How Dopamine and Reward Prediction Error Shape Motivation
Dopamine is widely misunderstood as the brain's "pleasure chemical," but it is actually a highly sophisticated learning signal that tracks surprise and expectation. Reward prediction error is the mathematical difference between the reward you expect to receive and the reward you actually experience. This continuous subconscious calibration mechanism drives human motivation, shapes our deepest habits, and explains the biological trap of behavioral and substance addiction.
The Myth of the Pleasure Molecule
For decades, popular culture and oversimplified science communication have equated dopamine with joy, satisfaction, and hedonism. This misconception stems from early neurobiological studies demonstrating that rewarding activities - such as eating calorie-dense foods, engaging in social interaction, or using illicit drugs - trigger massive dopamine release in the brain 12. Because dopamine spikes occurred during enjoyable activities, observers naturally assumed the chemical was the biological equivalent of pleasure. However, modern neuroscience has fundamentally decoupled dopamine from the subjective experience of enjoying a reward.
In the late 1980s and early 1990s, researchers like Kent Berridge demonstrated a critical neurobiological distinction between "wanting" (incentive salience) and "liking" (hedonic pleasure) 134. Through a series of elegant experiments, scientists depleted the brains of animal models of nearly all their dopamine. The results were striking: the animals lost all motivation to seek out food and would starve to death if left alone 4. Yet, if researchers placed sugar directly into the mouths of these dopamine-depleted animals, they still exhibited the exact same facial expressions associated with pleasure and satisfaction 4. They still liked the reward, but they no longer wanted it enough to pursue it.
Dopamine, therefore, is not the reward itself. It is the neurobiological spark that signals the brain to move toward a specific goal. It is a molecule of anticipation, pursuit, and learning, designed by evolution to optimize behavior for future survival 155. It focuses our attention on environmental cues that predict valuable outcomes and provides the motivational drive necessary to acquire them. When you smell coffee brewing from another room, the pleasure is not the smell itself; the dopamine is what compels you to get out of your chair and walk toward the kitchen 6.
The Discovery of Reward Prediction Error
The foundational understanding of dopamine's true function emerged in the 1990s through the pioneering work of neuroscientist Wolfram Schultz. While sitting in a laboratory at the University of Fribourg in Switzerland, Schultz was recording the activity of single dopamine neurons in the midbrains of awake macaque monkeys 87. He was initially attempting to figure out how neurons that release dopamine contribute to motor control, aiming to better understand movement disorders like Parkinson's disease 7.
Schultz and his colleagues developed an experiment where they monitored a monkey as it grabbed a treat from a small box with a door. The researchers assumed the dopamine neurons would fire as the monkey physically moved its arm to grab the treat. Instead, they noticed a peculiar pattern. The neurons were not responding to the movement itself; they were lighting up the moment the door snapped open to reveal the food 7. To investigate further, the researchers began delivering unexpected squirts of apple juice to the monkeys. Initially, the unexpected delivery of the sweet juice caused a massive, immediate spike in the firing rate of the dopamine neurons 8.
However, as the researchers began pairing the juice delivery with a predictive cue - such as a specific light or a tone - a fascinating transition occurred. The dopamine spike began to shift backward in time 48. Over repeated trials, the neurons began firing robustly at the exact moment the tone sounded, but they remained completely silent when the actual juice arrived moments later 89.
Schultz realized that the dopamine system is not a reward detector; it is a surprise detector. When the juice was preceded by a reliable tone, the arrival of the juice was no longer surprising. The brain had successfully updated its internal model of the environment to expect the juice whenever the tone played.
The Temporal Difference Algorithm
Schultz's meticulous electrophysiological work piqued the interest of computational neuroscientists and mathematicians, particularly Peter Dayan and P. Read Montague. In 1997, they published a landmark paper in Science analyzing Schultz's primate data through the lens of computer science 4710. They noticed a striking, almost impossible pattern: the biological firing behavior of the macaque dopamine neurons was mathematically identical to an artificial intelligence mechanism known as Temporal Difference (TD) reinforcement learning, originally developed by computer scientists Richard Sutton and Andrew Barto 471011.
