Neural and psychological basis of involuntary memory vocalizations
Introduction
Involuntary autobiographical memories (IAMs) refer to the spontaneous, unprompted recollection of specific, personal past events that emerge into consciousness without preceding deliberate retrieval efforts 123. While the phenomenon of spontaneous recall has historically been a challenging subject for empirical study - famously dismissed by early experimental psychologists such as Hermann Ebbinghaus and George Miller as being antithetical to controlled scientific inquiry due to its unpredictable nature 2 - contemporary cognitive science recognizes IAMs as a fundamental and ubiquitous mode of human memory. In daily life, naturalistic studies demonstrate that IAMs occur at least twice as frequently as voluntary, deliberately retrieved autobiographical memories 13.
A specific and highly prevalent subset of these involuntary memories involves the sudden intrusion of past events characterized by intense social pain, embarrassment, or shame. In colloquial terms, these intrusions are often described as "cringe attacks" 45. Accompanying the recall of these acutely distressing memories is a distinct behavioral phenomenon: the spontaneous emission of involuntary vocalizations (e.g., gasping, humming, uttering taboo words, or groaning) and abrupt motor actions (e.g., wincing, shuddering, or facial grimacing) 46.
The neurological and psychological basis for these spontaneous vocalizations resides at the intersection of memory retrieval networks, emotional pain processing centers, and motor execution pathways. The prevailing neurobiological model suggests that the intense affective payload of the retrieved memory generates a transient state of autonomic hyperarousal. This sudden emotional surge, processed primarily within the dorsal anterior cingulate cortex (dACC) and anterior insula (AI), bypasses prefrontal inhibitory controls and "leaks" into adjacent motor cortices 567. Furthermore, from an evolutionary perspective, these vocal bursts share anatomical and functional continuity with primitive mammalian separation-distress vocalizations 89. This report details the mechanisms underlying IAM vocalizations, integrating structural neuroanatomy, evolutionary neurobiology, psychological emotion-regulation frameworks, and cross-cultural phonetic data.
Mechanisms of Involuntary Autobiographical Memories
Cue-Dependent Retrieval and Feature Matching
The activation of an involuntary autobiographical memory is fundamentally governed by cue-dependent retrieval within an associative, context-sensitive cognitive framework 3. Voluntary memory retrieval relies on top-down, executive control mechanisms - heavily mediated by the dorsolateral prefrontal cortex - to execute a gist-based search for past events 1. Conversely, the retrieval of an IAM is a bottom-up process requiring high levels of cue-item discriminability 2.
A memory is spontaneously triggered when an incidental environmental or internal stimulus matches a distinctive feature of a past experience. This cue is frequently peripheral to the individual's current focal activity but centrally relevant to the encoded memory 3. For example, a specific ambient sound, a passing scent, or an isolated phrase can trigger automatic spreading activation across neural networks. The principle of cue discriminability dictates that cues associated with a multitude of events (low discriminability) rarely trigger specific IAMs, thereby preventing constant cognitive flooding. High-discriminability cues, however, bypass conscious initiation and directly activate the episodic memory trace 2. Retention time also dictates frequency; the probability of spontaneous activation decreases systematically as a function of the temporal delay between encoding and the current context, although highly salient emotional memories exhibit prolonged susceptibility to spontaneous recall 110.
Phenomenological Characteristics of Socially Painful Recall
The subjective experience of an IAM often features a high degree of emotional intensity and specific, detailed reliving of the original event. Compared to voluntary memories, which often retrieve abstract representations of recurrent activities, IAMs are significantly more likely to target highly specific, singular events 3. When an individual experiences an IAM involving embarrassment or shame, the brain essentially forces a re-experiencing of the initial affective state 10.
Psychological research on socially painful memories reveals a specific encoding bias. During an embarrassing social encounter, the brain allocates disproportionate attentional resources to palpable negative emotions - such as self-consciousness or perceived social rejection - rather than contextualizing the misstep as a minor environmental blip 4. Because the memory is processed with an intense affective payload, its subsequent spontaneous retrieval acts as an acute internal stressor. The emotional valance of the memory directly correlates with the intensity of the ensuing autonomic reaction 1311.
