Hormesis in exercise, fasting, cold exposure, and longevity
Quantitative Characteristics of Hormetic Dose-Response Models
The hormesis principle describes a highly conserved biological phenomenon characterized by a biphasic dose-response relationship, wherein low-dose exposure to an environmental agent or stressor induces a beneficial, adaptive response, while high-dose exposure results in toxicity, inhibition, or cellular damage 21. Historically marginalized in favor of linear no-threshold or strict threshold models, modern toxicological and pharmacological databases now establish hormesis as a fundamental biological expectation. The phenomenon is highly generalizable, occurring across plants, viruses, bacteria, fungi, invertebrates, and mammalian species, and applies to a broad spectrum of chemical classes and physical stressors 234.
Parameters of the Biphasic Curve
The quantitative features of the hormetic dose-response curve are remarkably consistent regardless of the biological system, the specific stressor, or the endpoint measured 15. The idealized biphasic curve is divided into distinct functional zones. The initial range below the response threshold is the lag zone, where no observable effects are registered 6. This is followed by the hormetic stimulatory zone, a region of positive adaptation that sits immediately contiguous with the toxicological No Observed Adverse Effect Level (NOAEL) 26. Beyond the NOAEL lies the Zero Equivalent Point (ZEP), where the biological response crosses the baseline control level and enters the inhibitory or toxic phase 67.
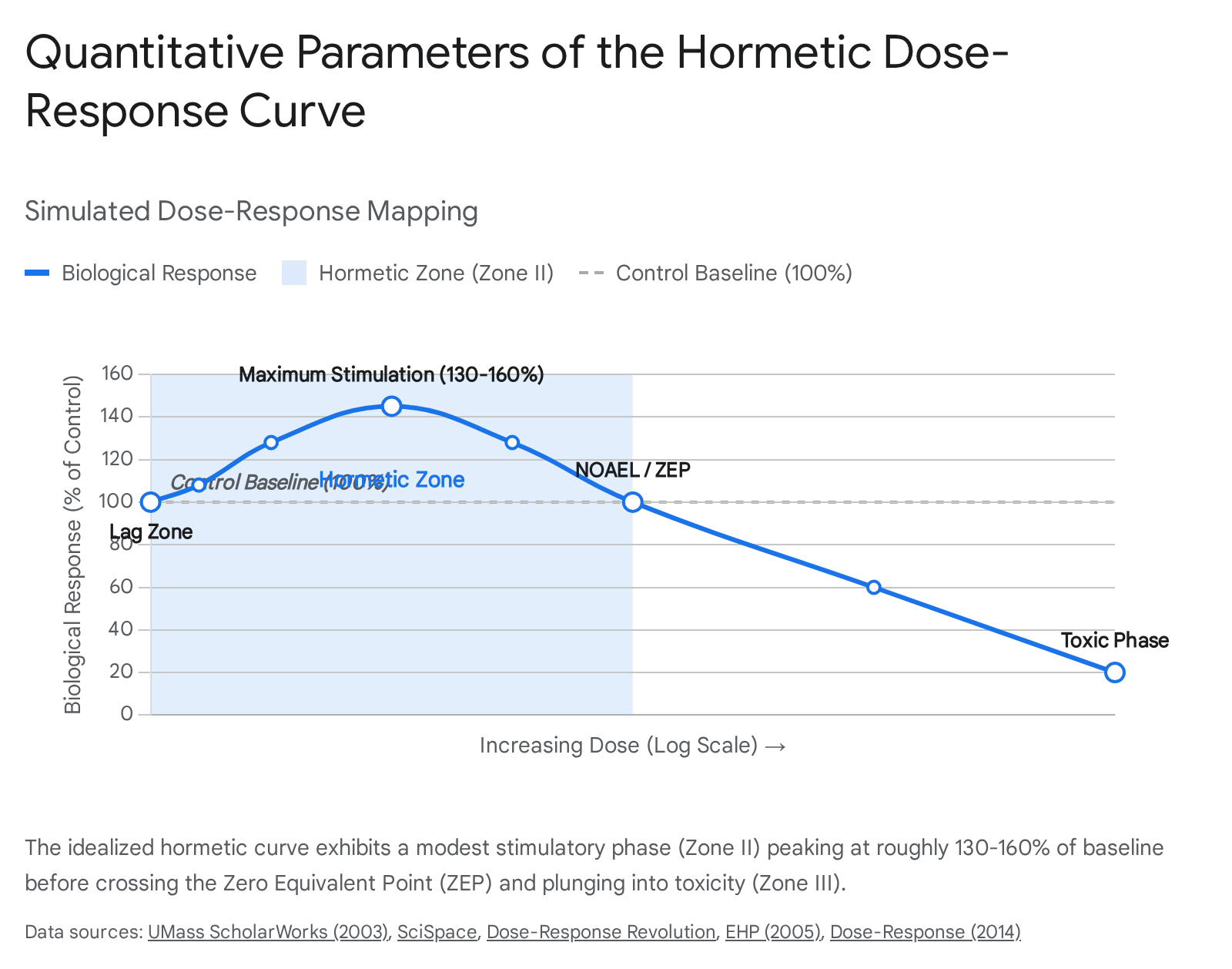
Evaluations of comprehensive relational retrieval databases containing several thousand dose-response datasets reveal that the amplitude of the hormetic response is inherently modest. The maximum stimulatory response rarely exceeds a twofold increase over the control group, typically manifesting at 30% to 60% above baseline values 257. The width of this stimulatory dose range is similarly constrained. It generally extends across a 10-fold to 20-fold dose range and only rarely exceeds a 1,000-fold range 25. This precise quantitative constraint represents the strict limits of phenotypic plasticity and underscores the fallacy of the conventional assumption that higher doses of a beneficial stressor will yield proportionally greater physiological returns 15.
