Mechanisms of the gut-brain axis in psychiatric disorders
Introduction: The Evolution of Microbiome-Neuroscience Research
The conceptualization of the microbiota-gut-brain (MGB) axis represents a paradigm shift in modern gastroenterology, neuroscience, and psychiatry. Historically, the central nervous system (CNS) and the gastrointestinal (GI) tract were understood to communicate primarily through the autonomic nervous system to regulate basic, largely mechanical processes such as motility, digestion, and satiety. However, over the past two decades, this traditional view has been radically expanded by the recognition that the trillions of commensal microorganisms residing in the human gut - comprising bacteria, archaea, viruses, and fungi - act as a dynamic, metabolically active organ capable of exerting profound influence on neurodevelopment, behavior, and mental health 123.
The foundational architecture for understanding host-microbiome interactions emerged largely from metabolic and dietary studies. Seminal research from the Gordon and Turnbaugh laboratories, notably the landmark 2006 Nature publication detailing an obesity-associated gut microbiome with an increased capacity for energy harvest, established the critical proof-of-concept that complex physiological traits could be fundamentally driven, and indeed transferred, by microbial communities 4467. By utilizing 16S rRNA gene sequencing on the distal gut microbiota of leptin-deficient ob/ob mice, researchers demonstrated a distinct shift favoring the Firmicutes phylum over the Bacteroidetes. Remarkably, by demonstrating that the colonization of germ-free mice with an "obese microbiota" resulted in a significantly greater increase in total body fat than colonization with a "lean microbiota," this research codified the microbiome as an active variable in host phenotype rather than a passive, symbiotic passenger 467. Subsequent longitudinal studies further confirmed that diet rapidly and reproducibly alters the human gut microbiome, highlighting the extraordinary plasticity of these microbial ecosystems 48.
Building upon this mechanistic validation of microbial trait transfer, pioneers in neurogastroenterology, notably John F. Cryan and Timothy G. Dinan, transposed these principles into the realm of psychiatry. Recognizing that germ-free animals exhibited profound alterations in brain development, stress reactivity, and neurotrophin expression, they introduced the concept of "psychobiotics" in 2013 567. Psychobiotics were originally defined as live microorganisms that, when ingested in adequate amounts, yield mental health benefits by modulating the MGB axis 58. Early preclinical models utilizing these specific strains demonstrated remarkable phenomena: the administration of selected Lactobacillus and Bifidobacterium species could attenuate anxiety-like behavior, enhance stress resilience, and modulate the central expression of critical neuroreceptors, including gamma-aminobutyric acid (GABA) receptors 8910. Furthermore, fecal microbiota transplantation (FMT) from human patients with major depressive disorder (MDD) to germ-free rodents was shown to induce depressive-like behavioral phenotypes in the recipients, seemingly satisfying a modified Koch's postulate for microbiomic psychiatric pathogenesis and cementing the microbiome's role in psychiatric etiology 111617.
However, as the field transitions from its initial discovery phase into the era of rigorous clinical translation, the narrative has grown exponentially more complex. The initial enthusiasm that characterized early psychobiotic research has frequently been co-opted by commercial wellness markets, generating a pervasive "psychobiotic hype" that heavily conflates preliminary, highly controlled animal data with proven clinical efficacy 121913. Current scientific consensus within rigorous gastroenterological and psychiatric literature demands a much more nuanced approach. The relationship between gut dysbiosis and psychiatric disorders such as depression, anxiety, autism spectrum disorder (ASD), and schizophrenia is no longer viewed through a simplistic, unidirectional causative lens. Instead, it is recognized as a highly complex, bidirectional interplay heavily confounded by variables such as host diet, regional baseline microbial diversity, and the profound, often unrecognized antimicrobial effects of psychotropic medications 14152316.
To advance the field, contemporary microbiome-psychiatry research must fulfill several critical imperatives. First, investigations must explicitly define the mechanistic routing of the MGB axis, moving beyond the historical focus on the vagus nerve to fully integrate complex immune, endocrine, and metabolic pathways 1718. Second, the stark translational gap between promising murine models and equivocal human clinical trials must be critically analyzed to recalibrate therapeutic expectations 919. Third, epidemiological analyses must break free from the WEIRD (Western, Educated, Industrialized, Rich, Democratic) cohort bias that currently skews global understanding of baseline microbiome health, acknowledging that the human microbiome is vastly diverse across geographies 2820. Finally, researchers must rigorously decouple true microbial pathogenesis from the confounding artifacts of pharmacology and lifestyle, requiring a fundamental shift in how clinical trials and observational studies are designed 2122. This report provides an exhaustive, critical evaluation of these domains, synthesizing literature from 2023 onwards to establish the current state, and future trajectory, of microbiota-gut-brain axis science.
Deconstructing the Microbiota-Gut-Brain Axis: Primary Communication Pathways
Bidirectional communication along the MGB axis facilitates the constant, dynamic exchange of signals between the GI tract and the CNS. This signaling network is essential for maintaining physiological homeostasis but is equally capable of propagating and exacerbating pathological states when dysregulated. While early research heavily emphasized neural pathways, contemporary biological models recognize four primary, intersecting routes of communication: neural (the vagus nerve and enteric nervous system), neuroendocrine (the hypothalamic-pituitary-adrenal axis), immune (cytokines, mucosal defense, and astrocyte modulation), and metabolic (microbial-derived signaling molecules) 31417.