Neural Networks Governing Spontaneous Recall
The Default Mode Network and Spontaneous Thought
The neurological predisposition for experiencing involuntary memories is deeply tethered to the activity of the Default Mode Network (DMN). The DMN consists of a set of midline and lateral cortical regions - most prominently the posterior cingulate cortex (PCC), the precuneus, the medial prefrontal cortex (mPFC), and the medial posterior parietal cortex (mPPC) - that exhibit preferential activation during internally oriented cognition, such as mind-wandering, day-dreaming, and autobiographical planning 1213.
Human neuroimaging and electroencephalographic (EEG) studies consistently link the propensity for mind-wandering and spontaneous thought to modulations in lower-frequency spectral power (e.g., slow and delta waves) reminiscent of non-rapid eye movement (NREM) sleep 1214. When an individual is engaged in automated, low-demand tasks, or resting with eyes closed, sensory processing of the external environment is attenuated, and the DMN becomes highly active 14. The ventral PCC is highly integrated within this network and functions as a primary node for memory retrieval and spatial information processing 1516.
The anatomy of the DMN provides the necessary access to the episodic memory system. The posterior cingulate cortex and retrosplenial cortex are densely connected with both lateral and medial parietal areas, providing a direct route to the hippocampal memory system 1517. Consequently, the DMN facilitates the convergence of ventral and dorsal processing streams within the hippocampus, synthesizing the "what" and "where" elements necessary to reconstruct a specific autobiographical episode 15.
Functional Interaction with the Salience Network
The transition from a passive state of mind-wandering to an acute "cringe attack" requires interaction between the DMN and the Salience Network (SN). While the DMN supports the internally directed retrieval of the memory, the SN - anchored by the dorsal anterior cingulate cortex (dACC) and the anterior insula (AI) - evaluates the emotional significance and physiological relevance of the retrieved content 1822.
When a retrieved IAM contains elements of social pain, embarrassment, or shame, the anterior insula detects the highly salient emotional features of the episodic trace. The insula then coordinates with the dACC to switch the brain's processing resources away from passive DMN activity and toward active, threat-oriented affective processing 1822. This abrupt network switching is the neurological substrate of the sudden, jarring realization that characterizes an embarrassing memory intrusion.
Summary of Neural Networks in Involuntary Recall
| Network | Primary Brain Regions | Function in IAM Processing | Role in Vocalization Sequence |
|---|---|---|---|
| Default Mode Network (DMN) | Posterior Cingulate Cortex (PCC), Precuneus, mPFC | Facilitates internal cognition, mind-wandering, and episodic retrieval. | Provides the initial spontaneous memory trace via hippocampal connectivity. |
| Salience Network (SN) | Anterior Insula (AI), Dorsal Anterior Cingulate Cortex (dACC) | Detects salient emotional stimuli; regulates affective pain. | Appraises the memory as socially painful; generates autonomic hyperarousal. |
| Executive Control Network (CEN) | Dorsolateral Prefrontal Cortex (dlPFC), Inferior Frontal Gyrus (IFG) | Top-down cognitive control, inhibition of prepotent responses. | Fails to inhibit the sudden high-arousal impulse, allowing motor leakage. |
| Somato-Cognitive Action Network / Motor | Cingulate Motor Area (CMA), pre-Supplementary Motor Area (preSMA) | Skeletomotor control, action selection, spatial orientation. | Executes the involuntary motor action (vocalization, wincing) via descending spinal tracts. |
The Neurobiology of Social Pain
The Pain Overlap Theory
The profound distress generated by an embarrassing IAM cannot be understood without examining the neurobiology of social pain. The Pain Overlap Theory posits that the experience of social pain - defined as the emotional distress caused by actual or potential damage to one's social connections, including rejection, exclusion, and intense embarrassment - shares significant underlying processing systems with the experience of physical nociception 23241920.
Neuroimaging of individuals experiencing physical pain reliably demonstrates activation in a matrix of regions. While the primary and secondary somatosensory cortices (S1, S2) and the posterior insula process the sensory components of pain (location and intensity), the affective, distressing component of pain is processed cortically by the dACC and the anterior insula 1921. Studies involving social exclusion paradigms, such as the Cyberball task, reveal that experiences of social rejection or shame activate these exact same affective regions - the dACC and AI 1920.
