Working memory capacity and human intelligence
Introduction to Working Memory Systems
Working memory operates as the foundational mental workspace of human cognition, providing the architecture required for the temporary retention, active maintenance, and goal-directed manipulation of information. It acts as the critical interface between sensory perception and long-term memory, facilitating complex cognitive phenomena such as language comprehension, mathematical reasoning, decision-making, and fluid intelligence 123. Unlike historical models that conceptualized temporary storage as a passive waystation preceding permanent memory consolidation, modern cognitive science recognizes working memory as a highly dynamic, capacity-limited executive control system embedded within distributed neural networks 45.
The theoretical distinction between short-term memory and working memory has undergone extensive debate, though a contemporary consensus differentiates the two primarily by the presence of active manipulation 167. Short-term memory generally describes the brief, passive maintenance of information, such as retaining a string of digits in sequential order for a matter of seconds. Working memory encompasses both the transient storage of that information and the concurrent executive processes required to transform, update, or shield the data from internal and external interference 13. This active manipulation necessitates robust attentional control, forming the basis of goal-directed behavior 68.
Because working memory capacity is strictly limited, its operational bottlenecks provide vital insights into the fundamental limits of human intelligence. The constraints governing how much information can be simultaneously maintained and manipulated correlate strongly with general fluid intelligence - the ability to reason abstractly and solve novel problems independent of acquired, crystallized knowledge 6910. Consequently, understanding the structural models of working memory, the nature of its capacity limits, and its neurobiological substrates is central to mapping the parameters of human cognitive ability.
Structural Models of Working Memory
Over the past several decades, cognitive psychology has generated multiple theoretical frameworks to explain the structural organization and operational mechanics of working memory. These models attempt to chart how distinct cognitive systems interact to produce temporary information retention and executive control.
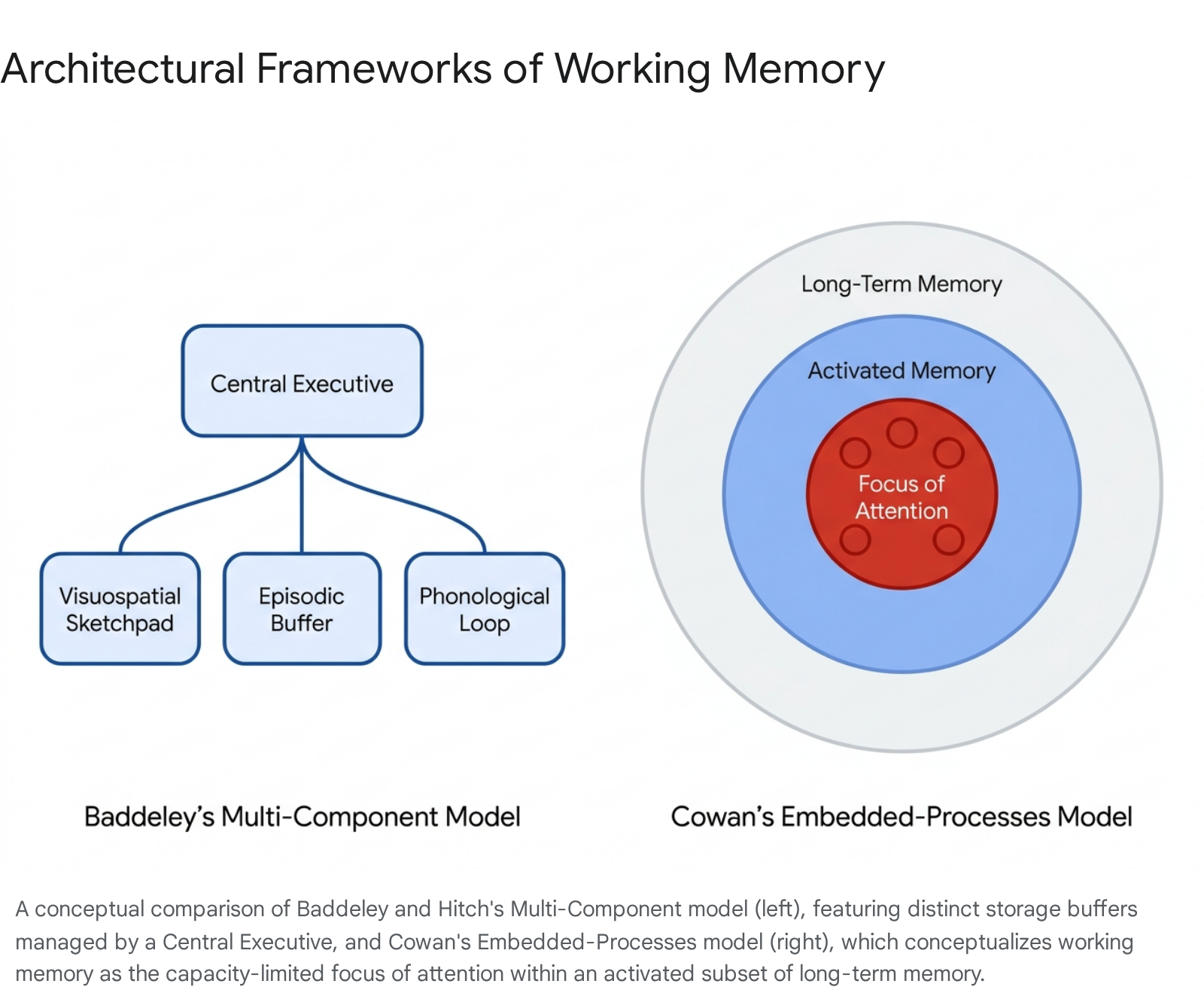
The Multi-Component Architecture