In Temporal Difference learning, an artificial agent learns not by waiting for a final outcome, but by continuously comparing its current prediction with its next prediction, spreading learning across an entire chain of events 410. The researchers realized that the mammalian brain continuously calculates a mathematically precise equation: Reward Prediction Error (RPE) = Actual Reward - Expected Reward 912. The brain acts as a relentless biological calculator, scanning the environment and using this formula to determine which cues are worth paying attention to and which actions should be repeated 4.
The Three States of Prediction Error
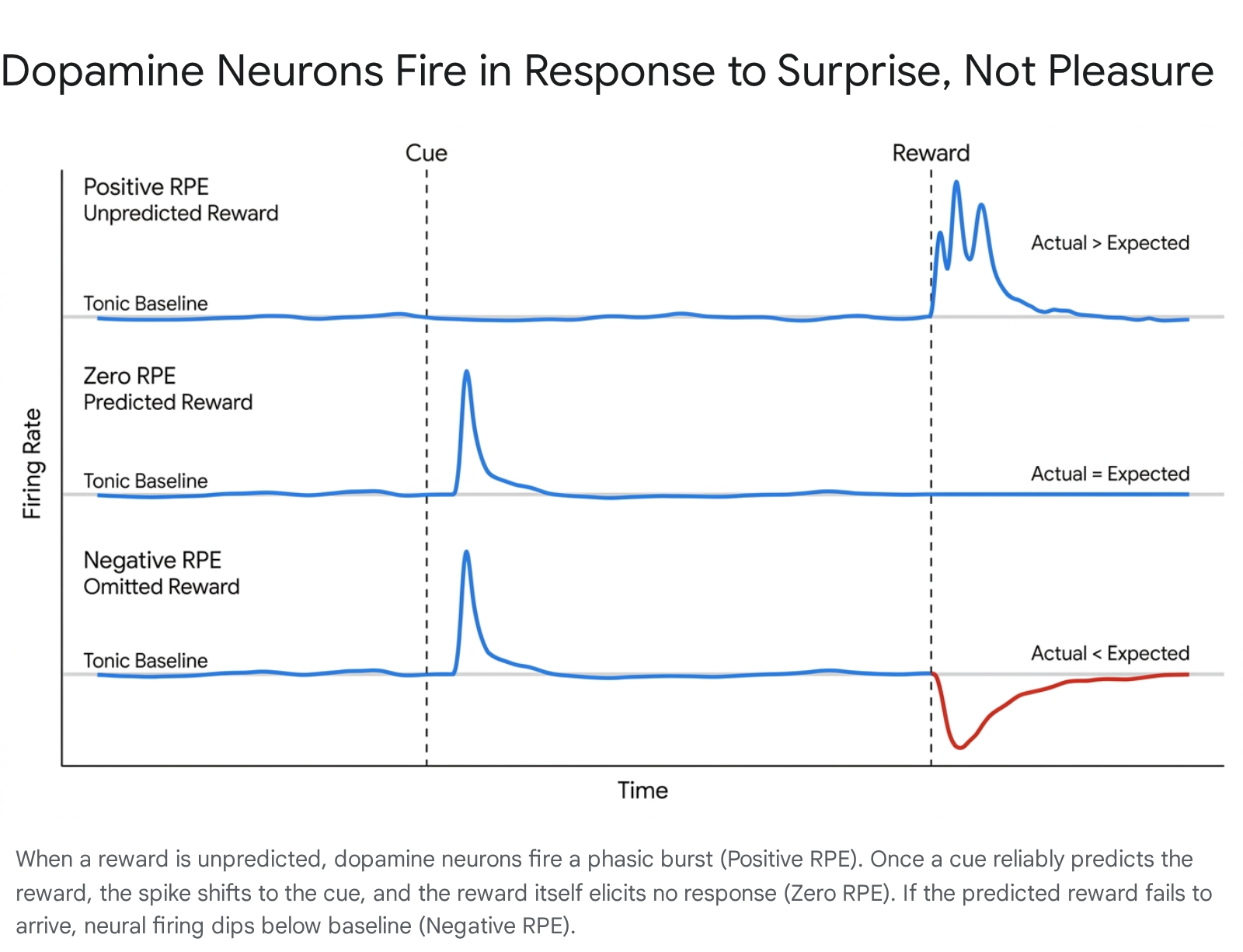
The dopamine system is continuously broadcasting updates to the rest of the brain based on the RPE equation. It operates on two distinct timelines. "Tonic" dopamine refers to the ambient, baseline hum of dopamine in the brain, setting the overall tone for motivation and mood 13. "Phasic" dopamine refers to the rapid, sharp bursts of neural activity that occur in response to specific events. This phasic firing operates in three distinct mathematical states 29.