This overlap indicates that, at a neurological level, the brain treats severe social embarrassment as a literal injury. The retrieval of a socially painful IAM re-activates the dACC and AI, inducing a state of acute suffering 1922. Behavioral studies reinforce this physical-social overlap; for example, the administration of acetaminophen (a central physical pain suppressant) has been shown to significantly reduce neural responses to social rejection in the dACC and AI, simultaneously decreasing self-reported daily social pain 20.
Insular Neuromodulation and Genetic Variables
The amplification of the behavioral response to social pain is mediated by chemical convergence in the insular cortex. Animal models employing social exclusion paradigms (e.g., the Fear of Missing Out task in mice) reveal that social exclusion alters physical pain perception via distinct neuromodulatory substrates. Endocannabinoid and oxytocin signaling in the insular cortex exhibit opposing responses during varying states of social exclusion, directly modulating the severity of the behavioral reaction to subsequent stressors 2324.
Furthermore, individual genetic differences heavily influence the intensity of the social pain response. Individuals possessing the low expression allele (MAOA-L) of the monoamine oxidase-A gene display significantly greater dACC reactivity to social exclusion compared to those with the high expression allele. This heightened socioemotional hypersensitivity suggests that specific genetic profiles predispose certain individuals to experience more intense distress upon social rejection 29. Similarly, polymorphisms in the mu-opioid receptor gene (OPRM1) are linked to both physical pain sensitivity and heightened dACC and AI activity in response to social exclusion 19. These neuromodulatory and genetic factors account for the wide individual variance in the frequency and intensity of "cringe attacks."
Evolutionary Origins of Mammalian Distress Vocalizations
The PANIC-GRIEF System and Infant Separation
To understand why a socially painful memory specifically elicits a vocalization, it is necessary to examine the evolutionary history of the mammalian brain. The neural circuitry connecting the dACC to vocal production is an ancient evolutionary adaptation designed to maintain social proximity. In comparative neurobiology, this circuit is frequently referred to as the PANIC-GRIEF system, which relies on a network coursing between the anterior cingulate gyrus, various basal forebrain nuclei, and the dorsal periaqueductal gray (PAG) 521.
In nearly all mammalian species, the survival of altricial (underdeveloped) infants relies entirely on sustained parental care. When mammalian infants are separated from their caregivers or social group, they emit spontaneous, unlearned distress vocalizations (e.g., crying in humans, isolation calls in non-human primates). These vocalizations serve an adaptive evolutionary purpose: they broadcast a high-urgency signal that cues the caregiver to retrieve the infant, preventing prolonged separation 923.
This contrasts sharply with reptilian neurobiology. Reptilian newborns, which receive virtually no parental care, do not possess a developed cingulate gyrus and do not produce separation-distress vocalizations; in their ecological context, vocalizing distress would merely attract predators 2324. The development of the cingulate cortex in mammals allowed for the evolution of social attachment and the vocal signaling of social pain.
Lesion and Stimulation Studies in Primates
The causal role of the dACC in the production of distress vocalizations is definitively supported by primate research. In seminal studies conducted by MacLean and Newman, squirrel monkeys were subjected to localized ablations of the medial prefrontal cortex. The researchers found that lesions specifically targeting the dorsal anterior cingulate cortex completely eliminated the production of separation distress vocalizations (isolation calls) 82324. Lesions to other surrounding areas did not halt the vocalizations, and the dACC-lesioned monkeys retained the physical capacity to make other types of non-distress sounds, proving that the dACC is the specific engine of affective vocal bursts 8.
Conversely, electrical stimulation of the dACC in rhesus monkeys induces the spontaneous, continuous production of distress vocalizations, even when the animal is not isolated or physically harmed 92324. Notably, stimulation of primate cortical areas homologous to human speech centers (e.g., Broca's area) elicits movement of the vocal cords but does not produce distress vocalizations 2324.
This evolutionary architecture remains intact in modern humans. The dACC processes both the distress of physical pain and the distress of social exclusion because, throughout mammalian evolution, social isolation was tantamount to physical death 20. When a human experiences an IAM involving intense embarrassment, the dACC registers severe social damage. Operating on an ancient biological imperative, the dACC triggers a primal distress vocalization - a reflex originally designed to solicit maternal care or signal a critical threat to social cohesion.