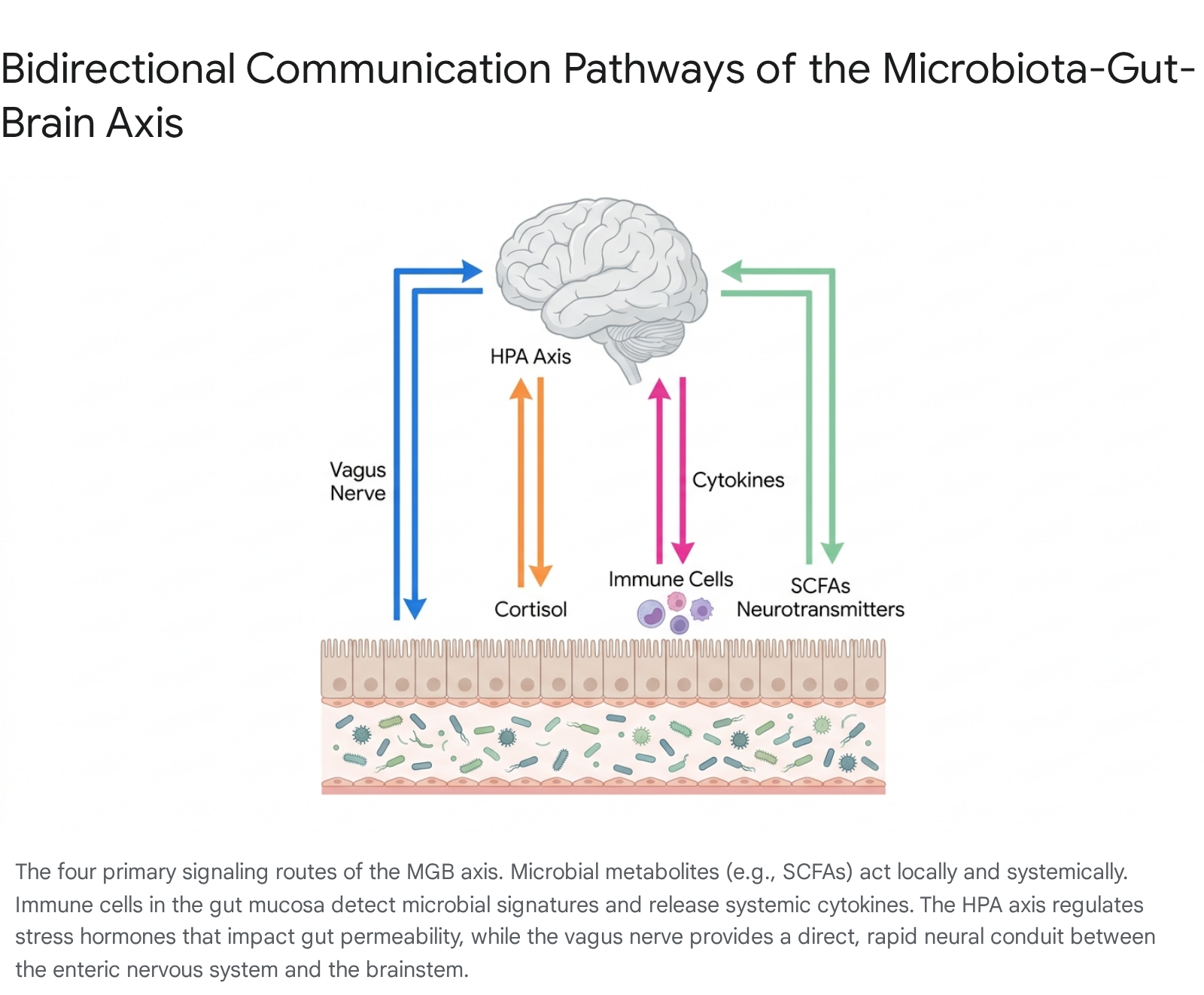
Neural Routing: The Vagus Nerve and Enteric Nervous System
The vagus nerve (cranial nerve X) operates as the most direct and rapid anatomical connection between the gut and the brain. It is a highly complex mixed nerve comprising approximately 80% afferent sensory fibers, which relay information from the viscera to the brain, and 20% efferent motor fibers, which transmit signals from the brain to the gut 2323. The vagal afferent fibers bypass conscious processing and terminate in the nucleus tractus solitarius within the brainstem, from which signals are rapidly relayed via the central autonomic network to higher-order cognitive and emotional centers, including the hypothalamus, amygdala, and prefrontal cortex 162324.
Crucially, vagal afferents do not penetrate the intestinal epithelium to interact directly with the luminal microbiota; rather, they sense microbial signals indirectly through the mucosal interface. Enteroendocrine cells, notably enterochromaffin cells, serve as the primary transducers. These specialized epithelial cells possess neuropod extensions that form synaptic-like connections directly with vagal afferent terminals 25. Upon stimulation by microbial metabolites or structural components, these cells release neurotransmitters - primarily serotonin (5-HT) and glutamate - which rapidly activate vagal firing 2627. Furthermore, certain microbial metabolites, such as short-chain fatty acids (SCFAs), can act on vagal afferents via specific receptor mechanisms. For instance, the SCFA butyrate exerts a direct stimulatory effect on afferent terminals, while long-chain fatty acids, such as oleate, typically require cholecystokinin (CCK)-mediated mechanisms to activate the nerve 23. Additionally, Toll-like receptor 4 (TLR4) is expressed directly on vagal afferent fibers, allowing the nerve to sense bacterial products such as lipopolysaccharide (LPS) and relay inflammatory signals to the brain 23.
The critical necessity of the vagus nerve in microbial-behavioral modulation has been proven surgically. Preclinical models utilizing subdiaphragmatic vagotomy demonstrate that severing this neural conduit completely abolishes the anxiolytic and antidepressant-like effects of specific probiotic strains, such as Lactobacillus rhamnosus JB-1, confirming that neural routing is an indispensable pathway for specific psychobiotic mechanisms 610. Conversely, vagal efferent fibers transmit anti-inflammatory signals back to the gut via the cholinergic anti-inflammatory pathway. By releasing acetylcholine, these efferent fibers can reduce digestive inflammation and reinforce tight junction integrity, mitigating stress-induced intestinal permeability ("leaky gut") and indirectly shaping the environment that dictates microbiota composition 2326.