Evaluative Criteria and Algorithm Development
To rigorously assess the presence of hormesis in toxicological literature, researchers have developed a priori evaluative criteria based on study design characteristics. These criteria evaluate whether an experiment displays a clear NOAEL, the number of test doses below the NOAEL, the magnitude of the low-dose stimulation, the statistical significance of the stimulatory effects, and the reproducibility of the findings 22. These variables are synthesized into mathematical algorithms that estimate the likelihood of a dose-response relationship fitting the hormetic hypothesis 22. Analyses utilizing these algorithms consistently demonstrate that responses at sub-NOAEL dosages are not randomly distributed fluctuations, but rather represent systematic, low-dose overcompensation to disruptions in homeostasis 2.
Molecular Mediators of the Hormetic Response
The systemic benefits of environmental stressors - such as physical exertion, dietary restriction, and temperature extremes - are transduced via highly conserved molecular networks. These pathways act as biological sensors, detecting transient metabolic deficits and upregulating protective mechanisms that overshoot the baseline, resulting in enhanced stress resistance and extended longevity 8119.
The AMPK, SIRT1, and mTOR Signaling Network
At the core of nutrient and energy sensing lies the complex interplay between AMP-activated protein kinase (AMPK), Sirtuin 1 (SIRT1), and the mechanistic target of rapamycin (mTOR) 101112. The mTOR kinase functions within two structurally and functionally distinct complexes, mTORC1 and mTORC2 1013. mTORC1 operates as a primary anabolic driver, promoting cellular mass growth, ribosomal biogenesis, and protein synthesis in response to nutrient abundance, growth factors, and insulin 81314. However, the nutrient-sensing mTOR pathway also drives cellular aging. Hyperactivation of mTORC1 inhibits macroautophagy and promotes the accumulation of aggregation-prone proteins, cellular senescence, and the acceleration of age-related pathologies 810.
