke, and modulating maladaptive emotional memories in affective disorders and PTSD 2125. However, results in certain populations, such as motor rehabilitation in older adults, remain mixed, likely due to underlying age-related alterations in baseline sleep architecture 21.
Cerebral Extracellular Waste Clearance
Beyond the electrical consolidation of memories, sleep fulfills a vital mechanical function: the large-scale clearance of metabolic waste from the brain parenchyma. The high metabolic rate of cerebral neurons generates vast quantities of interstitial by-products, including amyloid-beta (Aβ), tau proteins, and lactate. The pathological accumulation of these neurotoxic proteins is a fundamental mechanism of cognitive aging and neurodegenerative disorders such as Alzheimer's disease (AD) 262728. Because the central nervous system lacks a traditional lymphatic capillary network within the parenchyma, it relies on specialized fluid dynamics for homeostasis.
The Glymphatic System
The most prominent framework for brain clearance is the glymphatic (glial-lymphatic) system. This macroscopic pathway utilizes cerebrospinal fluid (CSF) to wash the brain tissue. The glymphatic process involves three sequential steps: 1. Periarterial Influx: CSF from the subarachnoid space flows rapidly into the brain parenchyma along the perivascular spaces (Virchow-Robin spaces) surrounding penetrating cerebral arteries 283229. 2. Parenchymal Exchange: CSF is driven into the interstitial space, where it mixes with interstitial fluid (ISF). This fluid exchange is heavily dependent on aquaporin-4 (AQP4) water channels, which are densely concentrated on the astrocytic endfeet that ensheathe the cerebral microvasculature 283230. 3. Perivenous Efflux: The resulting mixture of CSF, ISF, and metabolic waste flows toward the perivenous spaces, eventually exiting the brain via meningeal lymphatic vessels located in the dura mater, which connect to the peripheral cervical lymph nodes 2731.
Glymphatic function is highly state-dependent, operating primarily during sleep. Transitioning from wakefulness to sleep results in a roughly 60% expansion of the extracellular space volume due to the shrinkage of glial cells. This expansion dramatically lowers parenchymal hydraulic resistance, increasing the clearance of neurotoxins like amyloid-beta by over 60% compared to waking states 2628.
The motive force for this clearance is intrinsically linked to neurophysiology. In 2024, distinct research groups demonstrated that the highly synchronized neural activity characteristic of slow-wave sleep is responsible for propelling this fluid. As large populations of neurons fire and cease firing simultaneously during slow oscillations, they act as miniature biological pumps. This synchronized activity drives rhythmic vascular motion (vasomotion) that propels fluid through dense brain tissue 3233.
Furthermore, human in vivo evidence of the glymphatic system was definitively established in late 2024. Utilizing gadolinium-based intrathecal contrast agents and specific fluid-attenuated inversion recovery (FLAIR) MRI protocols in neurosurgical patients, researchers directly visualized CSF moving selectively through distinct perivascular channels in the living human brain, confirming decades of murine models 343536.
Competing Models: IPAD and BBB Direct Transport
While the glymphatic hypothesis is widely accepted, it exists alongside competing models of brain clearance, resulting in ongoing debate within the neurovascular community.
Intramural Periarterial Drainage (IPAD): The IPAD model proposes an anatomically distinct efflux pathway. Instead of waste exiting via perivenous spaces, the IPAD hypothesis suggests that ISF and soluble amyloid-beta are driven out of the brain along the basement membranes of capillaries and the tunica media of arteries 3237. Crucially, this drainage occurs counter-current to arterial blood flow. The motive force for IPAD is the contraction of vascular smooth muscle cells 323738. Proponents argue that IPAD better accounts for the neuropathological presentation of Cerebral Amyloid Angiopathy (CAA), where Aβ deposits are found heavily in arterial walls rather than venous networks 3237.
Blood-Brain Barrier (BBB) Transvascular Exchange: A third pathway involves direct clearance across the BBB. This mechanism uses selective, receptor-mediated transport - specifically the low-density lipoprotein receptor-related protein 1 (LRP1) and the receptor for advanced glycation end products (RAGE) - to move Aβ directly from the interstitial space into the systemic bloodstream 2939.