The Motor Leakage Hypothesis
Anatomy of the Cingulate Motor Area
The translation of a retrieved emotional memory into a physical vocalization is governed by the structural contiguity of the limbic system and the motor cortex. The cingulate cortex is functionally subdivided; while the anterior portions (areas 24, 25, 32) mediate emotional processing, the midcingulate cortex (MCC) functions as a higher-order motor area, explicitly termed the Cingulate Motor Area (CMA) 1617.
The CMA, located in the cingulate sulcus adjacent to the primary and supplementary motor areas, is crucial for skeletomotor control and action-outcome learning 1617. It is divided into the rostral CMA (associated with avoidance behavior and fear tasks) and the caudal CMA (associated with spatial orientation) 617. The gyral surface of the MCC contains large layer Vb neurons that project directly to the spinal cord, as well as to the primary motor cortex and the pre-supplementary motor area (preSMA) 16.
The anatomical four-region model of the cingulate cortex highlights dense functional connectivity (FC) between these subregions. The pregenual anterior cingulate cortex (pACC) acts as an association area, heavily connected with both the affective networks (subgenual ACC) and the sensorimotor networks (posterior MCC) 25. Because the dACC receives dense nociceptive and social pain inputs, and is immediately adjacent and strongly connected to the CMA, an intense affective signal can readily transmit into motor pathways 25.
Executive Control Failure and the Right Inferior Frontal Gyrus
Under normal psychological conditions, involuntary urges to vocalize or perform sudden motor actions are suppressed by the Executive Control Network (CEN). The internal inhibition of socially undesirable speech - such as the urge to utter a taboo word - relies heavily on the right inferior frontal gyrus (rIFG) and the dorsomedial prefrontal cortex (dmPFC) 1326.
Functional magnetic resonance imaging (fMRI) studies utilizing the Spoonerisms of Laboratory Induced Predisposition (SLIP) paradigm demonstrate that the brain actively deploys neural inhibition to suppress involuntary utterances 26. The rIFG internalizes external social display rules, functioning as a continuous neurological "stop-signal" against inappropriate vocalizations 26.