The most widely cited framework in cognitive literature is the multi-component model proposed by Alan Baddeley and Graham Hitch in 1974 311. Originally developed as an alternative to the unitary short-term store detailed in Atkinson and Shiffrin's 1968 modal model, Baddeley and Hitch divided working memory into distinct, domain-specific storage systems governed by a central controller 2812.
The multi-component model is constructed from four primary elements: The central executive functions as the core supervisory system. It is responsible for coordinating the subsidiary slave systems, allocating attentional resources, shifting between tasks, and overriding prepotent responses 2311. The central executive does not possess independent storage capacity; rather, it dictates how information within the subsidiary systems is processed and maintained.
The phonological loop is a specialized sub-system for maintaining verbal and acoustic information. It consists of a passive "inner ear" that holds auditory traces for one to two seconds, coupled with an active articulatory rehearsal process ("inner voice") that prevents the decay of these traces through subvocal repetition 2.
The visuospatial sketchpad serves as a parallel sub-system dedicated to visual and spatial data. It enables individuals to maintain and manipulate mental images, object features such as color and shape, and spatial maps 311. Early criticisms of this component argued for the separation of visual and spatial elements, noting that spatially aware but congenitally blind individuals lack prior visual input, yet maintain robust spatial working memory 2.
The episodic buffer was integrated into the model in 2000 to address theoretical gaps in how disparate information types are synchronized. This limited-capacity component acts as a multi-dimensional workspace that integrates phonological, visual, and spatial representations with chronological sequencing and long-term memory, forming cohesive, multi-modal episodic representations 21112.
Embedded-Processes Framework
An alternative theoretical approach eschews modular, domain-specific buffers, conceptualizing working memory instead as an activated state of long-term memory 2. The most prominent iteration of this approach is Nelson Cowan's embedded-processes model 812.
In Cowan's framework, working memory is not a separate structural compartment. It consists of neural representations located in long-term memory that have been temporarily activated above a baseline resting threshold. Within this broad field of activated memory, a narrower "focus of attention" exists. This focal area is strictly capacity-limited, typically capable of holding only about four individual chunks of information simultaneously 6812.
