Applications and limits of Shannon information theory in biology
Foundations of Information Theory in Biology
Information theory, formally established by Claude Shannon in 1948 through his seminal publication A Mathematical Theory of Communication, was initially designed to address the engineering limits of data compression and the reliable transmission of signals over noisy telecommunication channels 123. Progressing beyond the foundational work of Harry Nyquist, Ralph Hartley, Alan Turing, and Norbert Wiener, Shannon's framework provided rigorous mathematical definitions for quantifying uncertainty 34. The central metric of this framework, information entropy, measures the statistical uncertainty associated with a random variable and allows engineers to define the absolute capacity of both lossless and lossy communication channels 123.
The application of Shannon's mathematics rapidly transcended the boundaries of electrical engineering. During the 1950s and 1960s, parallel to the discovery of the structure of DNA and the formulation of the central dogma of molecular biology, researchers recognized profound structural analogies between the sequential encoding of genetic material and the digital encoding of binary data 35. Driven by the early optimism of cybernetics and the "new movement" championed by Henry Quastler in 1953, biologists began mapping the variables of living phenomena - such as nucleic acid sequences, protein folding structures, and metabolic regulatory networks - into the framework of information theory 36. This cross-disciplinary translation provided the mathematical foundation for modern computational biology, enabling researchers to quantify the order and complexity inherent in living organisms 14.
Despite the mathematical robustness of these early applications, treating biological systems strictly as information channels introduces profound theoretical complications. Shannon explicitly established that his theory was concerned exclusively with the syntactic properties of data - whether bits are transmitted accurately - and was entirely indifferent to the semantic meaning or functional utility of a message 2778. This fundamental limitation has triggered an enduring theoretical debate regarding the precise boundaries of the information metaphor in biology. The core scientific challenge lies in distinguishing between the objective measurement of statistical correlations within biochemical networks and the normative, functional ways in which living systems actively exploit that information to survive, adapt, and construct their environments 91110.
DNA and Gene Expression as a Communication Channel