Modern computational models, such as Lattice Boltzmann Method (LBM) simulations, suggest these systems are not mutually exclusive but operate synergistically. Glymphatic flow provides bulk advection of solutes, while BBB transport provides localized, receptor-mediated clearance, and IPAD manages specific intramural efflux 2939.
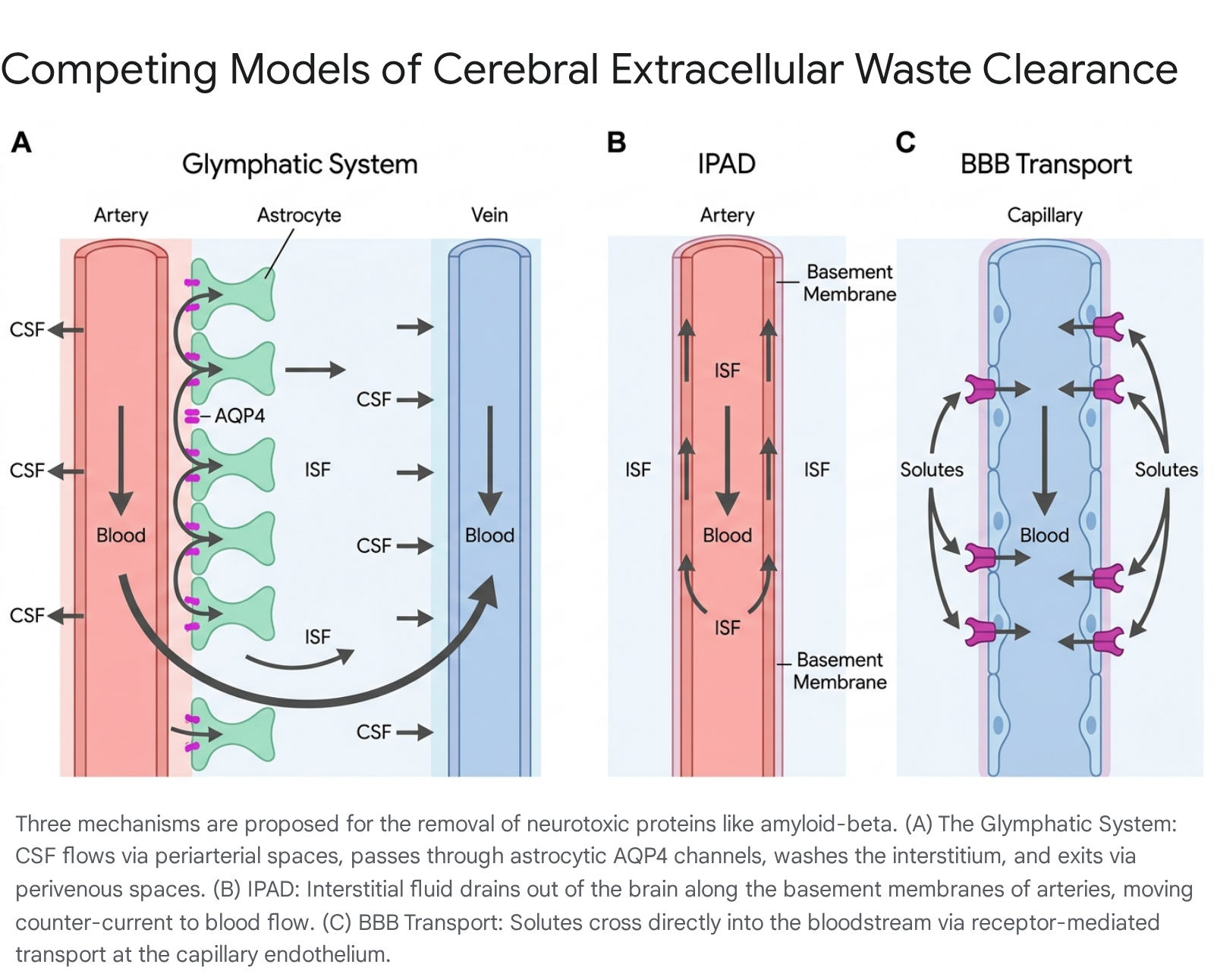
Comparative Models of Extracellular Solute Efflux
| Clearance Model | Primary Anatomical Route | Directionality and Mechanism | Primary Physiological Drivers | Key Molecular Mediators |
|---|---|---|---|---|
| Glymphatic System | Periarterial influx to perivenous efflux. | Forward bulk advection connecting to meningeal lymphatics. | Synchronized slow oscillations, arterial pulsatility. | Aquaporin-4 (AQP4) water channels. |
| IPAD Pathway | Basement membranes of capillaries and the tunica media. | Counter-current efflux opposite to arterial blood flow. | Vascular smooth muscle cell contraction (vasomotion). | Basement membrane proteins (Laminin, Collagen IV). |
| BBB Transvascular | Capillary endothelial cells. | Direct transport from interstitium into systemic circulation. | Concentration gradients and receptor binding. | LRP1, RAGE, ABCA1 receptors. |
Cognitive and Physiological Consequences of Sleep Restriction
The neurophysiological mechanisms of memory consolidation and mechanical waste clearance are highly susceptible to sleep disruption. Failure to achieve adequate sleep architecture profoundly impacts both immediate cognitive performance and long-term neurocognitive health.