This model emphasizes central attentional capacity over domain-specific storage, suggesting that working memory limits are fundamentally constraints on executive attention rather than architectural storage space 913. By viewing memory as a continuous system, the embedded-processes model elegantly accounts for how prior knowledge effortlessly influences immediate recall, an effect more cumbersome to explain via isolated storage buffers 1213.
Structural Model Comparisons
| Theoretical Feature | Baddeley & Hitch Multi-Component Model | Cowan Embedded-Processes Model |
|---|---|---|
| Core Architecture | Distinct, dedicated sensory storage buffers managed by an executive controller. | Working memory is the actively attended portion of continuous long-term memory. |
| Source of Capacity Limits | Structural constraints within specific buffers (e.g., temporal decay of phonological traces). | Executive constraints on the central "focus of attention" (limited to ~4 distinct chunks). |
| Long-Term Memory Integration | LTM is a separate system accessed indirectly via the Episodic Buffer and Central Executive. | LTM is a continuous system; working memory exists inherently within the LTM architecture. |
| Domain Specificity | Highly domain-specific operations with segregated verbal and visual-spatial stores. | Emphasizes domain-general central capacity, though recognizes varying sensory activations. |
Theoretical Debates on Capacity Limits
One of the most intensely contested subjects in cognitive psychology involves the precise nature of working memory capacity, particularly regarding visual working memory. While researchers universally acknowledge that visual working memory is severely restricted, the mathematical and psychological definitions of this limit remain polarized between discrete slot theories and continuous resource theories 141516.
The Discrete Slot Hypothesis
The discrete slot model posits that visual working memory is composed of a fixed number of quantifiable storage locations, or "slots," typically estimated at three to four 141516. When the number of visual targets in the environment is less than or equal to the number of available slots, each target object is stored with high, relatively fixed precision.
When environmental demands exceed this capacity limit - such as viewing a display of six distinct colored squares - the discrete slot model predicts a strict "all-or-none" retention dynamic 1517. The cognitive system stores three or four items with high precision but retains absolutely no information regarding the remaining items. During continuous report tasks, where participants must select a remembered color on a continuous 360-degree color wheel, the slot model hypothesizes that response errors reflect a probabilistic mixture of two distinct distributions. Items successfully stored in a slot generate a narrow Gaussian distribution centered on the correct target, while items that did not receive a slot result in a uniform distribution spread equally across the entire color wheel, representing pure random guessing 1819.
Response-time modeling also supports elements of the discrete slot framework. Mixed-state discrete-slots models theorize that response times arise from a probabilistic mixture of memory-based evidence accumulation (occurring when a probed item was successfully stored in a slot) and guessing-based accumulation (occurring when the item was missed) 17. The ability of discrete-slots models to provide strong quantitative fits to response-time distributions suggests an abrupt threshold representing discrete item limits remains a plausible architectural constraint 1617.
The Continuous Resource Framework
In direct opposition, the continuous resource model rejects the notion of a fixed upper boundary on the absolute number of items that can be stored 1419. This framework proposes that working memory relies on a central, fluid pool of representational resources that can be continuously divided among an unlimited number of items 15.
Under the continuous resource model, an observer can theoretically store six, eight, or even ten items, but at the cost of precision 1418. As the number of encoded items increases, the allocation of resources per item decreases, resulting in increasingly noisy, low-resolution memory representations. When tested on larger set sizes, the continuous resource model predicts that observers do not resort to uniform, random guessing. Instead, they possess highly degraded, imperfect information regarding all presented items 1416.
Proponents of the resource model point to empirical data demonstrating that memory precision declines monotonically as set sizes increase. This phenomenon poses a significant challenge for pure discrete slot models, which can only account for declining precision by assuming that multiple discrete slots can be flexibly assigned to a single high-priority object when set sizes are small 151819. To reconcile these conflicting datasets, hybrid models - often termed "slots plus resources" - have been proposed. These hybrid frameworks suggest that working memory possesses a fixed upper limit on the number of discrete objects, but the resolution within those fixed slots is dictated by a continuously allocatable resource 151617.