The application of Shannon information theory to molecular biology conventionally begins with the conceptualization of the genome as a highly stable, digital data repository. Within this paradigm, evolutionary adaptation and molecular replication are viewed as mechanisms of information transfer, where the preservation of genetic fidelity against environmental noise is a primary biological imperative 511.
The Genetic Code and Error Minimization
The central dogma of molecular biology - the transcription of DNA into messenger RNA (mRNA), followed by the translation of mRNA into functional amino acid sequences - can be modeled mathematically as a noisy communication channel 1215.
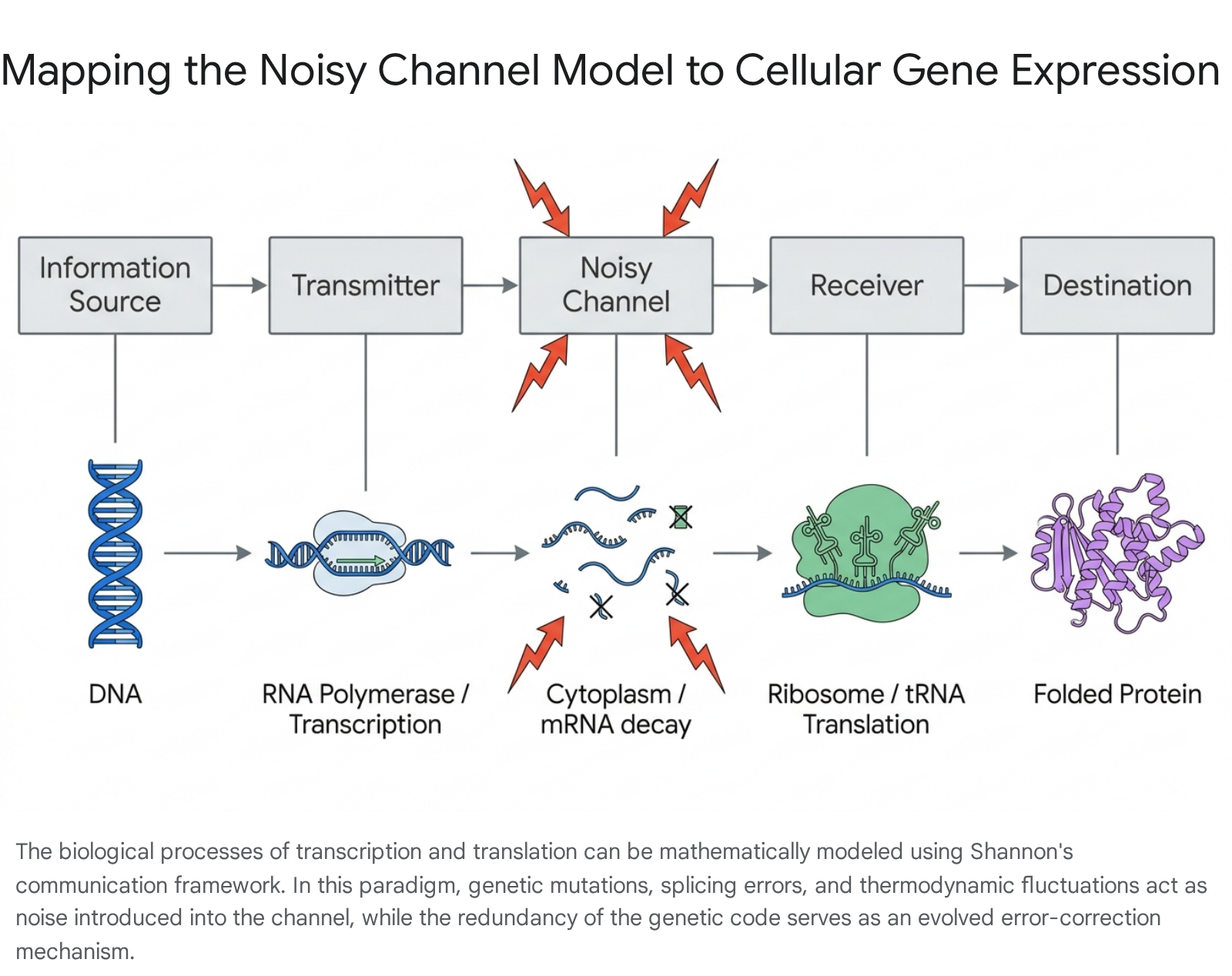
The genetic code maps sixty-four possible nucleotide triplets, or codons, to twenty fundamental amino acids. While early biological hypotheses suggested this specific, redundant mapping might be a "frozen accident," information-theoretic models demonstrate that the code likely emerged through evolutionary optimization to minimize the impact of translation errors and genetic mutations 1215.
Under this framework, the stochastic mapping of codons to amino acids is subjected to rate-distortion theory. Organisms compete based on the fitness of their translation codes, and mathematical models indicate that a stable genetic code emerges at a supercritical phase transition within the noisy channel, moving the system from a random, non-coding state to a structured, coding one 1215. The topology of this genetic "error-graph" - in which codons are connected if their physical or chemical similarities make them likely to be confused by the cellular translation machinery - imposes strict mathematical limits on the upper bound of possible amino acids 1215. This topological limit is conceptually related to the classical map-coloring problem in mathematics. Consequently, the redundancy of the genetic code functions as an evolved error-correction mechanism, strictly analogous to the parity bits utilized in digital telecommunications to ensure signal integrity across noisy transmission lines 512.
Transcriptional Dynamics and Noise
Information theory is heavily utilized to dissect the parameters of transcriptional dynamics, which exhibit substantial variability even under highly controlled conditions. The synthesis of mRNA involves complex, multi-state promoter cycles, elongation phases, and co-transcriptional splicing events 16. These biochemical reactions depend on the interactions of molecules present in very small numbers within the cell. Consequently, the inherent stochasticity of molecular binding and intracellular diffusion generates significant noise along the cascade that leads from DNA to the synthesis of a folded protein 13.