Conversely, energetic stressors activate AMPK by increasing the intracellular AMP-to-ATP and NAD+-to-NADH ratios 101516. Upon activation, AMPK initiates a catabolic shift. It directly inhibits mTORC1 via the phosphorylation of the tuberous sclerosis complex 2 (TSC2) and Raptor 101112. Furthermore, AMPK activates SIRT1, an NAD+-dependent histone deacetylase that enhances DNA repair, cell survival, and mitochondrial function by modulating transcription factors such as NF-κB, FOXO, and PGC-1α 101215. The accumulation of PGC-1α in the nucleus subsequently drives mitochondrial biogenesis and improves oxidative phosphorylation efficiency 1516.
While chronic suppression of mTOR via pharmacological agents is widely recognized to extend lifespan in model organisms, the hormetic principle suggests that periodic, low-dose inhibition interspersed with periods of activation provides superior outcomes compared to total, continuous suppression 1314. Low concentrations of agents targeting this pathway induce partial mTOR inhibition, triggering protective autophagy while preserving essential cellular functions - a balance that highlights the necessity of cyclical stress and recovery in longevity biology 13.
Mitohormesis and the Mitochondrial Unfolded Protein Response
Hormetic stress fundamentally alters organelle dynamics, particularly within mitochondria. Mild oxidative stress or temporary electron transport chain (ETC) impairment initiates a process known as mitohormesis 1721. In this paradigm, transient elevations in reactive oxygen species (ROS) act as vital signaling molecules rather than purely destructive agents 17. This mild mitochondrial perturbation triggers a positive feedback loop involving nuclear factor erythroid 2 - related factor 2 (Nrf2) signaling, which upregulates endogenous antioxidant defenses, including heme oxygenase 1 (HO-1), glutathione peroxidase (GPX), and superoxide dismutase (SOD) 1721.
Simultaneously, mitochondrial stress activates the mitochondrial unfolded protein response (UPRmt). In Caenorhabditis elegans, this response is mediated by the transcription factor ATFS-1, which contains both a mitochondrial targeting sequence and a nuclear localization signal 21181920. Under basal conditions, ATFS-1 is imported into healthy mitochondria and degraded by the Lon protease 20. During mitochondrial stress, import efficiency declines, allowing ATFS-1 to traffic to the nucleus, where it alters gene expression to recover mitochondrial homeostasis by upregulating chaperones like HSP-6 and HSP-60 212021. In mammals, this conserved response is orchestrated by the ortholog ATF5 and the integrated stress response mediator ATF4 212026.
The application of mitohormesis remains strictly bounded by dose constraints. Research demonstrates that while mild UPRmt activation is pro-longevity, constitutive or excessive activation of ATFS-1 paradoxically decreases lifespan 19. This dynamic indicates a critical trade-off between stress resistance and longevity; a hyperactivated stress response becomes detrimental to overall organismal survival, reinforcing the necessity of precise stressor dosage 1922.
Endoplasmic Reticulum Stress and the UPRER Pathway
Similar to mitochondria, the endoplasmic reticulum (ER) manages proteostasis through the unfolded protein response (UPRER), regulated by the PERK, IRE1, and ATF6 signaling branches 21182324. Mild ER stress promotes the dissociation of the binding immunoglobulin protein (BiP) from inositol-requiring enzyme 1 (IRE1), triggering unconventional splicing of X-box binding protein 1 (XBP1) mRNA 21. The resulting XBP1s isoform functions as a transcription factor to upregulate ER chaperones, thereby restoring proteostasis 21.
The UPRER exhibits clear age-dependent hormetic thresholds. Transient, pharmacological ER stress imposed early in development enhances proteostasis, prevents the age-related decline in UPRER efficiency, and extends lifespan via the IRE-1/XBP-1 branch 2325. Furthermore, adaptive UPRER signaling mediated by PERK increases the assembly of mitochondria-ER contact sites (MERCS), facilitating beneficial lipid transfer, calcium signaling, and mitochondrial remodeling 182632.