Meaningful Stimuli and Capacity Expansion
Recent neurophysiological research adds further nuance to the capacity debate by demonstrating that the semantic meaning of a stimulus directly impacts apparent storage limits. Traditional visual working memory experiments have relied on abstract, meaningless stimuli, such as colored circles or oriented lines, to isolate purely perceptual capacity limits 20.
However, when subjects are tasked with remembering real-world, meaningful objects, behavioral capacity estimates expand significantly. Electroencephalography (EEG) recordings measuring contralateral delay activity (CDA) - a neural marker strongly associated with visual working memory capacity - show reliable increases in neural activity amplitude for meaningful and familiar stimuli compared to abstract stimuli 20. This expanded capacity indicates that visual working memory is not an isolated, low-level perceptual bottleneck. Instead, it interacts deeply with semantic knowledge, allowing familiar visual inputs to leverage established long-term memory representations to optimize limited active storage space 20.
Neural Substrates of Information Maintenance
The cognitive phenomena associated with working memory are instantiated by complex, distributed neural networks. Neurophysiological research has progressively shifted from viewing working memory as localized predominantly within the prefrontal cortex to understanding it as a dynamic, oscillatory interaction across widespread cortical and subcortical regions.
Persistent Neuronal Activity and the Prefrontal Cortex
Historically, the prefrontal cortex (PFC), specifically the dorsolateral prefrontal cortex (dlPFC), has been identified as the primary biological node for working memory. Early in vivo single-cell recordings in non-human primates established the paradigm of "persistent activity." This refers to the phenomenon wherein specific PFC neurons fire continuously during the delay period of a memory task, even after the sensory stimulus has been removed from the environment 212223.
This persistent delay-period firing, sustained by recurrent excitatory glutamate networks and regulated by lateral inhibition from GABAergic interneurons, was traditionally viewed as the direct neural correlate of information storage 322. Neurons located in the principal sulcus (Walker's area 46) exhibit highly selective spatial tuning, firing selectively when a subject maintains the memory of a stimulus in a specific spatial location 2224.
However, modern cellular analysis reveals greater complexity within the PFC. Research differentiating between WM-selective neurons (which are tuned to specific stimulus locations) and WM-sustained neurons (which maintain elevated activity regardless of the specific stimulus characteristics) shows that both neuron types exhibit similar spike variability, measured via Fano factor analysis 25. Furthermore, both types exhibit high Spike Phase Locking (SPL) in the alpha and beta frequency bands, coupling local field potentials to spike times. The presence of non-selective WM-sustained neurons indicates that the PFC's role extends beyond merely storing specific sensory features; it is heavily involved in maintaining abstract rules, cognitive control, and global task engagement 25.
Distributed Fronto-Parietal Networks
Current neurobiological consensus recognizes working memory as an emergent property of a distributed fronto-parietal network 212627. The dlPFC maintains dense reciprocal connections with the posterior parietal cortex (PPC). Both regions exhibit sustained activity during working memory tasks, and transcranial magnetic stimulation (TMS) applied to either area temporarily impairs task performance 3.
The operational division of labor between these regions suggests that the PPC is responsible for representing the prioritized sensory features of the retained items, while the PFC exercises top-down control to shield these representations from distraction and update them dynamically as task demands shift 2122. Functional magnetic resonance imaging (fMRI) combined with machine learning decoding has demonstrated that neural patterns in these regions are functionally interchangeable with those used for spatial attention. Classifiers trained to decode which item a subject is selectively attending to in their external visual field can successfully decode which item a subject is internally selecting within their working memory, strongly suggesting a shared neural mechanism for external perception and internal cognitive maintenance 28.