To quantify this multimodal genomic data, researchers utilize Transcriptional Information Maps (TIMs) that measure the flux of transcriptional information between localized genetic variants, such as Single Nucleotide Polymorphisms (SNPs), and continuous downstream gene expression levels 14. In these models, a channel in the transcriptional mapping indicates a regulatory mechanism, and the mutual information between the two linked nodes evaluates the degree of their dependence, effectively isolating regulatory signals from statistical microarray noise 14. Analyzing these maps clusters SNPs and genes into specific causal groups, demonstrating how genetic architecture channels information flow.
Evolutionary Constraints on Transcriptional Noise
The advent of single-cell transcriptomics has demonstrated that stochastic gene expression (SGE) causes isogenic cell populations to display wide phenotypic variability, even when existing in entirely homogeneous environments 13. Genome-wide assessments indicate that this transcriptional noise is not merely a physical limitation but is actively shaped by evolutionary constraints. Noise levels in mRNA distributions correlate significantly with three-dimensional nuclear domain organization, gene age, and the precise position of the encoded protein within a broader biological pathway 13.
Because transcriptional noise propagates through gene networks, it acts as an important component of the organism's overall phenotype. Rather than being universally suppressed, the variance of expression itself serves as a target of adaptation 13. Evolutionary simulations of regulatory channels reveal that identical steady-state protein levels can arise from distinct parameter genotypes, and small network mutations allow bacterial populations to explore vast regions of functional space 15. By maintaining specific levels of expression variance, biological systems operate near their theoretical channel capacity, preserving the phenotypic plasticity required to adapt to rapidly fluctuating external environments 131520.
Cellular Computation and Network Inference
While early applications of information theory focused predominantly on the static storage capacity of genomes, contemporary systems biology treats the living cell as a dynamic, real-time computational entity 1617. Cells do not passively warehouse DNA; they actively process incoming information to respond to chemical gradients, mechanical forces, and adjacent paracrine signaling.
Mutual Information in Molecular Networks
The calculation of mutual information and channel capacities within living cells has substantially advanced the reverse-engineering of signal transduction cascades. Algorithms based on information theory, such as ARACNE and CLR, are routinely deployed to infer the structural topology of complex biological networks by determining the mutual information shared between molecular nodes 18.
However, measuring information transmission within cellular systems presents unique constraints. In many biochemical networks, the average channel capacity at the single-cell level approaches roughly one to two bits, indicating that an individual cell can reliably distinguish between only a few distinct states of an external stimulus, such as the complete absence or high concentration of a specific ligand 18. Furthermore, accurately estimating the probability distribution functions required for Shannon's metrics in high-dimensional omics data requires exceptionally large sample sizes 1618. When analyzing time-series data, the computational burden scales exponentially, and mutual information metrics alone cannot resolve the directionality of causation without supplementary perturbation experiments 18.
Multicellular Information Processing
Despite the relatively low channel capacity of single cells, biological systems overcome this limitation through distributed computation. Complex signaling modalities operate collectively across spatial and temporal dimensions. For example, information theory metrics applied to time-series datasets of Xenopus laevis embryonic stem cells reveal intricate patterns of information flow concerning endogenous calcium and cytoskeletal actin dynamics 19. By mapping active information storage and transfer entropy between minimally manipulated tissue explants, researchers quantify how cells collectively integrate external cues 19. This distributed network approach demonstrates that the reliable transmission of developmental signals relies on multicellular collectivity, mirroring the architecture of parallel computing systems.
Synthetic Biology and Engineered Biological Circuits
The conceptualization of cells as programmable information processors has given rise to the field of synthetic biology, where the principles of electrical engineering, control theory, and information theory are directly applied to create artificial biological circuits 172021. Researchers design genetic regulatory modules capable of executing logic operations tailored for specific biopharmaceutical or agricultural outcomes.