Neuroendocrine Routing: The Hypothalamic-Pituitary-Adrenal (HPA) Axis
The HPA axis serves as the core neuroendocrine coordinator of the host's physiological stress response and represents a primary systemic target for microbial modulation. Under psychological, environmental, or physiological stress, the hypothalamus secretes corticotropin-releasing factor (CRF), which stimulates the anterior pituitary gland to release adrenocorticotropic hormone (ACTH). This, in turn, drives the adrenal cortex to synthesize and secrete massive quantities of glucocorticoids (primarily cortisol in humans and corticosterone in rodents) 22829.
The gut microbiota is fundamentally responsible for regulating the baseline set-point and the reactivity of the HPA axis. Foundational neurodevelopmental studies in germ-free mice reveal highly exaggerated, hyperactive HPA axis responses to acute stress. This hyper-reactivity can be partially normalized by colonization with a diverse, specific-pathogen-free microbiota, but only if the colonization occurs during a critical early-life developmental window, highlighting the microbiome's role in programming endocrine hardware 12.
In adulthood, chronic psychological stress disrupts microbial ecology, frequently leading to a reduction in SCFA-producing taxa and an overgrowth of pathobionts. This stress-induced dysbiosis increases intestinal permeability by degrading tight junction proteins, a phenomenon that allows the continuous translocation of pathogen-associated molecular patterns (PAMPs), such as LPS, into the systemic circulation 32328. The systemic presence of these bacterial endotoxins profoundly impacts the HPA axis. LPS binding to TLR4 on circulating and tissue-resident immune cells triggers the massive release of pro-inflammatory cytokines. These cytokines can utilize active transport mechanisms across the BBB or interact with circumventricular organs (regions of the brain devoid of an intact BBB) to directly stimulate the hypothalamus, thereby maintaining a chronic state of HPA hyperactivity 272829. The resulting hypercortisolemia further suppresses the host immune system, delays gastric emptying, and alters gut motility, creating a deleterious, self-sustaining feedback loop characteristic of major depressive disorder, severe anxiety, and irritable bowel syndrome (IBS) 22325.
Immune System Signaling: Cytokines, T-Cell Modulation, and Astrocytes
The intestinal mucosa harbors the largest compartment of the host immune system, positioning it as a critical interface for the MGB axis. The microbiota plays an essential, lifelong role in training and maintaining both innate and adaptive immune responses 27. Dysbiosis acts as a primary catalyst for systemic low-grade inflammation, a biological state that is no longer viewed merely as a symptom, but rather as a core pathophysiological driver of treatment-resistant depression, schizophrenia, and neurodegenerative disorders such as Alzheimer's disease and Parkinson's disease 242730.
Microbial modulation of the immune system centers heavily on maintaining the delicate equilibrium between pro-inflammatory Th17 cells and anti-inflammatory regulatory T cells (Tregs) 284041. Beneficial commensal microbes, frequently operating through their SCFA byproducts, promote the differentiation, expansion, and functional capacity of FOXP3+ Tregs within the colonic lamina propria 628. These Tregs are vital for suppressing excessive inflammatory responses and maintaining mucosal tolerance against dietary antigens and commensal flora. In pathological states, a reduction in commensal diversity allows for the proliferation of pathobionts that provoke the secretion of pro-inflammatory cytokines, primarily Interleukin-1β (IL-1β), Interleukin-6 (IL-6), and Tumor Necrosis Factor-alpha (TNF-α) 3132.
These peripheral cytokines influence the central nervous system through multiple highly regulated mechanisms. Once central signaling is initiated, peripheral inflammation rapidly translates into neuroinflammation. Microglia, the resident macrophages of the CNS, become activated, altering their morphology and functional state. This activation leads to the aberrant pruning of healthy synapses and the profound suppression of neurogenesis, particularly within the dentate gyrus of the hippocampus 13334. Furthermore, inflammatory cytokines dramatically alter tryptophan metabolism. They upregulate the enzyme indoleamine 2,3-dioxygenase (IDO), which shunts the essential amino acid tryptophan away from the biosynthesis of serotonin and toward the kynurenine pathway. This generates neurotoxic kynurenine metabolites, directly depleting central serotonin levels while simultaneously inducing excitotoxicity, a dual mechanism that contributes heavily to anhedonia and depressive phenotypes 13435.
Recent systematic reviews encompassing literature through 2024 have also highlighted the critical role of astrocytes in MGB axis immune signaling. Astrocytes are essential for maintaining the BBB and regulating synaptic function. In models of chronic neurodegeneration and acute brain injury, astrocytic reactivity (typically measured by increased glial fibrillary acidic protein, or GFAP, expression) is profoundly influenced by the gut microbiota 36. An increased Bacteroidetes to Firmicutes ratio and compromised gut barrier integrity have been directly linked to increased GFAP expression and astrocyte dysfunction. Analyses of microbial metabolites and fecal microbiota transplants suggest that the gut microbiota dictates astrocyte biology and neuroinflammation, making astrocytes a critical, though highly complex, cellular target within the MGB axis 36.
Microbial Metabolites: SCFAs and Neurotransmitters
Microbial metabolites serve as the highly diverse biochemical currency of the MGB axis, exerting potent effects both locally within the gastrointestinal tract and systemically across the BBB.