| RPE State | Mathematical Condition | Dopamine Firing Response | Biological and Behavioral Output |
|---|---|---|---|
| Positive RPE | Actual Reward > Expected Reward | Sharp, rapid spike above tonic baseline | Strengthens synaptic connections; acts as a "teaching signal" driving the organism to repeat the preceding action. |
| Zero RPE | Actual Reward = Expected Reward | Remains steady at tonic baseline | Maintains the current behavioral model; signals that the environment is fully predicted and no new learning is required. |
| Negative RPE | Actual Reward < Expected Reward | Transient pause or dip below tonic baseline | Weakens synaptic connections; signals the organism to abandon the current strategy and update its internal model. |
When you experience a Positive RPE, the brain learns that the world just delivered more value than it promised. This pleasant surprise acts as an instructional spotlight, telling the brain to pay close attention to whatever happened just before the reward so it can be replicated 1614. If the outcome perfectly matches expectations, learning ceases because the internal model is already perfectly calibrated to reality.
The Mathematics of Disappointment
While positive prediction errors drive us forward, negative prediction errors are equally vital for survival. When an expected reward fails to materialize - or when an outcome is worse than anticipated - dopamine neurons briefly pause their firing, dropping below their baseline tonic rate 88. This drop is incredibly fast, typically lasting only 100 to 200 milliseconds, and can involve a near-complete cessation of neural activity 8.
This brief silence functions as the brain's ultimate disappointment signal. A negative RPE acts to actively weaken the synaptic connections that led to the failed outcome 8. It is the biological mechanism for updating a flawed mental model and unlearning a useless behavior 415. If you open a cabinet expecting to find your favorite snack and it is empty, the resulting drop in dopamine is how your brain learns not to check that specific cabinet again tomorrow. The brain efficiently calculates that the expectation model has collapsed, triggering a withdrawal of motivation: whatever just happened, do less of it 4816.
The Neurobiological Circuitry of Expectation
To compute the precise difference between an expectation and a reality, the brain relies on a highly sophisticated anatomical network centered in the midbrain. The primary sources of these dopamine signals are the ventral tegmental area (VTA) and the substantia nigra pars compacta (SNc) 817. These regions do not act in isolation; they integrate a massive influx of data from sensory, motor, and cognitive centers to generate the final RPE broadcast.
When a primary reward is delivered unexpectedly, excitatory glutamatergic inputs from areas like the lateral hypothalamus (LH) and the pedunculopontine tegmental nucleus (PPTN) feed into the midbrain 8. These excitatory signals force the dopamine neurons to fire rapidly, creating the phasic burst that we experience as a positive RPE. However, to function as an error detector, the brain must also have a way to cancel this excitatory burst when a reward is already expected.
How the Brain Cancels Surprises
This cancelation is achieved through a beautifully orchestrated mechanism of delayed synaptic inhibition. When an organism learns that a specific cue (like the ring of a bell or a push notification on a phone) predicts a reward, specific pathways in the striatum are activated. D1 receptor-expressing medium spiny neurons (MSNs) are triggered by the cue and project to GABAergic inhibitory cells that connect back to the midbrain dopamine neurons 810.
The timing of this inhibitory circuit is impeccable. The inhibitory signal acts like a delayed fuse, timed to peak at the exact millisecond the reward is historically scheduled to arrive 8.