Programming Logic Gates in Cellular Systems
Synthetic gene circuits function by integrating multiple customizable input signals - such as small molecules, hormones, or external light - through processing units constructed from biological parts, ultimately producing a predictable output 2022. By modularly combining specialized promoters, repressor proteins, and engineered DNA-binding domains, scientists have successfully implemented complex Boolean logic within both mammalian and plant systems. Using recombinases and specialized control elements, researchers have activated transgenes corresponding to YES, OR, and AND logic gates, and repressed them using NOT, NOR, and NAND gates 2122.
Through genetic recombination, these synthetic circuits can create stable, long-term changes in gene expression, effectively acting as biological memory units that record past environmental stimuli 22. In mammalian cells, advanced systems leverage feedforward control loops and promoter editing mechanisms to fine-tune transcription factor levels, allowing researchers to accurately dial the expression of therapeutic synthetic genes up or down 2023.
High-Throughput Antigen Discovery
The capacity to engineer cellular computation is heavily utilized in advanced therapeutics. Technologies such as TCR-MAP (T Cell Receptor Mapping of Antigenic Peptides) utilize synthetic receptor-stimulated circuits within immortalized T cells 2430. This circuit activates the sortase-mediated tagging of engineered antigen-presenting cells expressing specific peptides on major histocompatibility complexes (MHCs). The synthetic circuit allows researchers to query T cell receptors with unknown specificities against massive, barcoded peptide libraries in a high-throughput, pooled screening context 2430. By functioning as a targeted information retrieval system, these synthetic circuits accelerate antigen discovery for complex diseases, including cancer and autoimmunity 2430.
Biocontainment and Security in Synthetic Biology
As synthetic biology develops increasingly sophisticated information processing capabilities, researchers have raised fundamental security and biocontainment concerns. Extreme bioengineering initiatives, such as the creation of "mirror life" - organisms built entirely from left-handed proteins and right-handed DNA - demonstrate the ultimate extent of cellular reprogramming 31. Because all known biological processes are strictly dependent on molecular chirality, a mirrored organism would operate on an information architecture entirely invisible to natural immune systems, predators, and degradation pathways 31. This highlights a severe consequence of altering the fundamental information substrate of biology: natural systems lack the correlational history required to interpret or neutralize artificially engineered biological code, rendering perfect biocontainment practically impossible 31.
Thermodynamic Boundaries and Active Matter
To fully bridge the mathematical abstraction of information theory with the physical reality of biology, researchers investigate the energetic and thermodynamic costs of cellular computation. Living systems are fundamentally defined by their status as active matter - nonequilibrium many-body systems in which individual components continuously consume free energy to sustain autonomous motion, structural self-organization, and persistent information processing 3225.
Dissipative Adaptation
The physical mechanism linking energy flow to the emergence of biological computation is formalized through the theory of dissipative adaptation, pioneered by biophysicist Jeremy England 34262728. Under the laws of non-equilibrium statistical mechanics, classical equilibrium principles such as detailed balance and time-reversal symmetry are invalidated 32. When a system of interacting particles is driven by an external energy source (such as chemical fuel or solar radiation) and surrounded by a heat bath, it will spontaneously restructure itself into configurations that maximize the dissipation of energy 262728.
This framework implies that the emergence of complex, self-replicating molecular structures - the precursors to biological life - is not a statistical anomaly but a highly probable thermodynamic outcome 3426. Living organisms are exceptionally efficient at capturing energy and routing it through complex metabolic pathways. In this thermodynamic view, Darwinian evolution by natural selection is recontextualized as a specialized macro-biological instance of a universal physical principle: matter spontaneously adapts to its energetic environment to foster the incessant dispersal of energy and increase the overall entropy of the universe 342628.
Non-Equilibrium Dynamics in Complex Environments
Active matter encompasses a broad spectrum of phenomena, ranging from nanomotors and protein filaments to bacterial swarms and multicellular tissues 322529. The collective dynamics of these systems often exhibit emergent behaviors, such as motility-induced phase separation, hydrodynamic bound states, and synchronized chemotaxis 29. By driving molecular components out of equilibrium, active matter avoids the inherent limitations of isolated physical systems, executing spatiotemporal patterns that enable macro-scale biological functions 25. Current interdisciplinary research efforts map how these systems interact within geometrically confined, complex fluid environments, combining active elasticity and fluid mechanics to understand the autonomous processing capabilities of microbial habitats 25.