Acute Deprivation Versus Chronic Restriction
Scientific literature historically focused on acute total sleep deprivation (TSD), such as 24 to 72 hours of prolonged wakefulness. However, modern society more frequently normalizes chronic partial sleep restriction (CSR) - obtaining 4 to 6 hours of sleep per night over consecutive days 40.
A comprehensive 2024 meta-analysis encompassing 147 studies and thousands of participants conclusively demonstrated that even moderate CSR produces severe, dose-dependent cognitive deficits. Attention and vigilance exhibit the largest effect sizes, with performance on tasks like the Psychomotor Vigilance Task (PVT) decreasing by 20% to 50% 45. The analysis revealed that subjective sleepiness increases significantly even after a single night of restricted sleep (Standardized Mean Difference = 0.986), alongside significant impairments in sustained attention 41. Working memory capacity, crucial for complex cognitive tasks, also suffers significant 15% to 30% impairments following CSR 4542. The overall cognitive dysfunction induced by accumulating sleep debt is comparable to legal levels of alcohol intoxication, severely impacting occupational safety and academic performance 45.
Neurophysiological Markers of Sleep Loss
The brain under mild sleep restriction exhibits distinct neurophysiological signatures. Resting-state EEG reveals an increase in low-frequency bands - specifically delta and theta power - which are primary markers of drowsiness that replace normal resting alpha activity ("alpha power dropout") 43.
Furthermore, EEG microstate analysis, which maps the spatial and temporal characteristics of resting brain networks, reveals significant topographical changes following sleep restriction. Studies show a significant increase in the occurrence of Microstates C and D following sleep loss. The increased presence of Microstate D correlates directly with disrupted phonological information processing and altered attention resource allocation 44.
Sleep restriction also alters functional connectivity and metabolic regulation. Functional MRI (fMRI) studies show that even a mild 1.5-hour delay in bedtime upregulates neural reward networks - specifically the right insula and inferior frontal gyrus - in response to food stimuli, providing a direct neurobiological mechanism for the established epidemiological link between short sleep duration, metabolic disruption, and obesity 45.
Evolutionary Baselines and Metabolic Health
The widespread assumption that modern industrial lifestyles and artificial lighting have artificially reduced human sleep duration from an ancestral 8-to-9-hour baseline is frequently challenged by contemporary anthropological data.
Pre-Industrial Sleep Patterns
Extensive studies monitoring geographically isolated hunter-gatherer and pre-industrial societies - the Hadza of Tanzania, the San of Namibia, and the Tsimane of Bolivia - reveal that these populations sleep an average of only 5.7 to 7.1 hours per night 46524748. Objective actigraphy data show that these ancestral proxy groups do not go to sleep at sunset. Instead, they remain awake for an average of 3.3 hours after dusk. Their sleep onset is strongly governed by environmental thermoregulation rather than light alone; they fall asleep as ambient temperatures drop and consistently wake just before dawn when temperatures reach their nadir 4649.
Intriguingly, while total sleep duration is shorter in these pre-industrial groups, recent comparative data from 54 global populations suggest that modern humans in industrial societies actually spend more time in bed and have a higher sleep efficiency (87.9% versus 73.9% in non-industrial groups) 5051. The primary modern sleep deficit is therefore not necessarily total duration, but rather profound circadian disruption caused by the decoupling of internal biological clocks from natural temperature and light cycles 4650.