If the reward arrives exactly as expected, the excitatory input of the reward and the delayed inhibitory input of the expectation perfectly collide. They cancel each other out entirely, resulting in a flat baseline firing rate - a Zero RPE 8. But if the reward does not arrive, the excitatory signal never fires. The unchecked inhibitory signal, however, still arrives right on schedule, suppressing the dopamine neurons below their baseline and creating the transient silence of the Negative RPE 8.
Dopamine Ramping and the Movement Debate
While the reward prediction error hypothesis is the most successful and widely accepted theory of dopamine function, it is not without controversy. In recent years, researchers have utilized advanced recording technologies, such as imaging mice navigating complex virtual reality corridors, which have revealed firing patterns that challenge the simplest interpretations of RPE 101819.
One of the most intensely debated phenomena is "dopamine ramping." As animals physically move closer to a distant goal in spatial navigation tasks, their dopamine levels slowly and continuously ramp up 17. At first glance, this gradual climb looks more like a signal of increasing "value" or pure motivation rather than a discrete calculation of mathematical error 1018.
However, neurocomputational theorists argue that ramping does not necessarily invalidate RPE. Some models suggest that if the internal representation of value is "convex" - meaning it curves sharply upward as the animal gets very close to the goal - the continuous sequence of prediction errors generated at every step would look exactly like a smooth ramp 10. Another prevailing theory, the "value decay model," posits that ramping results from state uncertainty; as an animal gets closer to a goal, sensory feedback becomes more informative, generating continuous mini-updates to the prediction error 10.
Real-Time Control Versus Learning
Another ongoing debate centers on whether dopamine acts strictly as a retrospective learning signal (updating values for future behavior) or as a real-time controller of movement and motivation (dictating current behavior). Acute optogenetic experiments - where researchers use light to artificially stimulate or silence specific neurons in awake animals - show that manipulating dopamine can alter movement choice, speed, and initiation in real time 10.
Some researchers argue that dopamine conveys "perceived saliency," which is a combination of how intense a stimulus is and how much attention it requires, regardless of whether it is good or bad. This theory attempts to explain why certain dopamine neurons occasionally fire in response to aversive stimuli, like foot shocks or loud noises, which a pure reward-based prediction error model struggles to neatly accommodate 1018. While the RPE framework remains dominant, these nuances suggest that the dopamine system may be multiplexing several different types of information - reward, movement kinematics, and sensory surprise - simultaneously 1017.
Beyond Averages: Distributional Reinforcement Learning
For over two decades, the standard temporal difference learning model operated on a simplifying assumption: it presumed that dopamine neurons encoded a single scalar value representing the expected average of all future rewards 202421. If a situation could yield either zero dollars or ten dollars with equal probability, traditional models assumed the dopamine system simply expected an average of five dollars.
However, real-world environments are rarely predictable enough for simple averages to be useful. Relying solely on the mean discards critical survival information about the variance, extremes, and risk profile of a given situation 22. In 2020, researchers at DeepMind, working at the intersection of artificial intelligence and neuroscience, proposed a radical update to the theory, which was subsequently validated in live animal recordings 20242123. The brain does not calculate a single average; it utilizes Distributional Reinforcement Learning.
Rather than all dopamine neurons acting uniformly as a choir to report the exact same mean prediction error, researchers found immense functional diversity among individual dopamine cells in the ventral tegmental area 242122. Some neurons act "optimistically"; they require a massive reward to trigger a positive RPE and are easily disappointed. Other neurons act "pessimistically," maintaining low thresholds and frequently firing positive RPEs even for minor rewards 17.
Collectively, this diverse population of differently tuned neurons allows the brain to encode the full probability distribution of potential future rewards 2122. By effectively representing multiple future outcomes simultaneously and in parallel, the brain captures the entire shape of risk and reward. Recent studies in 2024 and 2025 have further expanded this concept. Researchers have identified that distributional learning mechanisms are also ubiquitous in the prefrontal cortex, which handles higher-order decision-making 23. Furthermore, an advanced model called Time-Magnitude Reinforcement Learning (TMRL) suggests that dopamine neurons also encode the joint distribution of both the magnitude of the reward and the specific time delay before it arrives, creating a rich, multidimensional map of the future 22.