Short-Chain Fatty Acids (SCFAs): Acetate, propionate, and butyrate are the primary SCFAs produced via the anaerobic bacterial fermentation of indigestible dietary fibers and resistant starches 2728. SCFAs interact with highly specific G protein-coupled receptors, particularly free fatty acid receptor 2 (FFAR2, also known as GPR43) and free fatty acid receptor 3 (FFAR3, or GPR41) 1728. These receptors are expressed abundantly on enteroendocrine cells, mucosal immune cells, and vagal afferents 1728. Butyrate, in particular, is critical for both neuro-gastrointestinal and systemic health. Locally, it serves as the primary energy source for colonocytes, reinforcing the physical integrity of the epithelial barrier by upregulating tight junction proteins. Systemically, butyrate operates as a potent histone deacetylase (HDAC) inhibitor. By exerting this epigenetic control, butyrate effectively suppresses neuroinflammation and enhances the expression of brain-derived neurotrophic factor (BDNF) in the hippocampus and cortex, thereby promoting synaptic plasticity, memory formation, and recovery from neurological insults 17273648.
Neurotransmitters and Precursors: The gut microbiome encodes the complex enzymatic machinery required to synthesize and metabolize many of the exact same neurotransmitters utilized by the human CNS, including GABA, dopamine, serotonin, and norepinephrine 161327. While the prevailing scientific consensus maintains that microbially derived neurotransmitters do not cross the blood-brain barrier in significant, physiologically active quantities, they act powerfully on local receptors within the enteric nervous system and on vagal afferents 33. For example, upwards of 90% of the body's entire serotonin supply is synthesized by enterochromaffin cells in the gut, a process strictly regulated by microbial metabolites, including secondary bile acids and SCFAs 2537. Additionally, specific strains of Lactobacillus and Bifidobacterium are prolific producers of GABA, the primary inhibitory neurotransmitter. Increased luminal GABA can bind directly to receptors on the enteric nervous system, initiating complex vagal signaling cascades that ultimately influence central GABAergic tone, exerting measurable anxiolytic effects in preclinical models 102427.
Summary of Major Communication Pathways
The following table synthesizes the primary communication pathways of the MGB axis, detailing the proposed physiological mechanisms and summarizing the current strength of scientific evidence based on recent systematic evaluations and umbrella reviews from 2024 and 2025 31617232736.
| Pathway Route | Primary Mediators | Proposed Physiological Mechanism | Current Strength of Evidence (2024 - 2025) |
|---|---|---|---|
| Neural (Vagus & ENS) | Vagal afferents, Enteric Nervous System, Acetylcholine | Direct anatomical connection. Microbes stimulate enterochromaffin cells to release serotonin/glutamate, altering vagal firing rates to the brainstem (nucleus tractus solitarius), modulating central mood and stress circuits. Efferent fibers reinforce gut tight junctions. | High. Vagotomy models conclusively demonstrate the necessity of the vagus nerve for specific behavioral effects (e.g., L. rhamnosus JB-1 anxiolysis). |
| Immune System | Cytokines (IL-6, TNF-α, IL-10), Tregs, Th17 cells, Astrocytes (GFAP) | Dysbiosis increases intestinal permeability ("leaky gut"), allowing LPS translocation. This triggers systemic inflammation. Peripheral cytokines cross the BBB or stimulate vagal receptors, inducing neuroinflammation, microglial activation, and astrocyte reactivity. | High. Robust clinical correlation between elevated peripheral inflammatory markers, dysbiosis, and major depressive disorder. Mechanistic links well-established in both animal and human cohorts. |
| Neuroendocrine | HPA Axis (Cortisol / Corticosterone) | Microbes regulate the baseline stress set-point. Chronic stress alters microbiota, maintaining hypercortisolemia via inflammatory feedback loops. Cortisol subsequently impacts gut motility, delays emptying, and compromises mucosal barrier function. | Moderate-High. Clear evidence in germ-free mice demonstrating exaggerated HPA response. Human data confirms HPA dysregulation in stress-related GI disorders (e.g., IBS) linked to profound microbial shifts. |
| Microbial Metabolites | SCFAs (Butyrate), GABA, Serotonin, Bile Acids | SCFAs act systemically as HDAC inhibitors, promoting BDNF and reinforcing the BBB. Microbially synthesized neurotransmitters act locally on the ENS to initiate vagal signaling or influence systemic precursor availability (e.g., Tryptophan shunting). | Moderate. Strong mechanistic data in cellular and murine models for SCFAs. However, evidence demonstrating that direct oral supplementation of microbially-derived neurotransmitters effectively alters human brain chemistry remains limited and highly debated. |
The Translational Gap: Preclinical Promise vs. Clinical Reality
The immense scientific enthusiasm surrounding the MGB axis has been largely fueled by extraordinary findings in preclinical animal models. Through the sophisticated use of germ-free animals, specific-pathogen-free (SPF) models, and highly controlled environmental conditions, researchers have demonstrated that introducing specific microbial strains can unequivocally alter rodent behavior, neurochemistry, and neurophysiology 110. However, as these targeted interventions move into human clinical trials, the outcomes almost universally fail to mirror the robust efficacy seen in rodents, exposing a significant and deeply problematic translational gap.
The divergence between preclinical success and clinical ambiguity is most strikingly illustrated by the rigorous evaluation of specific "psychobiotic" strains, notably within the Lactobacillus rhamnosus species, encompassing well-studied strains such as JB-1 and GG 69.
Preclinical Efficacy: The Case of L. rhamnosus JB-1
In highly controlled animal models, L. rhamnosus JB-1 operates as an exceptionally effective psychobiotic. A comprehensive 2024 meta-analysis evaluating L. rhamnosus in stress-induced anxiety models reported significant anxiolytic effects in 12 out of 15 selected studies 6. In specialized behavioral models utilizing the inherently anxious BALB/c mouse strain, chronic administration of JB-1 consistently reduced stress-induced hyperthermia, significantly decreased immobility time in the forced swim test (FST) and tail suspension test (TST) - both indicative of potent antidepressant-like activity - and dramatically increased open-arm exploration time in the elevated plus maze (EPM) 6810.