However, in aged organisms, the UPRER becomes increasingly inefficient 2425. When older subjects experience chronic ER stress, they exhibit an exaggerated, maladaptive metabolic flux signature characterized by a profound suppression of global protein translation, impaired de novo lipogenesis, and a shift toward pro-apoptotic signaling 1824. This divergence underscores that the physiological capacity to harness hormetic stress diminishes with age, making calibrated interventions critical in older populations.
Applications of Hormesis in Physical Exercise
Physical exercise represents a highly potent, pleiotropic hormetic stressor that induces profound muscular, cardiovascular, and neurological adaptations 1633. By altering energy balance and inducing transient hypoxia and mechanical stress, exercise acts as a metabolic challenge that upregulates the AMPK/SIRT1/PGC-1α axis, driving mitochondrial biogenesis and the release of myokines like irisin and brain-derived neurotrophic factor (BDNF) 1633.
Cardiorespiratory Fitness and Mortality Predictors
The systemic adaptation to regular physical stress is most accurately quantified by cardiorespiratory fitness (CRF), often measured as VO2max. Clinical evidence establishes that low CRF is an independent and potentially stronger predictor of mortality than established risk factors such as smoking, hypertension, and type 2 diabetes 3427. Individuals in the highest fitness percentiles carry roughly half the all-cause mortality risk of the least fit, with high CRF also correlating to a 40% lower risk across specific cancer types and reduced incidences of surgical complications and dementia 3427.
Minimum Effective Dose and Dose-Response Plateaus
Extensive epidemiological data demonstrate that the dose-response relationship between physical activity and mortality fundamentally challenges the "more is better" paradigm, strictly following an L-shaped or curvilinear trajectory 282930. A 2024 meta-analysis encompassing over 3.36 million participants confirmed that high volumes of moderate-to-vigorous physical activity (MVPA) reduce mortality risk, but the greatest relative benefits occur at the lowest initial doses 2939.
In a massive prospective cohort study of 109,407 Chinese adults, researchers identified a "minimum effective dose" or "longevity floor" of merely 75 minutes of total physical activity per week, which yields an immediate 11% to 23% reduction in all-cause mortality 28. As exercise volume increases, benefits deepen but eventually reach a point of diminishing returns. This plateau typically occurs around 2,400 minutes per week (approximately 5.7 hours per day of total activity) 28. Beyond this ceiling, the mortality benefit curve flattens completely, and confidence intervals widen, indicating no statistically significant additional survival advantage for extreme endurance volumes 28.
For vigorous-intensity leisure-time activity, the dose-response curve is even steeper. The minimal optimal dose is identified at approximately 60 to 70 minutes per week, with maximal risk reduction plateauing near 200 minutes per week 2939. Exceeding these bounds can push the physiological response into the inhibitory phase of the hormetic curve. Extreme endurance training displays a U-shaped association for specific clinical outcomes, elevating the risk for atrial fibrillation and degenerative musculoskeletal disorders (MSD) due to unmitigated mechanical and oxidative stress 2831.
Vigorous Intermittent Lifestyle Physical Activity
The hormetic benefits of exercise do not strictly require prolonged, structured sessions. Research into Vigorous Intermittent Lifestyle Physical Activity (VILPA) - measured via wrist-worn accelerometers in cohorts such as the NHANES 2011-2014 study - demonstrates that short bursts of vigorous activity lasting up to one minute significantly reduce mortality risk 32. The dose-response for VILPA is similarly non-linear. A median frequency of 5.3 bouts per day is associated with a 44% lower risk of all-cause mortality compared to zero bouts 32. However, the curve flattens significantly beyond 8 to 10 bouts per day, reinforcing the concept that brief, intense stressors trigger maximal adaptive transcription without the need for sustained, exhausting volume 32.