Action Prediction Error and Habit Formation
The RPE model elegantly explains how we learn what is valuable. However, it historically struggled to answer a fundamental paradox of human behavior: Why do we keep doing things that we know are no longer rewarding?
If you walk into a dark room where you know the lightbulb is burned out, you will likely still reach up and flick the switch. You do this without thinking, even though you know it will not produce the reward of light. According to strict RPE theory, a behavior that yields no reward should generate a negative prediction error and rapidly extinguish. Yet, human habits stubbornly persist long after the reward has vanished 24.
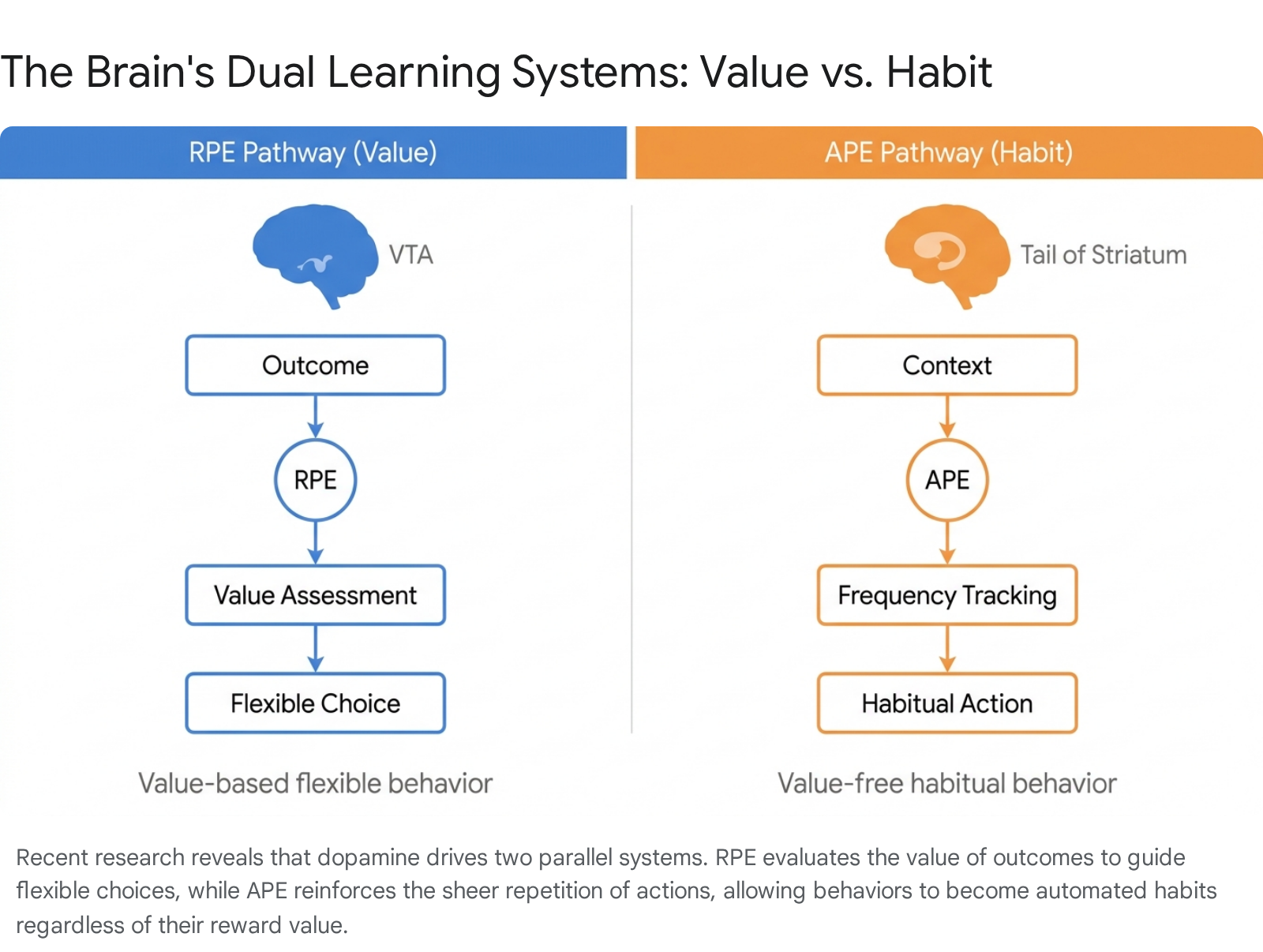
In May 2025, a landmark study published in the journal Nature by researchers at the Sainsbury Wellcome Centre at University College London (UCL) solved this paradox by identifying a parallel, second learning system in the brain 24252627. The researchers discovered that alongside Reward Prediction Error, the mammalian brain simultaneously utilizes an Action Prediction Error (APE) 25262829.