Mechanistically, JB-1 administration induced profound central alterations. Neurochemical analyses revealed significant modulation of regional GABA receptor mRNA expression - specifically upregulating $GABA_{B1b}$ in the cortical regions (cingulate and prelimbic areas) while concomitantly reducing $GABA_{A\alpha2}$ expression in the prefrontal cortex and amygdala 610. Furthermore, the strain profoundly attenuated the HPA axis response, significantly blunting the surge of plasma corticosterone release following an acute 30-minute restraint stressor 8. These neurochemical, endocrine, and behavioral effects were proven to be entirely vagus nerve-dependent; subjecting the mice to subdiaphragmatic vagotomy prior to administration completely eradicated the therapeutic benefits of the probiotic, establishing a direct causal mechanism 61038.
Clinical Reality: The Human Translational Failure
Driven by these immensely compelling preclinical findings, clinical researchers attempted to translate the efficacy of L. rhamnosus JB-1 to human subjects. In a rigorously designed, randomized, single-blind, placebo-controlled cross-over trial, a cohort of healthy male volunteers was administered $1 \times 10^9$ colony-forming units (CFU) of JB-1 daily for four weeks 939.
The results of the human trial were unequivocally negative. Treatment with JB-1 was not superior to placebo in any measured psychometric or physiological domain. It completely failed to alter subjective, self-reported measures of mood, anxiety, stress, or sleep quality. Physiologically, it did not attenuate the HPA axis response - measured via salivary cortisol levels - to the socially evaluated cold-pressor test (SECPT), a rigorous method for inducing acute stress in humans. Furthermore, extensive cognitive batteries yielded no improvements in cognitive performance, visuospatial memory, attention switching, or emotion recognition. Associated resting electroencephalography (EEG) measures showed no relevant alterations, nor did the probiotic induce any significant anti-inflammatory effects based on analyses of basal or stimulated peripheral cytokine levels (IL-1β, IL-6, TNF-α, IL-10) 59.
While other closely related strains, such as Lactobacillus rhamnosus GG, have demonstrated some modest clinical benefits in specific human trials - such as slight improvements in NIH Toolbox Total Cognition Scores among older adults and marginal reductions in inflammatory markers in patients with chronic ethanol exposure - the overall magnitude of the effect is vastly diminished compared to the sweeping behavioral reversals routinely observed in rodent models 323552.
Evaluating the Broader Evidence Base: 2024 - 2025 Umbrella Reviews
This translational gap is not limited to isolated strains like JB-1; it is a systemic characteristic of the current psychobiotic literature. Recent umbrella reviews (comprehensive overviews of multiple systematic reviews) published in 2024 and 2025 have critically evaluated the clinical efficacy and methodological quality of probiotic, prebiotic, and synbiotic interventions for psychiatric symptoms globally 4041.
A comprehensive 2025 umbrella review analyzing 30 systematic reviews of randomized controlled trials (RCTs) found that while probiotic interventions demonstrated statistically consistent, albeit modest, benefits for patients formally diagnosed with major depressive disorder (Standardized Mean Difference [SMD] = -0.50), the findings for anxiety disorders were markedly inconsistent and largely statistically insignificant (SMD = -0.19) 40. The evidence supporting prebiotics alone for depression was even weaker (SMD = -0.25), and completely inconclusive for anxiety 40.
Crucially, the methodological quality of the underlying literature is deeply concerning. An umbrella review analyzing 30 systematic reviews of probiotic interventions revealed that over 76.6% of these reviews were graded as moderate, low, or critically low quality using the rigorous AMSTAR-2 quality appraisal tool. This indicates a pervasive risk of bias in the current clinical literature 4055. The primary reasons for downgrading the quality of these reviews included the lack of prior protocol registration, failure to adequately assess the risk of bias of individual included studies, extreme statistical heterogeneity, and a failure to account for publication bias 42.
The profound difficulty in translating animal data to humans stems from insurmountable biological and environmental discrepancies. Preclinical psychobiotic models utilize genetically identical inbred mice (e.g., BALB/c or C57BL/6), housed in sterile, highly controlled laboratory environments, fed strictly standardized chow, and subjected to artificial, acute, and perfectly uniform stressors 68. In stark contrast, human psychiatric etiology is highly heterogeneous, deeply polygenic, and shaped by decades of complex environmental, socio-economic, dietary, and psychological variables that simply cannot be controlled for, much less replicated, in a laboratory mouse model 4344.
Comparative Outcomes: Lactobacillus rhamnosus Translational Gap
| Domain | Animal Models (L. rhamnosus JB-1 / GG) | Human Clinical Trials (L. rhamnosus JB-1 / GG) |
|---|---|---|
| Behavioral Symptoms (Anxiety/Depression) | Consistent, highly significant reduction in anxiety-like (Elevated Plus Maze) and depressive-like (Forced Swim Test) behaviors in stressed murine models. 68 | Equivocal to negative. No significant improvement in subjective mood, anxiety, or stress scores over placebo in healthy volunteers. 59 |
| HPA Axis Regulation | Robust, predictable attenuation of stress-induced plasma corticosterone levels. Rapid restoration of baseline HPA tone following chronic stress models. 810 | Complete failure to demonstrate effect on salivary cortisol response during rigorous stress induction (socially evaluated cold-pressor testing). 59 |
| Neurochemical Alterations | Significant, region-specific modulation of central GABA receptor expression ($GABA_{B1b}$ up, $GABA_{A\alpha2}$ down). 610 | Central neurochemistry unquantifiable directly in vivo, but no corollary improvements in associated EEG resting states or cognitive performance tasks. 59 |
| Inflammatory Markers | Significant reduction in pro-inflammatory cytokines (IL-6, TNF-α) and increased regulatory T-cell markers in gut and systemic tissue. 652 | No significant changes observed in either basal or stimulated peripheral circulating cytokine levels (IL-1β, IL-6, TNF-α, IL-10). 59 |
Disentangling Correlation from Causation: The Confounding Roles of Diet and Medication
A fundamental vulnerability in MGB axis research, and the primary driver of the aforementioned "psychobiotic hype," is the persistent and scientifically dangerous misinterpretation of correlation as causation. Cross-sectional observational studies routinely and reliably demonstrate that patients with psychiatric conditions - particularly major depressive disorder, bipolar disorder, and schizophrenia - harbor distinct gut microbial signatures. These signatures are generally characterized by a marked reduction in SCFA-producing taxa (e.g., Faecalibacterium, Bifidobacterium) and a concomitant enrichment of pro-inflammatory bacterial families (e.g., Eggerthella, Enterobacter) 16124445. However, simply observing dysbiosis in a depressed or anxious patient does not inherently dictate that the dysbiosis caused the depression.