Table 1: Physical Activity Intensity and Mortality Dose-Response Parameters
| Activity Modality | Minimum Effective Dose | Peak Benefit Threshold (Plateau) | Primary Physiological Adaptations |
|---|---|---|---|
| Total Physical Activity (TPA) | 75 mins/week | ~2,400 mins/week | Baseline metabolic regulation, CRP reduction, basic cardiovascular maintenance. |
| Moderate-to-Vigorous (MVPA) | 100 - 150 mins/week | ~340 mins/week | Enhanced CRF (VO2max), mitochondrial biogenesis, sustained insulin sensitivity. |
| Vigorous Intensity | 60 - 70 mins/week | ~200 mins/week | High-level AMPK activation, increased cardiac stroke volume, peak autophagic flux. |
| Vigorous Intermittent (VILPA) | ~3-4 bouts/day | 8 - 10 bouts/day | Acute sympathetic nervous system activation, rapid glucose clearance. |
Nutrient Deprivation and Fasting Modalities
Fasting and caloric restriction (CR) induce systemic hormesis by depriving cells of exogenous energy, forcing the organism to shift from glucose dependence to lipid metabolism and ketosis 11333444. This transition acts as a powerful signaling event that downregulates the mTORC1 growth pathway while simultaneously upregulating macroautophagy, cellular repair, and stress resistance networks 44353647.
Metabolic Shifts and the Induction of Autophagy
Under conditions of nutrient abundance, high mTOR signaling inhibits the initiation of autophagy 3548. However, as fasting depletes hepatic glycogen stores and drastically lowers insulin levels, the rising AMP-to-ATP ratio activates AMPK 443548. Activated AMPK directly phosphorylates and activates the ULK1 kinase complex, overriding mTOR suppression and triggering the formation of autophagosomes 124849.
Autophagy functions as an intracellular recycling program. The autophagosome engulfs misfolded proteins, damaged organelles, and intracellular pathogens, subsequently fusing with a lysosome for enzymatic degradation 443537. The resulting macromolecules are recycled into raw materials for energy production and de novo protein synthesis, ensuring cell survival and contributing to systemic longevity 443637.
Timelines of Fasting-Induced Cellular Repair
The temporal dynamics of human fasting align perfectly with the delayed overcompensation model of hormesis. During the initial 12 to 24 hours of fasting, the body experiences glycogen depletion, a significant drop in insulin, and rising glucagon levels 4838. By 18 to 24 hours, beta-hydroxybutyrate (ketone) levels rise, and the earliest autophagic pathways flicker on in response to the perceived fuel shortage 444837.
Between 24 and 48 hours, systemic autophagy signaling accelerates meaningfully 4837. Clinical studies measuring autophagic flux in human peripheral blood mononuclear cells demonstrate that intermittent nutrient restriction effectively modifies this primary hallmark of biological aging 39. Molecular analyses of subjects undergoing dawn-to-dusk intermittent fasting (e.g., Ramadan cohorts) confirm the significant upregulation of core autophagic genes, including ULK1, ATG5, BECN1, LAMP2, and LC3B, alongside transient elevations in inflammasome markers that later normalize 4940.
Prolonged fasting reaching 48 to 72 hours represents the peak zone of deep cellular repair 483738. During this intense metabolic reset, mTOR suppression is maximal, and deep ketosis creates an optimal environment for cellular cleansing 48. Furthermore, the clearance of old, damaged white blood cells during this period sends a potent regenerative signal to hematopoietic stem cells residing in the bone marrow, prompting a renewal of the immune system upon refeeding 333738.
Limitations and Maladaptive Responses to Extended Fasting
While 72 hours strikes an optimal balance between maximum biological benefit and safety, extending fasting beyond this threshold without medical supervision often bypasses the hormetic ceiling, shifting the body into systemic distress 48. The "more is better" fallacy applied to nutrient deprivation ignores the fundamental biological requirement for periods of growth and synthesis 13.