The Information Processing Threshold
While dissipative adaptation provides a robust physical mechanism for self-organization, it encounters strict definitional boundaries when attempting to account for the uniquely computational nature of life. Critics point out that numerous inanimate, non-equilibrium systems fall under the umbrella of dissipative structures. For instance, turbulent vortices or Jupiter's Great Red Spot are highly dissipative, non-equilibrium structures that have maintained stable organization for centuries 2628. Yet, these systems are not classified as living.
The distinction resides in explicit information-processing capacity 28. Living active matter does not merely channel heat; it actively utilizes molecular sensors to gather environmental information, stores this data within algorithmic polymer sequences, and executes programmed, functional responses that insulate the organism from entropic decay 3228. Therefore, while thermodynamic dissipation is a necessary prerequisite for the origin of structured order, the emergence of a semantically closed information-energy loop - where the system's material operations are regulated by its own interpreted symbols - is required to define a complex system as biologically alive 30.
The Semantic Information Problem
The crux of the theoretical friction regarding the use of information theory in biology is the distinction between syntactic information and semantic information 91131. Shannon's foundational theory deliberately ignores meaning. From a strict information-theoretic perspective, a random, nonsensical sequence of nucleic acids can possess the exact same entropy as a highly conserved, functional gene essential for survival 267.
Syntactic Correlation Versus Biological Function
Shannon information is strictly correlational, symmetric, and ubiquitous. If physical variable A correlates with variable B, they carry Shannon information about one another, regardless of whether any biological machinery utilizes this correlation 1110. Under this definition, almost any physical system - from tree rings to weather patterns - carries massive amounts of information.
Semantic information, conversely, is normative, asymmetric, and functional. It possesses a specific "direction of fit" to its environment 111032. A biological signal, such as an animal alarm call or a cellular transcription factor, carries semantic information because it is teleologically "supposed" to elicit a specific biological response based on a history of natural selection 1032. Crucially, semantic information possesses the capacity for misrepresentation or error if that response fails, a feature entirely absent from pure statistical correlations 111032.
To systematically distinguish between these definitions and clarify the ongoing debate in theoretical biology, the table below compares the primary interpretations of information:
| Feature | Shannon (Syntactic) Information | Algorithmic (Kolmogorov) Complexity | Semantic (Functional) Information |
|---|---|---|---|
| Core Definition | Measures the reduction of statistical uncertainty between random variables 231. | Measures the length of the shortest possible computer program required to generate a specific sequence 113133. | Measures the subset of syntactic information that causally contributes to a system's viability or intrinsic goal 934. |
| Primary Focus | Data transmission limits, compression ratios, error rates, and channel capacity 123. | Mathematical compressibility, sequential patterns, and absolute structural randomness 1133. | Biological utility, contextual meaning, and normative correctness regarding survival 91134. |
| Directional Symmetry | Symmetric: Statistical correlation is inherently bidirectional 11. | Asymmetric: Flows strictly from the generating algorithm to the final output sequence 11. | Asymmetric: Flows from an environmental source to an interpreting, functional receiver 11. |
| Capacity for Error | None: There are no "false" correlations, only observed probability distributions 11. | None: Not applicable to concepts of truth, falsity, or biological correctness. | High: Capable of misrepresentation, malfunction, and biological misfiring 1032. |
| Biological Example | Quantifying the absolute entropy (in bits) of a DNA binding site sequence 320. | Assessing the structural complexity required to perfectly describe a folded protein chain 46. | A genetic sequence successfully encoding a protein necessary to neutralize a specific pathogen 932. |
Mathematical Formulations of Semantic Information
Recognizing that biological agents require a formal measure for meaning, theorists have sought to mathematize semantic information 473536. A prominent model introduced by Kolchinsky and Wolpert mathematically defines semantic information in direct relation to a system's viability function - the quantitative requirement for a system to maintain its existence within a specific environment over time 934.