While RPE calculates the value of an outcome ("was that better than expected?"), APE completely ignores the reward. Instead, APE tracks the sheer statistical frequency of an action ("how often do I normally do this in this exact situation?") 242730.
The Dual-System Architecture
To prove the existence of this second system, researchers trained mice on auditory discrimination tasks and mapped the precise anatomical pathways involved. They discovered that the brain's dual learning systems are physically separated into different neural circuits 252935.
| Learning System | Biological Signal | Primary Brain Region | Core Function | Behavioral Result |
|---|---|---|---|---|
| Reward Prediction Error (RPE) | Value-based evaluation | Ventral Tegmental Area (VTA), Nucleus Accumbens | Tracks surprise based on outcome quality. Asks: Is this better than expected? | Drives goal-directed behavior, motivation, and flexible decision-making. |
| Action Prediction Error (APE) | Value-free frequency tracking | Tail of the Striatum (TS) | Tracks the repetition of an action in a specific context. Asks: Is this what I usually do? | Consolidates automated routines, habit formation, and compulsive repetition. |
When an animal is in a familiar context, the APE system updates the tendency to repeat whatever action was taken previously, acting as a value-free teaching signal 293035. Lead author Dr. Marcus Stephenson-Jones uses a sandwich shop analogy to explain the interaction: The first few times you visit a new shop, you use your RPE value-based system to carefully evaluate the menu, judging which sandwich tastes the best. However, after your fiftieth visit, you no longer expend cognitive energy evaluating the choices; you simply order your usual 2628.
Once a preference is initially established via the RPE value system, the APE system takes over, bypassing the brain's evaluative centers and turning the behavior into an automated default policy 242627. This dual architecture is highly efficient, freeing up cognitive resources for other complex decisions. However, it also explains why bad habits and compulsions are notoriously difficult to break. Interventions that solely focus on removing the reward often fail because the behavior is now being reinforced by the APE system simply through the act of repetition 2425.
Implications for Parkinson's Disease
The discovery of the Action Prediction Error system has immediate clinical implications, particularly for Parkinson's disease. Parkinson's is characterized by the progressive death of midbrain dopamine neurons. Clinicians have long observed a phenomenon known as "paradoxical movement," where patients who struggle to initiate habitual movements like walking can suddenly ride a bicycle or catch a thrown ball with ease 2627.
The APE framework provides a neurological explanation: the specific movement-related dopamine neurons in the tail of the striatum that die off in Parkinson's are the ones that drive the habitual APE system. Consequently, movements that rely on automated, habitual circuitry are severely compromised, while movements that rely on the flexible, value-based RPE circuitry remain largely intact 2627. This distinction opens new avenues for therapeutic interventions targeting specific dopaminergic circuits.
Observational Learning and Threat Omission
While dopamine is heavily associated with rewards we experience firsthand, recent research demonstrates that prediction error systems are also vital for observational learning and managing fear.
A 2025 study from the National Institute for Physiological Sciences (NIPS) in Japan explored how primates learn by watching others. By studying macaques, researchers identified that neurons in the dorsal part of the medial prefrontal cortex (DMPFC) are dedicated to encoding "Other-Reward Prediction Errors" (O-RPE) 31. These neurons fire when a primate observes a peer receiving an unexpected reward or facing a surprising disappointment. Remarkably, the brain processes the S-RPE (Self-RPE) and O-RPE simultaneously in parallel but distinct neural populations 31. This ability to vicariously process prediction errors is believed to be the neurobiological foundation of "mentalizing" - the cognitive capacity to infer the beliefs, motivations, and emotional states of others, serving as a critical mechanism for social learning and empathy 3137.