This directional ambiguity is severely complicated by ubiquitous environmental confounders. The human gut microbiome is an exquisitely dynamic ecosystem, fiercely responsive to daily extrinsic inputs 22. Two of the most potent drivers of microbial community structure - diet and pharmaceutical medication - are intimately tied to psychiatric illness, creating a tangled web of reverse causality that observational studies frequently fail to address 232122.
Diet as a Primary Driver of Reverse Causality
Psychiatric disorders profoundly alter human behavior, most notably long-term dietary habits. Individuals suffering from major depression, severe anxiety, or anhedonia frequently undergo massive shifts in appetite and food selection, heavily gravitating toward highly processed, low-fiber, high-sugar, and high-fat "Western" diets that provide rapid caloric density and short-term dopaminergic reward 16.
Diet, more than almost any other variable, dictates microbial architecture. A diet devoid of complex fermentable carbohydrates rapidly starves SCFA-producing bacteria. To survive, the microbiome undergoes a structural shift, leading to a compensatory proliferation of proteolytic and mucin-degrading pathobionts. These organisms erode the intestinal mucosal barrier to harvest endogenous glycoproteins, triggering the systemic inflammation associated with depression 1646. Therefore, the dysbiosis observed in a depressed cohort may simply be the downstream artifact of a depression-induced dietary shift, rather than the pathogenic, neuro-inflammatory origin of the psychiatric state itself. Foundational studies have repeatedly demonstrated that extreme dietary shifts rapidly and reproducibly alter the human microbiome within a matter of days, often superseding the influence of host genetics 4822. Failing to strictly control for detailed, long-term dietary intake via comprehensive food frequency questionnaires in clinical trials effectively renders comparative microbiome analyses between depressed and healthy cohorts moot.
Pharmacomicrobiomics: The Unseen Influence of Psychotropic Drugs
Perhaps the most significant, yet historically underreported, confounding variable in microbiome-psychiatry research is the ubiquitous use of psychotropic medications. Recent, highly rigorous systematic reviews from 2024 and 2025 reveal that common psychiatric drugs - including SSRI antidepressants, atypical antipsychotics, and mood stabilizers - exert profound, direct antimicrobial effects on the gut microbiome, independent of the underlying psychiatric condition they are prescribed to treat 214748.
The nascent and rapidly expanding field of pharmacomicrobiomics demonstrates that drugs initially designed to target human CNS receptors interact directly and powerfully with bacterial metabolism in the gut lumen 1547. For instance, antipsychotics such as quetiapine and mood stabilizers like lithium have been shown to significantly alter both alpha and beta diversity metrics in medicated patients 6349. While some of these drug-induced shifts may incidentally enrich beneficial bacteria, the vast majority are deeply disruptive.
Long-term usage of psychotropic drugs has been strongly and consistently associated with an increased prevalence of gut bacteria carrying multidrug antibiotic resistance genes. Furthermore, the odds of isolating extended-spectrum beta-lactamase (ESBL)-producing Enterobacteriaceae are notably higher in patients treated with antipsychotic drugs for over a year 486350. Furthermore, antipsychotic administration is linked to the rapid proliferation of pathogenic bacteria associated with severe metabolic dysfunctions, providing a clear mechanistic explanation for the severe metabolic side effects (e.g., rapid weight gain, dyslipidemia, insulin resistance) frequently observed in patients prescribed these life-saving medications 4950. Interestingly, these microbiomic shifts are frequently more pronounced in female patients, who demonstrate greater microbial diversity shifts following treatment than male cohorts 486349.
Crucially, the microbiome is not merely a passive recipient of these pharmaceutical insults; it actively modifies the pharmacokinetics of the drugs themselves 15. The efficacy of, and tolerance to, specific antidepressants can be accurately predicted by baseline microbiome composition 47. In trials involving bipolar disorder, responders to quetiapine - those experiencing improved depressive symptom scores - displayed distinct post-treatment microbial profiles that more closely resembled those of healthy individuals compared to non-responders, suggesting that the microbiome dictates the bioavailability and active metabolism of the psychotropic agent 486350.
Consequently, when researchers analyze the stool of medicated, severely depressed patients and compare it to unmedicated, healthy controls, the identified "disease signature" is almost entirely contaminated by the "medication signature" 2321. As evidenced by large-scale retrospective studies utilizing years of electronic health records, past and present medication usage accounts for a massive proportion of inter-individual microbiome variability, often completely superseding the effect of the disease itself 21. Debunking the psychobiotic hype requires an explicit, field-wide acknowledgment that without rigorous, mathematical stratification for polypharmacy and dietary habits, correlative microbiome signatures cannot be utilized to establish psychiatric pathogenesis.
The Global Microbiome: Overcoming the WEIRD Cohort Bias
The generalizability of current MGB axis research is severely restricted by a pervasive, systemic epidemiological bias. The overwhelming majority of foundational microbiome data, including large-scale initiatives like the Human Microbiome Project, have been derived almost exclusively from WEIRD (Western, Educated, Industrialized, Rich, Democratic) populations 1920. Relying heavily on cohorts from the United States, Europe, and select urban centers in Asia creates a fundamental, structural distortion in our biological understanding of a "healthy" or "normal" baseline microbiome 1920.