However, the "motor leakage" hypothesis posits that during an acute IAM ("cringe attack"), the sudden onset of profound autonomic hyperarousal temporarily overwhelms executive function. Anxiety and intense emotional stress are known to hijack cognitive resources, consuming attention and overwhelming the working memory loop 34. The massive spike in dACC activation effectively short-circuits the inhibitory capacity of the rIFG. Without adequate top-down prefrontal suppression, the high-arousal social pain signal "leaks" from the dACC into the adjacent CMA and preSMA, instantly triggering a pre-programmed motor sequence. The result is a brief, uninhibited vocal burst or physical shudder that executes before conscious cognitive control can reassert itself 5726.
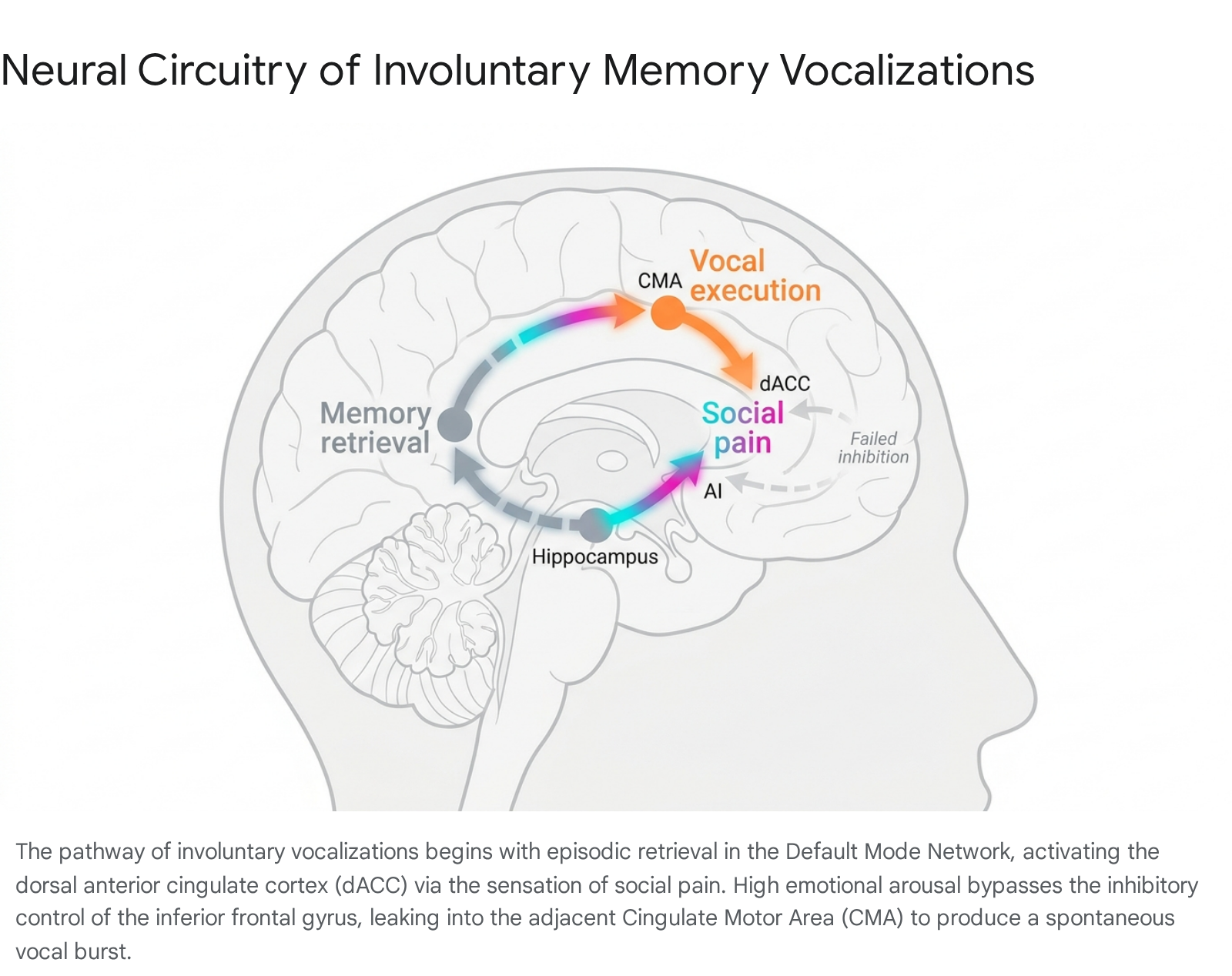
Vocalization as an Emotion-Regulation Strategy
Comparison to Formal Thought Blocking
While the vocalization is mechanically the result of motor leakage, psychological and neurobiological models suggest it also serves a highly functional purpose: it operates as a drastic emotion-regulation strategy intended to abort the painful memory sequence 3527. This behavior aligns with psychoanalytic conceptualizations of "acting out" - a defense mechanism wherein an individual utilizes abrupt physical action to suffocate an unendurable internal mental process, thereby preventing rational but painful conscious interpretation 6.
The sudden termination of the IAM narrative via vocalization bears striking mechanical similarities to the clinical phenomenon of "thought blocking." In formal psychiatric contexts, thought blocking (or derailment of thought) is categorized as a formal thought disorder, classically presenting as a negative symptom in schizophrenia spectrum disorders and severe psychosis 2829. Clinical thought blocking manifests as an abrupt, involuntary cessation of speech and thought, leaving the patient mentally "blank" mid-sentence due to chronic dysfunction in the working memory loop and hypoactivity in the dorsolateral prefrontal cortex 3428.
However, transient thought blocking can also occur in neurotypical individuals as an acute trauma response or during high-stress situations 3428. When an IAM inflicts severe social pain, the memory is experienced as an intrusive threat. The spontaneous vocal burst acts as a self-induced, temporary thought block. By forcefully engaging the motor and vocal apparatus, the brain redirects limited cognitive resources away from the internal retrieval network (DMN) and toward external sensory-motor execution. This effectively derails the train of thought, terminating the painful episodic simulation 3428.
Autonomic Homeostasis and Vagal Regulation
Beyond cognitive disruption, the physical act of vocalizing exerts a direct regulatory influence on the autonomic nervous system (ANS). The emergence of a socially painful IAM triggers a rapid sympathetic nervous system response - a "fight or flight" state characterized by elevated heart rate, blood pressure, and psychological self-fragmentation 131130.