Prediction error logic also governs how we learn about safety. In the context of fear conditioning, the unexpected absence of a predicted threat - known as a threat omission prediction error - triggers a learning response that is virtually identical to a positive reward prediction error 32. When an anticipated pain or danger fails to occur, humans experience a wave of pleasant relief. Neuroimaging studies confirm that this relief triggers activations within the exact same reward and salience pathways (such as the ventromedial prefrontal cortex and nucleus accumbens) as finding unexpected food or money 32. The brain treats the absence of expected pain as a highly valuable reward, updating its models to designate the environment as safe.
Addiction and the Hijacking of the Prediction System
Understanding the precise mathematics of reward prediction error fundamentally demystifies how drugs of abuse and supernormal digital stimuli trap the human brain.
Under normal evolutionary conditions, the RPE system works flawlessly as a self-correcting thermostat. Hunger motivates food-seeking, the unexpected discovery of a high-calorie food generates a positive RPE, and the successful behavior is reinforced 114. Over time, as the organism learns the location of the food source and it becomes entirely predictable, the dopamine spike neutralizes to baseline 48. Learning is complete, and the organism is no longer obsessively driven by surprise.
Addictive substances bypass this delicate learning calibration entirely. Drugs like cocaine physically force the release of dopamine from the presynaptic neuron or aggressively block its reuptake in the synaptic cleft, generating an artificially massive, unearned positive prediction error 933.
Because the chemical artificially induces the dopamine spike directly at the receptor level, it bypasses the brain's internal expectation models. The brain interprets the drug event as an endlessly surprising, infinitely valuable outcome. The prediction model can never properly adjust to a "Zero RPE" state because, no matter how much the user consciously expects the high, the chemical forces the error signal to stay positive upon consumption 9. To the brain's learning algorithms, the drug appears to be the most critical survival resource ever encountered.
Receptor Downregulation and Tolerance
Faced with these artificially massive dopamine surges, the brain attempts to protect its delicate neural architecture via homeostasis. To muffle the overwhelming noise, the brain initiates a structural change: it physically reduces the density of dopamine receptors (specifically D2 receptors) in the reward circuitry 5103441.
This downregulation creates a devastating dual crisis for the addicted brain: 1. Tolerance: Because there are now fewer receptors available to process the dopamine signal, the exact same dose of the substance produces a much smaller relative prediction error 81341. The user must consume increasingly larger quantities of the substance simply to generate the teaching signal that once occurred naturally. 2. Anhedonia and Negative RPEs: The individual's baseline expectation has been shifted violently upward by the drug, while their receptor sensitivity has plummeted 58. Natural, everyday rewards - such as a good meal, completing a task, or seeing a friend - which used to provide mild, healthy positive RPEs, now pale in comparison to the skewed internal model. Because reality can no longer meet the new, chemically altered expectation, normal life generates a constant, exhausting stream of negative reward prediction errors. The brain calculates that everything in normal life is worse than expected, leading to profound disappointment, apathy, and anhedonia 58.
Functional magnetic resonance imaging (fMRI) studies on individuals with chronic cocaine addiction clearly illustrate this deficit. When addicted individuals participate in standard decision-making tasks involving monetary rewards, they show significantly weakened prediction error responses in the ventral striatum and orbitofrontal cortex compared to healthy controls 33. The addiction has physically blunted the brain's ability to process normal rewards and make flexible, value-based decisions.
The Neuroscience of Dopamine Fasting
As public awareness of dopamine's role in digital addiction, social media scrolling, and compulsive behaviors has grown, the trend of "dopamine fasting" has proliferated across the internet. Proponents often advocate for drastic measures: sitting in dark rooms, fasting from all food, avoiding screens, and shunning human contact for 24 hours in an attempt to "reset" brain chemistry 1335.