The Impact of Geography and Lifestyle
The human gut microbiome is deeply entwined with geography, cultural dietary practices, and socioeconomic status. The transition from traditional agro-pastoral or hunter-gatherer lifestyles to highly urbanized, industrialized environments fundamentally rewrites the microbial architecture of the human gut 2820. Western populations, consuming diets notoriously high in processed foods and subjected to high rates of broad-spectrum antibiotic utilization and heavily sanitized environments, typically present with significantly lower gut microbial alpha diversity compared to non-Western populations 2846.
When psychiatric-microbiome associations are established strictly within WEIRD populations, the diagnostic and therapeutic applications of those findings fail to translate globally. A marker of dysbiosis in an urban North American patient (e.g., specific shifts in the Firmicutes to Bacteroidetes ratio) may represent a completely normal, homeostatic state in a rural South American patient due to radically different dietary inputs, such as varying loads of complex plant polysaccharides versus processed fats and animal proteins 1620.
Evidence from Diverse Cohorts: ASD, SAMBAR, and Latinbiota
The critical necessity of geographical diversity in microbiome research is powerfully highlighted by recent studies investigating Autism Spectrum Disorder (ASD). A 2021 longitudinal study comparing cohorts in Arizona and Colorado found that the gut microbiomes of children with ASD differed significantly between the two geographic locations. In Arizona, but not Colorado, GI symptoms were higher in individuals with ASD than controls. Crucially, gut microbiome composition was significantly associated with ASD only when researchers statistically controlled for study site location. The researchers concluded that geographical differences in gut microbiome composition contribute heavily to the wildly inconsistent findings regarding the ASD microbiome in global literature 66.
Recent initiatives are beginning to rectify this critical data deficit by targeting non-WEIRD populations. The South Asian MicroBiome ARray (SAMBAR) cohort, an extensive 16S gut microbiome study encompassing 575 adults from 10 socio-culturally diverse South Asian communities across both ancestral villages and urban centers, revealed profound insights. SAMBAR microbiomes occupy a distinct compositional space with a much stronger correlation to geography and cultural community membership than to specific lifestyle transitions. Microbiome responses to urbanization proved highly heterogeneous even at regional scales, underscoring the need for community-specific investigations of health impacts rather than blanket applications of Western microbiome models 2867.
Similarly, the Latinbiota Consortium aims to bridge the severe data gap in Latin America and the Caribbean. This region is characterized by unique ethnic diversity, distinct dietary traditions, and a highly complex dual burden of disease: pervasive infectious diseases (undernutrition, parasites) overlapping with rising rates of chronic metabolic diseases (obesity, diabetes). Addressing these overlapping health challenges requires locally generated data, as relying on high-income country models to develop microbiome-based interventions for Latin American populations is fundamentally flawed 20.
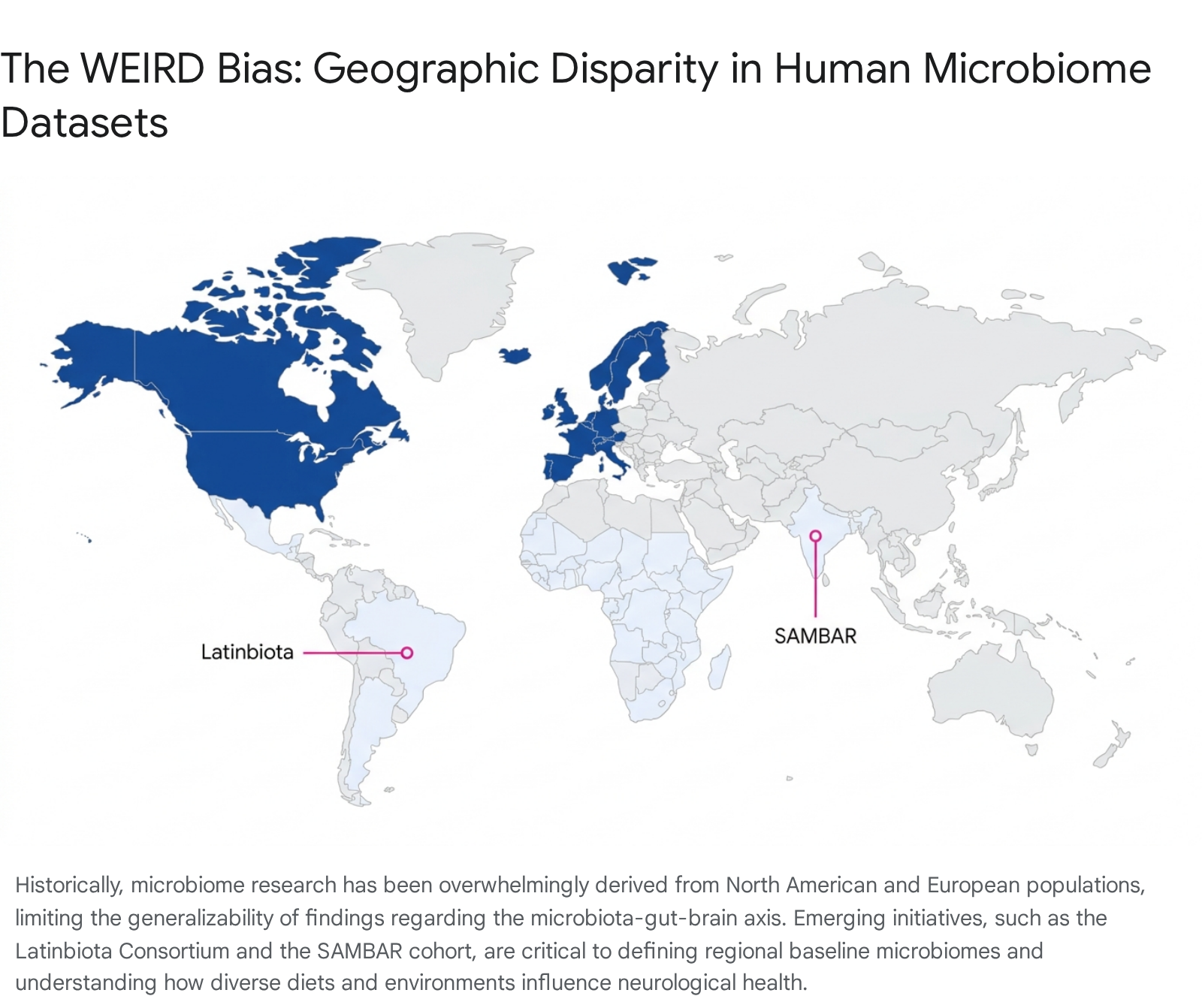
Until global cohort data is robustly integrated into psychiatric microbiome models, the development of universal diagnostic biomarkers or broad-spectrum psychobiotic therapies remains scientifically premature. Effective microbiome-based interventions will likely necessitate highly localized, region-specific formulations tailored to the underlying microbial ecology and genetic background of the target population.