Oscillatory activity serves as the functional binding mechanism across this widespread network. Simultaneous EEG and fMRI recordings demonstrate that working memory encoding generates synchronized oscillatory activity in the theta frequency range (4 - 8 Hz). This theta synchrony effectively couples diverse regional networks - including the dlPFC, the supplementary motor area, the left dorsal premotor cortex, and various parietal regions - into a cohesive neural state necessary for converting transient sensory inputs into stable, maintainable constructs 2629.
Connectome-Based Connectivity and Pathologies
The importance of whole-brain connectivity in working memory function is further highlighted by connectome-based predictive modeling (CPM). Using resting-state and task-based fMRI, researchers can predict an individual's working memory capacity purely by analyzing the distributed functional connectivity patterns across their entire brain 3031. High-resolution diffusion-weighted imaging (DWI) reveals that structural white matter connectivity, particularly within sensorimotor, ventral attention, and subcortical networks, mediates age-related declines in working memory and fluid cognition 3233.
Pathological disruptions to these networks severely degrade working memory. For instance, individuals with Neurofibromatosis type 1 (NF1) - a neurodevelopmental disorder characterized by executive function deficits - exhibit altered functional connectivity during spatial working memory tasks 34. Relative to neurotypical controls, NF1 participants display reduced deactivation of the posterior cingulate cortex (PCC) and temporal regions, alongside increased, inefficient connectivity between visual, sensorimotor, and dorsal attention networks. This network reorganisation towards greater assortativity forces an accuracy-speed trade-off, demonstrating how optimal working memory requires precise, efficient network communication rather than merely generalized brain activation 34.
The Role of the Medial Temporal Lobe
While traditionally associated almost exclusively with long-term episodic memory, the medial temporal lobe (MTL) - including the hippocampus and parahippocampal gyrus - is increasingly recognized as a vital component of the working memory network. The MTL is particularly engaged during the maintenance of complex, highly detailed, or novel visuospatial configurations 2735.
The MTL facilitates "mnemonic discrimination" or pattern separation, a computational process allowing individuals to maintain high-resolution representations of objects and successfully resist interference from highly similar, but distinct, perceptual lures 363738. Lesions in the MTL impair the ability to discriminate between similar objects in working memory and severely degrade facial working memory capacity, confirming that the ventral pathway is critical for high-fidelity object maintenance 2737.
The Intersection of Working Memory and Fluid Intelligence
One of the most robust findings in psychometrics and differential psychology is the profound correlation between an individual's working memory capacity and their general fluid intelligence (gF) 61039. Fluid intelligence represents the ability to reason abstractly, identify complex patterns, and solve novel problems without relying on prior crystallized knowledge. In large-scale structural equation modeling, latent factors representing working memory capacity routinely correlate with fluid intelligence at magnitudes approaching $r = 0.80$ to $0.90$, and occasionally near unity 10.
Shared Variance and Interference Control
The near-isomorphic statistical relationship between working memory and fluid intelligence poses a critical neurocognitive question: what underlying operational mechanism do they share? Research indicates that this shared variance is not driven merely by passive storage volume, but rather by domain-general executive control mechanisms - specifically, the neural ability to control attention and suppress interference 394041.
In demanding cognitive environments, both fluid reasoning tasks and complex working memory paradigms require an individual to maintain focus on task-relevant information while actively inhibiting proactive and retroactive interference 642. Functional MRI studies utilizing the n-back task demonstrate that on high-interference "lure" trials (where a presented stimulus matches a recently seen item but is not the correct target), performance relies heavily on specific bilateral regions of the dlPFC (middle frontal gyrus; BA9) and parietal cortex (inferior parietal cortex; BA 40/7) 3940.
Crucially, path analyses demonstrate that the neural activity evoked by interference resolution statistically mediates the relationship between individual differences in working memory span and fluid intelligence scores. Interference-related performance accounts for a significant proportion of the shared variance between the two constructs. Thus, individuals with high gF are fundamentally those possessing highly efficient neural networks for interference control and attention allocation 39404142.