The production of vocal sounds - particularly primal, non-linguistic vocalizations such as groans, sighs, and shouts - forces alterations in respiratory reflexes. To vocalize, the individual must delay immediate exhalation, maintaining contraction of inspiratory muscles to regulate airflow through a partially closed glottis 11. This forced alteration of the respiratory cycle directly stimulates the vagus nerve. According to Polyvagal Theory, engagement of the vagal "brake" is essential for down-regulating sympathetic hyperarousal and reinstating a state of social safeness and physiological calm 31.
Furthermore, research into the psychobiology of crying and distress vocalizations reveals that these expressive actions are correlated with a reduction in plasma cortisol levels, the primary stress hormone 41. Non-linear, "rough" acoustic parameters found in screams and sudden vocal bursts uniquely engage the central autonomic network to modulate visceromotor and neuroendocrine systems 542. Therefore, the IAM vocalization operates as a rapid-response somatic tool to discharge sympathetic tension, shift control to the parasympathetic nervous system, and restore autonomic homeostasis following the internal stressor of the memory 1141.
Cross-Cultural Variances in Expression
Universal Acoustic Signatures versus Cultural Display Rules
While the neural architecture connecting the dACC to the motor cortex is universal, the ultimate physical expression of the IAM vocalization is significantly moderated by cultural parameters 3233. Bioacoustic studies analyzing nonlinguistic vocal bursts (e.g., cries of pain, disgust, joy) across 131 distinct languages have identified consistent, universal acoustic signatures for the expression of pain. Cross-culturally, vocalizations of physical and social pain heavily feature open vowels (e.g., "a") and wide falling diphthongs (e.g., "ai" or "aw"), indicating a conserved evolutionary vocal niche designed to hijack the receiver's salience system 423435.
Despite this universal physiological base, the frequency, volume, and manifestation of spontaneous vocalizations are heavily dictated by cultural "display rules." Display rules are socially learned norms that dictate how, when, and to whom emotions should be expressed 333637.
Individualistic versus Collectivistic Modulations
Extensive cross-cultural psychological research highlights deep disparities in emotional expressivity between different societal structures. Individuals from high-context, collectivistic cultures (such as many East Asian societies) are generally socialized to emphasize emotional restraint, perspective-taking, and social harmony. Conversely, individuals from low-context, individualistic cultures (such as Western European and American societies) are often encouraged to prioritize open emotional expression and individual authenticity 333738.
These cultural environments actively shape the brain's top-down inhibitory mechanisms. Studies comparing empathic responses to social and physical pain show that East Asian participants often report lower overt affective arousal and utilize more subdued physical expressions compared to White British or American counterparts, who are more likely to display highly visible and audible reactions 333850.
In the context of IAMs, an individual raised in a culture that strictly enforces emotional restraint is likely to possess a more rigidly conditioned right inferior frontal gyrus (rIFG). As a result, when an embarrassing memory triggers dACC hyperarousal, the threshold for motor leakage is higher. The culturally conditioned executive network exerts stronger suppression, meaning the IAM might only result in a covert physical reaction (such as an internal wince or altered breathing) rather than a loud, overt vocalization 263337. Thus, while the internal experience of the "cringe attack" remains universal, its acoustic footprint is deeply enculturated.
Clinical Distinctions and Differential Diagnosis
Because IAM vocalizations present as sudden, involuntary motor and phonic outputs, they share distinct phenotypic similarities with various neurological and psychiatric conditions. Distinguishing the neurotypical "cringe attack" from pathological disorders requires careful evaluation of etiology, onset, and underlying neurology.
Primary Tic Disorders (Tourette Syndrome)
Primary tic disorders, most notably Gilles de la Tourette syndrome (TS), are characterized by sudden, rapid, non-rhythmic motor movements and vocalizations that persist for more than one year 394053. TS is a highly heritable, polygenic neurodevelopmental disorder that almost exclusively manifests in early childhood (onset typically between 4 to 6 years of age, peaking around 10 to 12 years) 41.
The critical distinction between TS and IAM vocalizations lies in the antecedent trigger. Tics in TS are universally preceded by a "premonitory urge" - an uncomfortable, localized physical tension or sensory phenomenon that builds until the tic is executed, providing temporary relief 41. The etiology of TS involves dysfunction within the cortico-striato-thalamo-cortical loops 4142. In contrast, IAM vocalizations lack a physical premonitory urge; the antecedent is entirely cognitive and emotional (the retrieved episodic memory). IAM vocalizations also do not present as fixed, rhythmic stereotypies and can emerge at any point in the human lifespan 1041.