Within this framework, researchers differentiate between two distinct phases of information. Stored semantic information refers to the information exchanged between a biological agent and its environment within its initial distribution state 34. In contrast, observed semantic information denotes the syntactic information that is continuously and dynamically acquired by an autonomous agent during environmental interaction, which causally prevents the decay of the agent's viability 34.
This distinction has profound implications for synthetic biology. In recent experiments involving smart drug delivery via synthetic cells (SCs), researchers modeled SCs interacting with cancerous cells. The SCs sensed signal molecules released by the cancer cells and subsequently produced a cytotoxic drug 34. By mapping the maximum degree of environment randomization that did not decrease the SC's viability, researchers objectively quantified the observed semantic information in the scenario at precisely 3.91 bits 34. This demonstrates that by using counter-factual intervened distributions - selectively scrambling syntactic information to observe subsequent viability drops - researchers can objectively quantify exactly which bits of data are biologically "meaningful" to an organism's survival 934.
Generalized Semantic Information Theory
Further expanding on this, Generalized Semantic Information Theory (G Theory) attempts to supplant the subjective distortion metrics used in classical Shannon communication models 3536. G Theory replaces the standard distortion constraint with a semantic constraint, utilizing a set of truth functions as a semantic channel 3536. Under this criterion, maximum semantic information is mathematically equivalent to the maximum likelihood criterion 35. From a statistical physics perspective, if Shannon information is analogous to raw free energy, semantic information represents free energy within local equilibrium systems, effectively measuring the efficiency of that energy in performing necessary biological work 35.
Epistemological Limits and Developmental Systems Theory
While information-theoretic formalisms yield undeniably powerful quantitative tools, treating biological entities purely as hardware executing digital software risks profound epistemological errors. The uncritical adoption of engineering terms like "code," "program," and "instruction" has drawn fierce criticism from evolutionary biologists and philosophers, most notably formalized through the framework of Developmental Systems Theory (DST) 503738.
The Parity Thesis and Genetic Determinism
Theorists such as Richard Lewontin, Susan Oyama, and Paul Griffiths argue forcefully that the concept of genetic information often functions as a "metaphor that masquerades as a theoretical concept," which routinely leads to a distorted, deterministic view of molecular biology 75037. The core critique leveled by DST is encapsulated in the parity thesis. The parity thesis argues that there is no justifiable, naturalistic reason to assign a unique, privileged causal role to DNA while relegating all other developmental and environmental factors to the status of mere background material or passive channel noise 71137.
If information in biology is defined strictly by statistical correlation (Shannon's sense), then non-genetic environmental variables - such as incubation temperature, DNA methylation patterns, cytoplasmic gradients, and organelle structures - carry just as much objective information about the resulting adult phenotype as the nucleotide sequence itself 71137. The genome does not contain an isolated, executable computer program; rather, biological development is a massively contingent, epigenetic process where the operative "information" is actively constructed in real-time by the intersection of the genome and the highly specific cellular environment 5053.
Critiques of Preformationism and the Program Metaphor
Lewontin and Oyama point out that treating genes as unilateral "instructions" quietly resurrects an Aristotelian or preformationist view of biology, wherein the adult organism's form is presumed to be already fully represented - albeit translated into a microscopic code - within the zygote 5038. This metaphor encourages an extreme form of biological determinism and creates a false dichotomy between "nature" (viewed erroneously as active, instructive information) and "nurture" (viewed as passive, malleable structural support) 73739.
When computational researchers confuse the measurement of Shannon entropy with the existence of an autonomous genetic program, they bypass the fundamental biomechanical reality of how non-coded physical chemistry actually generates functional coding relations and living organisms 753.
Niche Construction and the Rejection of the Adaptive Landscape
Lewontin similarly criticized the pervasive metaphor of the "adaptive landscape," which visualizes evolving organisms as passive objects climbing static fitness peaks through the external force of natural selection 53. In physical reality, landscapes are not static. Through the process of niche construction, living organisms continually alter their own environments, effectively reshaping the adaptive landscape in real-time 53.