Excessive, continuous mTOR suppression prevents necessary cellular proliferation, muscle protein synthesis, and tissue repair 13. Extended fasts carry significant risks of severe muscle catabolism, potentially fatal electrolyte imbalances, dizziness, and disrupted blood glucose regulation 5455. Furthermore, extreme dietary restriction in aging populations, where basal autophagic capacity is already diminished, can exacerbate nutrient deficiencies rather than promote rejuvenation 54. Consequently, cyclical feeding - combining the targeted stress of fasting with the restorative anabolic phase of nutrient reintroduction - is an absolute biological requirement for harnessing the longevity benefits of autophagy 13.
Thermal Stress and Cold Water Immersion
Brief exposure to extreme environmental cold serves as a potent neuroendocrine and metabolic stressor that, when dosed correctly, triggers systemic adaptive resilience 414243. Deliberate cold exposure, documented as a therapeutic practice since antiquity in texts like the Edwin Smith Papyrus, relies on the rapid induction of physiological shock to force adaptation 43.
Thermoregulation and TRPM8 Receptor Activation
Cold water immersion (CWI) rapidly decreases skin temperature, triggering the activation of transient receptor potential cation channel melastatin 8 (TRPM8) receptors located in the neurons of peripheral ganglia, including the dorsal root and trigeminal ganglia 4142. TRPM8 receptors are highly sensitive to temperatures between 10°C and 28°C and govern calcium ion transmembrane transport, serving as the primary molecular determinants of cold sensitivity and thermotransduction 4142.
The activation of these somatosensory receptors sends immediate signals to the hypothalamus, initiating a massive sympathetic nervous system (SNS) response 42. This response is characterized by systemic peripheral vasoconstriction, shivering, and the acute release of norepinephrine and cortisol from the adrenal glands 4142. Concurrently, repeated cold exposure upregulates brown adipose tissue (BAT) thermogenesis, increasing cellular respiration and mitochondrial uncoupling to generate heat 414344.
Neurological and Psychological Adaptations
The hormetic benefits of cold stress extend beyond metabolic thermoregulation into profound neuroplasticity and mood modulation. Functional magnetic resonance imaging (fMRI) studies demonstrate that brief CWI significantly increases neural interaction across large-scale brain circuits involving multiple limbic structures, specifically the medial and left rostral prefrontal cortices, the left anterior insula, and the anterior cingulate cortex 41. This sudden neural activation facilitates positive affect, reduces nervousness and distress, and results in heightened alertness and motivation driven by acute dopamine and noradrenaline release 414260.
Toxicological Thresholds of Thermal Stress
The boundary between beneficial thermal hormesis and pathological stress is exceptionally narrow. The adaptive response requires the cold stress to be brief and strictly controlled (typically 10 - 15°C water for 1 to 5 minutes) 414243. Extending the duration or excessively lowering the temperature bypasses the hormetic threshold, leading to the primary danger of accidental hypothermia, defined as a drop in core body temperature below 35°C 41.
Even prior to the onset of clinical hypothermia, high-dose cold stress produces deleterious neurological effects. Severe cold water immersion can cause cognitive impairments - specifically reducing attention spans, processing speeds, and executive function - due to the slowing of neuronal conduction velocity and alterations in synaptic transmission mechanisms 4142. This rapid transition from enhanced alertness to cognitive impairment vividly illustrates the biphasic nature of the hormetic curve.