Post-Traumatic Functional Tic-Like Behaviors (FTLBs)
A more complex differential diagnosis involves Functional Tic-Like Behaviors (FTLBs). FTLBs are abrupt-onset motor and vocal manifestations that clinically resemble TS but differ by their sudden onset - often during adolescence or early adulthood - female predominance, and lack of response to standard pharmacological treatments (such as aripiprazole or clonidine) 3953.
Crucially, FTLBs are psychogenic symptoms explicitly triggered by severe psychosocial stressors and profound psychological trauma, such as sexual abuse, assault, or acute post-traumatic stress disorder (PTSD) flashbacks 395342. In documented clinical cases, adolescents have developed severe guttural vocalizations and motor jerks immediately following traumatic events, with symptoms worsening during periods of severe stress (e.g., legal trials) 3942. While IAM vocalizations and FTLBs share a mechanism of stress-induced motor leakage, IAMs are triggered by comparatively benign social embarrassment. FTLBs represent a severe, debilitating functional neurological disorder requiring trauma-focused cognitive behavioral therapy (CBT) and eye movement desensitization and reprocessing (EMDR) 3953.
Diagnostic Comparison of Involuntary Vocalizations
| Clinical Feature | Involuntary Memory Vocalizations ("Cringe Attacks") | Primary Tic Disorders (Tourette Syndrome) | Functional Tic-Like Behaviors (FTLBs) | Formal Thought Blocking (Schizophrenia) |
|---|---|---|---|---|
| Primary Trigger | Internal cognitive cue (embarrassing or socially painful memory). | Neurological; preceded by a somatic premonitory urge. | Psychosocial stressors; severe psychological trauma or PTSD. | Neurological/Psychiatric; disorganized working memory loop. |
| Typical Onset Age | Any age; tied to autobiographical memory development. | Early childhood (4 - 6 years), peaking at 10 - 12 years. | Abrupt onset, typically during adolescence or early adulthood. | Late adolescence to early adulthood. |
| Vocal/Motor Characteristics | Contextual to the memory (e.g., groans, cursing, wincing, humming). | Non-rhythmic, repetitive motor stereotypies and phonic tics. | Complex guttural sounds, sudden jerks, highly variable. | Silence mid-sentence; occasional neologisms upon resumption. |
| Underlying Mechanism | Motor leakage via dACC to CMA; acute emotional self-regulation. | Cortico-striato-thalamo-cortical circuit dysfunction. | Psychogenic response to trauma; altered functional connectivity. | Hypoactivity in dorsolateral prefrontal cortex (dlPFC). |
| Clinical Severity | Non-clinical; transient and generally not debilitating. | Chronic; ranges from mild to severe impairment. | Severe; highly disruptive to daily functioning, poor drug response. | Severe; core feature of psychosis and cognitive disorganization. |
Conclusion
The spontaneous vocalizations that accompany the involuntary recall of socially painful autobiographical memories represent a sophisticated intersection of memory retrieval, emotional appraisal, and motor execution. Rather than being erratic neurological anomalies, these "cringe attacks" are deeply structured physiological responses dictated by the evolutionary architecture of the mammalian brain.
The retrieval of a distinctive episodic trace via the Default Mode Network engages the salience network - specifically the dorsal anterior cingulate cortex and the anterior insula. These regions, evolutionarily adapted to process the critical survival threat of mammalian social separation, register the remembered embarrassment as acute social pain. When the resulting autonomic hyperarousal overwhelms the inhibitory capacity of the prefrontal cortex, the distress signal leaks into the adjacent Cingulate Motor Area. The ensuing vocal burst acts as a rapid, self-induced emotion-regulation strategy: it forcefully disrupts the painful cognitive sequence - functioning as transient thought blocking - and initiates respiratory changes that engage vagal tone to restore autonomic homeostasis.
While individual frequency and volume are heavily modulated by cultural display rules and genetic sensitivities to social pain, the underlying mechanism remains a universal testament to the brain's prioritization of social inclusion. In the human neurological paradigm, the memory of social pain is processed with the same immediacy and physical urgency as a physical wound, deploying ancient distress vocalizations to manage the modern experience of embarrassment.