Physiological Biomarkers of Intelligence
Meta-analyses of neuroimaging data consistently support this shared mechanism, showing profound anatomical overlap between the neural regions activated during fluid intelligence tasks and executive updating tasks. Both gF and working memory updating preferentially recruit the left middle frontal gyrus, the left inferior parietal lobule, and the anterior cingulate cortex 4344.
This overlap extends to physiological biomarkers during sleep. Simultaneous EEG-fMRI recordings demonstrate that sleep spindles - bursts of neural oscillations between 11 and 16 Hz during non-rapid eye movement (NREM) sleep - are highly correlated with fluid intelligence. Spindle events trigger activations in the thalamus, bilateral putamen, medial frontal gyrus, and precuneus. The extent of this spindle-dependent brain activation correlates specifically with reasoning abilities (gF), but not with simple short-term memory or crystallized verbal abilities, positioning sleep spindles as a unique physiological marker of the neural hardware supporting complex problem-solving 45.
Furthermore, as individuals age, the trajectory of fluid intelligence is intrinsically linked to memory mechanisms. Multivariate longitudinal analyses reveal that age-related changes in cognitive abilities are largely domain-general; individuals who experience significant losses in fluid abilities simultaneously experience smaller gains or declines in crystallized abilities, reinforcing the presence of a generalized cognitive factor anchored in working memory efficiency 46.
Efficacy of Cognitive Training Programs
Given the robust predictive power of working memory capacity for academic achievement, professional success, and fluid intelligence, a substantial commercial and academic industry has emerged dedicated to cognitive "brain training" 474848. Computerized cognitive training programs, such as adaptive n-back tasks and complex span paradigms, assert that rigorous working memory practice can expand biological capacity limits and, by extension, enhance general fluid intelligence.
The scientific evaluation of these claims hinges on the distinction between two types of cognitive transfer: near transfer and far transfer.
Near Transfer Effects
Near transfer refers to improvements on untrained tasks that are structurally and cognitively highly similar to the trained tasks (e.g., training extensively on an auditory n-back task and demonstrating subsequent improvement on a visual n-back task or a related short-term memory assessment).
Extensive second-order meta-analyses - which aggregate the results of hundreds of randomized controlled trials encompassing tens of thousands of participants - demonstrate that working memory training reliably produces near transfer 4849. Participants become substantially more proficient at the trained tasks and exhibit measurable improvements in highly related working memory assessments. This suggests that sustained practice facilitates the development of specific cognitive strategies, task familiarity, and specialized mental routines, effectively optimizing the use of existing capacity for specific task constraints 49.
The Absence of Far Transfer
Far transfer, however, refers to improvements on tasks that rely on fundamentally different cognitive domains, such as untutored fluid intelligence tests, reading comprehension, mathematical reasoning, or general executive control. This is the ultimate goal of commercial brain training programs.
The meta-analytic consensus regarding far transfer is decidedly pessimistic 4750. Across meticulously controlled studies that account for baseline performance differences, placebo effects, and publication bias, the aggregate effect size of working memory training on fluid intelligence and broad academic skills converges squarely on zero 474850.
When positive far transfer effects are occasionally reported in isolated studies, they are almost universally observed in trials utilizing passive, untreated control groups 4751. When cognitive training interventions are compared against active control groups - participants engaged in a visually similar but non-adaptive computer task designed to account for placebo effects, expectation bias, and general computer familiarity - the alleged gains in fluid intelligence consistently disappear 475152.
Data indicates a stark contrast in aggregate effect sizes between transfer types. Near transfer effect sizes for trained and untrained working memory tasks range from a standardized mean difference (g) of roughly 0.20 to 0.42. Conversely, far transfer effect sizes for fluid intelligence, nonverbal ability, reading comprehension, and arithmetic hover near absolute zero (g ~ 0.00 to 0.05), demonstrating no statistically significant functional improvement.