Table 2: Comparative Thresholds and Mediators of Hormetic Interventions
| Intervention Modality | Primary Stressor Mechanism | Key Molecular Mediators | Minimum Effective Dose (Floor) | Diminishing Returns / Toxicity (Ceiling) |
|---|---|---|---|---|
| Physical Exercise | ATP depletion, hypoxia, mechanical muscle stress | AMPK, PGC-1α, Irisin, BDNF, mTOR (cyclical) | 75 mins/week of moderate-to-vigorous activity | >2,400 mins/week (plateau); high intense volume increases AFib risk |
| Fasting / Dietary Restriction | Nutrient deprivation, hepatic glycogen depletion | mTORC1 inhibition, SIRT1, UPRmt, Autophagy (ULK1, ATG5) | 16 - 24 hours (initiates early autophagy pathways) | >72 hours (muscle catabolism, electrolyte imbalance, immune suppression) |
| Cold Exposure | Thermal shock, peripheral vasoconstriction | TRPM8, Norepinephrine, PGC-1α, BAT thermogenesis | Brief immersions (10 - 15°C for ~1 - 5 minutes) | Prolonged exposure induces hypothermia, oxidative damage, and cognitive decline |
Pharmacological and Phytochemical Modulators of Hormesis
In addition to behavioral stressors, the longevity network can be engaged through pharmacological agents and plant-derived phytochemicals that act as biological mimetics of caloric restriction and physiological stress 111.
Rapamycin and mTOR Modulation
Rapamycin, an antifungal antibiotic produced by soil bacteria, is currently one of the most robust pharmacological agents known to extend lifespan across multiple model organisms 81314. It functions by directly inhibiting mTORC1, thereby mimicking the cellular state of nutrient deprivation 14. However, rapamycin displays strict hormetic properties. At high, immunosuppressive doses, the drug is toxic to cells and impairs vital metabolic processes 13. Conversely, low-dose rapamycin induces incomplete mTOR inhibition, which allows for the upregulation of protective autophagy, rejuvenation of hematopoietic stem cells, and improved immune responses without triggering catastrophic cellular shutdown 111314.
Phytochemicals and the SIRT1-PGC-1α Axis
A diverse array of phytochemicals - including resveratrol, curcumin, spermidine, and specific flavonoids - exert anti-aging effects by modulating the same nutrient-sensing pathways activated by physical exercise and fasting 11516. Resveratrol, a polyphenol found in grapes, functions as a direct activator of the SIRT1/AMPK pathway 1517. By upregulating AMPK, resveratrol increases the NAD+-to-NADH ratio, further stimulating SIRT1 deacetylase activity, which in turn promotes the nuclear accumulation of PGC-1α 1517. This molecular cascade drives mitochondrial biogenesis, attenuates ROS accumulation, and preserves mitochondrial respiratory capacity in aging cells 1517.
Spermidine, a natural polyamine, promotes autophagy via the epigenetic regulation of ATG genes and has demonstrated the ability to increase critical autophagic proteins like Beclin-1 (BECN1) and ULK1 in human trials 11. Like all hormetic agents, the efficacy of these compounds is dose-dependent, acting to restore age-related suppressed autophagic flux at low concentrations while potentially inducing excessive autophagy-induced cell death at supraphysiological doses 1516.
Proteomic Clocks and Population-Level Aging Variations
The mechanisms of hormesis do not operate in a vacuum; their long-term efficacy ultimately dictates the divergence between an individual's chronological age and their true biological age. Advances in precision geromedicine have begun to quantify the systemic impact of lifestyle stressors using high-resolution biological clocks and large-scale population cohorts.
Organ-Specific Proteomic Aging Clocks
Recent developments utilizing plasma proteomics and machine learning have allowed researchers to construct organismal and organ-specific aging clocks that track the physiological toll of aging with unprecedented accuracy 4546. Studies analyzing diverse longitudinal cohorts, including the UK Biobank (n = 43,616), the China Kadoorie Biobank (n = 3,977), and United States cohorts (n = 800), demonstrate that accelerated biological aging of specific organs predicts disease onset, progression, and mortality far more accurately than chronological age or standard clinical risk factors 4546.