Methodological Imperatives and Future Directions
The current landscape of microbiota-gut-brain axis research is characterized by immense biological promise hampered by severe methodological limitations. To successfully navigate the translational gap, decouple correlation from causation, and advance psychiatric treatment, the field must adopt more rigorous, high-resolution analytical frameworks.
Advancing Beyond 16S rRNA Sequencing
A primary driver of inconsistency in the literature is the continued over-reliance on 16S rRNA amplicon sequencing. While historically cost-effective and essential for early discoveries, 16S sequencing provides only low-resolution taxonomic profiling, typically failing to accurately distinguish between highly similar bacterial species or strains 1928. Given that psychobiotic effects are exquisitely strain-specific - as seen in the wildly differing efficacies of various Lactobacillus rhamnosus isolates - genus-level data is entirely inadequate for therapeutic development 631.
Future research must standardize the use of whole-genome shotgun metagenomics, which not only provides precise strain-level identification but also elucidates the functional genetic potential of the microbiome. Crucially, metagenomics must be paired with transcriptomics and high-throughput metabolomics 1351. Identifying which microbial species are present in the gut is far less clinically relevant than quantifying what those microbes are actively synthesizing and secreting into the host system (e.g., SCFAs, tryptophan metabolites, inflammatory mediators) 192427.
Machine Learning and Model Validation
The integration of Artificial Intelligence (AI) and machine learning, such as Support Vector Machine-Recursive Feature Elimination (SVM-RFE) algorithms, is rapidly transforming microbiome analysis by managing the massive dimensionality of multi-omic datasets 1951. However, current algorithmic applications are frequently hindered by "black box" methodologies and reliance on undersized, highly homogeneous training datasets, leading to unacceptably high rates of overfitting and misinterpretation 19. The establishment of large-scale, longitudinal, and geographically diverse repositories is essential to train predictive models capable of distinguishing true pathophysiological signals from dietary and pharmacological noise.
Toward Precision Pharmacomicrobiomics
The era of the "one-size-fits-all" commercial psychobiotic supplement is biologically unfounded. The future of MGB axis intervention lies in precision pharmacomicrobiomics 1347. Clinicians will inevitably need to profile a patient's baseline microbiome architecture before prescribing psychotropic medications to predict pharmaceutical efficacy and proactively mitigate adverse metabolic events 63. Therapeutic interventions will shift away from generic probiotics toward customized synbiotics (calculated combinations of tailored probiotics and specific prebiotic fibers) designed to predictably modulate targeted pathways - such as administering engineered butyrate-producing consortia to repair compromised BBB integrity, or prescribing targeted phage therapies to eradicate specific pro-inflammatory pathobionts without disrupting the broader commensal ecosystem 133340.
Conclusion
The microbiota-gut-brain axis is irrefutably a central regulatory network governing neurodevelopment, stress resilience, and psychiatric health. Bidirectional communication is orchestrated through an elegant, highly integrated physiological matrix involving neural vagal signaling, neuroendocrine HPA axis regulation, complex immune modulation via cytokines and astrocytes, and microbial metabolite interactions. The foundational, historical recognition that gut microbiota can transfer phenotypic traits has successfully catalyzed a new era of neuro-gastroenterological research, proving that the gut is not merely a digestive tube, but a core component of human neurobiology.
However, the field currently stands at a critical, highly scrutinized inflection point. The leap from highly controlled, germ-free animal models to the complex, heavily confounded reality of human clinical psychiatry has exposed severe translational gaps, particularly regarding the clinical efficacy of widely touted psychobiotic strains like Lactobacillus rhamnosus. Pervasive public and commercial claims implying direct causation between gut dysbiosis and mental illness must be rigorously tempered by the reality of profound confounding variables. The diets patients consume and the psychotropic medications they are prescribed exert massive, bidirectional, and often antimicrobial influences on the microbiome, frequently generating microbial artifacts that merely mimic disease pathogenesis.
To realize the extraordinary clinical potential of the MGB axis, the scientific community must elevate methodological standards. This requires an uncompromising transition toward high-resolution shotgun metagenomics, the establishment of longitudinal human trials that rigorously control for pharmacology and diet, and a dedicated, well-funded effort to expand global datasets beyond WEIRD cohorts to encompass true human diversity. By discarding the reductionist search for a universal "psychobiotic cure" and embracing the staggering biological complexity of precision microbiomics, the medical community can ultimately harness the gut ecosystem to foster profound neurological and psychiatric health.