Mortality Associations and Metabolic Markers
Despite shorter sleep durations, hunter-gatherer populations exhibit minimal rates of clinical insomnia (so rare that their native languages often lack a specific word for the disorder) and possess excellent cardiovascular and metabolic health 4849.
However, in modern industrial populations, epidemiological data consistently exhibit a distinct U-shaped mortality curve associated with sleep duration. Both severe short sleep (≤5 hours) and excessively long sleep (>8 to 9 hours) are associated with elevated risks of all-cause mortality, cardiovascular disease, and stroke 525354. Short sleep disrupts metabolic homeostasis, correlating significantly with elevated fasting blood glucose, increased insulin resistance, and elevated inflammatory markers such as C-reactive protein (CRP) and Interleukin-6 (IL-6) 536155. Conversely, the mortality risk associated with excessively long sleep is heavily mediated by underlying health status, depression, and low physical activity rather than the sleep itself 5354.
Sleep Parameters: Industrial vs. Pre-Industrial Societies
| Parameter | Pre-Industrial Societies (e.g., Hadza, San) | Industrial Societies |
|---|---|---|
| Average Sleep Duration | 5.7 to 7.1 hours | 7.1 to 8.0 hours |
| Sleep Efficiency | ~73.9% | ~87.9% |
| Primary Sleep Regulators | Ambient temperature drops, natural light | Artificial lighting, occupational schedules |
| Insomnia Prevalence | Near zero (often undefined in native language) | 10% to 30% |
| Metabolic Health Impact | High fitness, low baseline inflammation | U-shaped risk curve for mortality/inflammation |
Aging, Cognitive Decline, and Pathological Manifestations
The aging brain experiences a compounding failure in both the electrical and mechanical functions of sleep, directly accelerating cognitive decline and neurodegeneration.
Oscillatory Misalignment
Structurally, normal aging is accompanied by gray matter atrophy in the medial prefrontal cortex, which severely disrupts the generation of slow oscillations. Consequently, older adults experience a dramatic reduction in deep slow-wave sleep 11.
More critically, aging deteriorates the precise phase-amplitude coupling required for memory consolidation. While healthy young adults exhibit precise nesting of fast spindles (12.5 - 16 Hz) within the exact peak of the SO up-state, older adults often show dysregulated coupling. In the aging brain, slow spindles (9 - 12.5 Hz) increasingly couple at the end of the SO up-state 11. This temporal misalignment means the hippocampus replays information outside the optimal window for calcium influx, preventing effective hippocampo-neocortical communication. This uncoupling correlates directly with age-related deficits in overnight associative memory retention 11.
Mechanical Clearance Failure
Simultaneously, the brain's mechanical clearance mechanisms fail. The glymphatic system's efficiency relies heavily on the strict polarization of AQP4 channels at the astrocytic endfeet. In aging and neurodegenerative diseases, astrocytes become reactive, and AQP4 depolarizes, distributing randomly across the astrocyte soma. This loss of polarity dramatically increases hydraulic resistance, choking off the perivascular clearance of CSF and ISF 2856.
Furthermore, arteriosclerosis and the age-related stiffening of cerebral arterial walls reduce the vascular pulsatility that provides the kinetic driving force for both glymphatic influx and IPAD efflux 57. The resulting accumulation of amyloid-beta in the perivascular spaces further obstructs flow. Radiologically, this manifests as enlarged perivascular spaces (EPVS) on MRI, which correlate strongly with white matter lesions, amyloid pathology, and advanced cognitive impairment 2857. This creates a self-amplifying cycle: protein accumulation impairs clearance channels, which in turn accelerates further neurotoxic buildup, culminating in the progressive cognitive decline characteristic of Alzheimer's disease.
Bioengineering efforts are actively exploring interventions to mitigate this mechanical decline. Emerging therapies such as Transcranial Radiofrequency Treatment (TRFT) are being investigated for their ability to upregulate Vascular Endothelial Growth Factor (VEGF) in Alzheimer's patients, ostensibly rejuvenating meningeal lymphatic flow and enhancing brain toxin clearance even during wakefulness 5859.