Brain aging, evaluated through multi-organ proteome-based biological age gaps (ProtBAGs), exhibits the strongest link to all-cause mortality 4546. Accelerated brain aging is deeply influenced by environmental determinants, modifiable lifestyle factors, and specific genetic markers such as the GABBR1 and ECM1 genes 4546. Remarkably, a "super-youthful" proteomic brain profile - often maintained through the successful, chronic application of hormetic stressors like exercise and metabolic regulation - appears to confer resilience even against severe genetic risk factors, such as the APOE4 haplotype associated with Alzheimer's disease 4546.
Genomic Studies in Long-Lived Populations
To separate the genetics of exceptional longevity from the benefits of environmental hormesis, researchers have conducted genome-wide association studies (GWAS) on specifically long-lived demographic pockets. For example, a GWAS analyzing longevity in 705 individuals from the Calabrian population in Italy identified distinct population-specific genetic effects 63. Gene-based tests highlighted biological processes central to aging, including proteostasis, DNA repair, telomere maintenance, and apoptosis 63. Notably, established longevity loci such as APOE and FOXO3 were absent in this specific cohort, underscoring that human lifespan extension arises from a complex combination of unique genetic influences and conserved, modifiable aging-related mechanisms 63.
Similar genomic studies of long-living adults (aged ≥90 years) indicate that healthy longevity is positively associated with variants within the MYO18B and TBC1D28 genes, while variants in APOE and APOC1 correlate with an increased risk of age-related diseases 47. However, the presence of disease-associated variants does not definitively preclude longevity, reinforcing the premise that behavioral hormesis and lifestyle interventions can significantly modulate genetic predispositions 47.
Global Perspectives on Healthspan and Precision Geromedicine
Despite the molecular clarity surrounding longevity biology, widespread public implementation remains a formidable challenge. Initiatives like the HEalthy LOngevity (HELO) consortium are actively investigating global populations to assess consumer preparedness for precision geromedicine 654849. A landmark HELO survey of over 3,000 residents in Singapore highlighted a profound gap in public health literacy. While 82% of respondents correctly understood "lifespan" as total years lived, only 43% were familiar with the concept of "healthspan" - the duration of years lived free of physical limitations and age-related chronic diseases 4868. Furthermore, fewer than 39% of respondents recognized that biological age is an active, mutable metric that can be lowered 68.
Simultaneously, massive initiatives like the Precision-CARRS cohort (encompassing over 20,000 South Asian adults) are tracking midlife cognitive impairment, subclinical vascular disease, and plasma biomarkers to advance the early detection of Alzheimer's and related dementias 50515253. Together, these global cohorts reveal that while public awareness of the exact molecular mechanisms of aging is low, there is overwhelming consumer interest (over 55% in the HELO survey) in longevity clinics that utilize biomarker tracking, gerodiagnostics, and personalized hormetic interventions to optimize healthy aging 4868.
Conclusion
The biological principle of hormesis fundamentally dismantles the linear assumption that environmental stressors are universally damaging, or conversely, that higher doses of a beneficial intervention will yield proportionally greater physiological returns. At the cellular level, the intricate orchestration of mTOR inhibition, AMPK activation, and the precise regulation of the unfolded protein responses (UPRmt and UPRER) relies entirely on intermittent, low-dose challenges to maintain proteostasis and mitochondrial vitality.
When translated to human behavior, clinical interventions such as physical exercise, dietary fasting, and cold water immersion are strictly governed by these biphasic dose-response curves. The minimum effective dose of physical activity or nutrient deprivation profoundly extends healthspan by initiating powerful autophagic and metabolic repair mechanisms. However, excessive application unequivocally breaches the hormetic ceiling, leading to overtraining syndromes, maladaptive endoplasmic reticulum stress, severe electrolyte imbalances, and accelerated cellular decline. Therefore, the optimization of human longevity and the mitigation of age-related disease fundamentally depend on the strategic, personalized calibration of stress and recovery, utilizing precise, moderate perturbations to continuously expand the organism's adaptive resilience.