From a strict neurobiological standpoint, a literal "dopamine detox" is impossible and reflects a fundamental misunderstanding of the system. Dopamine is not a toxic substance that builds up and needs to be flushed out. It is a critical neuromodulator that the brain produces continuously to manage motor control, baseline motivation, and executive functioning 541.
The original protocol, conceptualized by psychiatrist Dr. Cameron Sepah, was never about depleting dopamine. It was rooted in evidence-based cognitive behavioral therapy (CBT) designed to reduce compulsive engagement with specific, highly stimulating behaviors - such as pathological gaming, mindless scrolling, or emotional eating 134135. The goal was behavioral modification and stimulus control, not neurochemical manipulation via monastic sensory deprivation 13.
Rebuilding Receptor Sensitivity
When individuals report feeling better after modifying their digital habits, they are not experiencing a reduction in dopamine; they are experiencing a slow recovery of their RPE baseline. However, neuroscientists and clinicians stress that a 24-hour break accomplishes virtually nothing on a structural level 541.
A genuine neurobiological "reset" requires the recovery of dopamine receptor sensitivity. Receptor upregulation is a slow, structural biological process, akin to building muscle. By deliberately withdrawing from a specific, chronically dysregulating stimulus, the brain is gradually forced to adapt to a lower-stimulation environment, slowly increasing D2 receptor density over time 541.
Clinical literature consistently indicates that meaningful recalibration of the brain's reward architecture requires a minimum of two to four weeks of sustained behavioral change, with substantial recovery often requiring 60 to 90 days 541. Furthermore, high-quality slow-wave sleep is non-negotiable during this period, as the brain physically consolidates these structural neuroplastic adaptations during deep rest 41.
Managing Expectations for Better Learning
The mathematics of reward prediction error offer a highly practical, evidence-based framework for managing daily motivation, emotional well-being, and habit formation. Because the brain measures satisfaction purely as the delta between an expectation and reality, manipulating your own expectations directly alters your neurochemical and emotional outcomes 1436.
If you set impossibly high, rigid expectations for a project, a relationship, or a vacation, you are neurologically pre-programming a negative prediction error. Even if the event goes reasonably well, reality is unlikely to surpass the artificially inflated prediction 3637. The resulting dip in dopamine will cause you to feel frustrated and unmotivated, even in the face of objective success. Some psychologists suggest that self-sabotage and the "fear of success" may actually be a subconscious defense mechanism: an attempt to align expectations with a known reality to avoid the devastating sting of a massive negative prediction error 38.
Conversely, intentionally adopting modest, realistic expectations mathematically ensures that average outcomes generate positive RPEs 36. This is not merely a trick of psychological framing or forced pessimism; it is an active manipulation of the midbrain's teaching signals 3637. When you expect less, you create the mathematical headroom required for reality to surprise you.
This mechanism perfectly explains the efficacy of "micro-goals." Setting a massive goal to run a marathon in three months creates an expectation model that is difficult to satisfy daily, resulting in a string of negative RPEs that quickly crush motivation 1639. By setting a micro-goal to simply put on running shoes and jog to the end of the street, success easily exceeds the minimal expectation. The resulting positive RPE releases a burst of dopamine that reinforces the behavior, linking the associative cues to the action 1639. Over time, this repeated RPE reinforcement hands the baton over to the Action Prediction Error system in the tail of the striatum, forging a permanent, automated habit 1629.
Bottom line
Reward prediction error is the foundational mathematical algorithm the brain uses to align its internal models with reality. By firing dopamine when we are pleasantly surprised and pausing it when we are disappointed, the midbrain continuously shapes what we value and how we act. Recent neuroscientific breakthroughs, such as the discovery of Action Prediction Error and distributional reinforcement learning, reveal that our brains seamlessly shift from calculating the value of flexible choices to automating them into rigid habits. While modern supernormal stimuli and addictive substances can easily hijack this system by skewing our baseline expectations, understanding the mechanics of prediction error allows us to consciously leverage realistic goal-setting to rebuild motivation and break destructive loops.