Therefore, information theory is indispensable for mapping statistical correlations and estimating the theoretical limits on network processing capacity 1819. However, it is inherently incapable of substituting for the physical, causal, and deeply contextual explanations required to fully understand biological development, trait inheritance, and phenotypic plasticity 4719.
Algorithmic Complexity and Future Theoretical Frameworks
To bypass the limitations of both Shannon's syntactic metrics and the deterministic program metaphor, some researchers explore evolutionary dynamics through the lens of algorithmic information theory. Developed mathematically by Andrey Kolmogorov and Gregory Chaitin, algorithmic complexity measures information not by probability distributions, but by the computational length of the shortest program required to generate a specific structural output 113133.
Kolmogorov Complexity in Evolutionary Dynamics
While traditional population genetics relies on statistical models of gene frequency, it struggles to account for the origin of life or the sudden emergence of entirely novel genetic structures. Researchers like Christoph Adami apply algorithmic information concepts to re-imagine living things as self-perpetuating information strings interacting within a thermodynamic environment 113340. By framing biological life as information that actively maintains itself against entropic decay, researchers aim to quantify the precise mutational biases and computational creativity of evolutionary systems 1133. Studies utilizing genetic programming methods suggest that the complexity of evolutionary output can be mathematically bounded by the Kolmogorov complexity of the original ancestral state 33. However, critics note that isolating DNA as a four-letter digital string strips away the indispensable context of the cell, the organism, and the ecosystem, inherently limiting the predictive power of pure algorithmic models 33.
Global Institutional Initiatives
The integration of theoretical physics, information theory, and biological computation continues to drive major institutional research globally. At the Max Planck Institute for the Physics of Complex Systems (MPIPKS) in Dresden, dedicated research groups in biological physics model cooperative behaviors across scales, utilizing non-equilibrium statistical mechanics to decipher the self-organization of multicellular systems and active matter 414243. Concurrently, the RIKEN Center for Biosystems Dynamics Research (BDR) in Japan focuses on the multilayered biological processes spanning the entire life cycle, leveraging multiscale simulations, foundation models, and synthetic cellular communication systems to redesign organ functions and trace the physical boundaries of living systems 44454647. These multidisciplinary approaches reflect a unified recognition: advancing the physical understanding of life requires synthesizing the rigorous mathematics of information theory with the fluid, context-dependent reality of biophysics.
Conclusion
Shannon information theory provides a rigorously defined, model-agnostic mathematics that has profoundly shaped the foundations of computational biology. By conceptualizing the central dogma of DNA transcription and translation as a noisy communication channel, researchers can elucidate how evolutionary pressures optimize error-correction and stabilize living systems against constant thermodynamic noise. At the cellular level, the application of mutual information and channel capacity metrics permits the quantitative reverse-engineering of highly complex signal transduction pathways. This paradigm has empowered the design of sophisticated synthetic gene circuits, allowing researchers to program boolean logic directly into mammalian and plant cells for advanced therapeutic and diagnostic applications.
However, the definitive limits of the information metaphor in biology emerge precisely at the boundary between syntax and semantics. Shannon's metrics impeccably measure the probabilities, complexities, and transmission rates of physical states, yet they are entirely blind to biological meaning, purpose, and evolutionary function. Efforts to mathematically formalize semantic information - tying statistical correlation directly to an organism's thermodynamic viability and environmental survival - represent the current frontier in understanding how matter transitions from merely dissipating heat to actively processing knowledge.
Ultimately, while the engineering language of codes, algorithms, and programs offers a highly potent heuristic, the physical reality of living systems is vastly more entangled. As articulated by Developmental Systems Theory, the genome is not an isolated, autonomous architectural blueprint, but rather one of many highly codependent physical factors operating within a dynamic developmental matrix. Treating living organisms strictly as digital information processors remains a highly useful abstraction for specific synthetic and systems-level modeling, but it is an abstraction that must consistently be grounded in the causal, highly contingent, and non-equilibrium reality of biophysics.