Transfer Effects in Cognitive Training
| Transfer Category | Evaluated Cognitive Domain | Aggregate Effect Size (g / SMD) | Longevity of Effect |
|---|---|---|---|
| Near Transfer | Trained Working Memory Tasks | ~ 0.33 to 0.42 | Maintained at short-term follow-up. |
| Near Transfer | Untrained Working Memory Tasks | ~ 0.20 to 0.33 | Partially maintained; declines over long-term. |
| Far Transfer | Fluid Intelligence (gF) / Reasoning | ~ 0.00 to 0.05 (Null) | Not sustained; mostly observed against passive controls. |
| Far Transfer | Reading Comprehension & Word Decoding | ~ 0.00 (Null) | No long-term follow-up effect. |
| Far Transfer | Mathematical / Arithmetic Skills | ~ 0.00 (Null) | No robust generalized improvement. |
The implications of these meta-analytic findings are profound. They suggest that the tight correlation between working memory and fluid intelligence represents a structural, biological invariant of human cognition 5053. While an individual's efficiency in utilizing their available capacity can be optimized for a specific, practiced task, the underlying neural infrastructure - the raw biological capacity limit - cannot be fundamentally expanded through behavioral training regimens 4950.
Cross-Cultural Variability and Assessment Bias
A critical, yet historically under-examined, variable in the study of working memory and intelligence is the cultural context of the populations being researched. The vast majority of foundational cognitive models, psychometric capacity limits, and neuroimaging databases rely almost exclusively on samples drawn from Western, Educated, Industrialized, Rich, and Democratic (WEIRD) societies - primarily university undergraduates 545556.
Generalizability Across Populations
Relying heavily on WEIRD populations carries an implicit assumption that cognitive architecture is universally uniform regardless of environmental context. However, comparative cross-cultural psychology has demonstrated that members of WEIRD societies are frequently extreme statistical outliers in domains related to visual perception, spatial reasoning, fairness, and categorization 555658.
This demographic bias heavily impacts the applicability and fairness of neuropsychological assessments. Tools designed, normed, and validated in Western, English-speaking contexts frequently penalize culturally, linguistically, and educationally diverse populations, leading to severe misinterpretations of cognitive decline or baseline capacity deficits 5758. For example, structural equations modeling of working memory in non-WEIRD populations (such as multilingual, low-socioeconomic adults in South Africa) indicates that while the broad multi-component structure of working memory (separate verbal and visuospatial storage and processing) remains a valid baseline, the specific interrelationships between components and their correlation with fluid intelligence are heavily moderated by educational quality, bilingualism, and cultural reasoning styles 5459.
Cultural Modulation of Neural Encoding
Culture shapes not only behavioral performance metrics but also the specific neural mechanisms recruited during working memory encoding and recognition tasks. Cross-cultural neuroimaging (fMRI) studies have revealed distinct anatomical differences in how Western (e.g., American) and East Asian (e.g., Taiwanese) adults process visual working memory 363738.
Behaviorally, Western populations tend to exhibit highly analytic processing, focusing intensely on specific focal objects independently of their background. This cognitive style results in higher baseline memory specificity and better discrimination of novel perceptual lures. East Asian populations, conversely, tend to exhibit more holistic processing, prioritizing context and the relationships between objects in a visual field 3638.
Neurally, these cultural cognitive styles manifest in the recruitment of distinct brain regions during memory retrieval. When distinguishing similar perceptual lures from old targets, American subjects rely heavily on variations in left superior parietal cortex activity and exhibit a strong positive correlation between behavioral discriminability and activation in the right fusiform gyrus - a region associated with processing detailed visual features 3637. In contrast, Taiwanese populations exhibit a negative correlation in the fusiform gyrus and show different relational activations in the left inferior frontal gyrus (LIFG) and middle frontal gyrus for general object recognition 3638.
Such findings strongly indicate that while the baseline biological hardware of working memory is ubiquitous, lifelong cultural and environmental experiences functionally re-wire the specific neural pathways used to encode, maintain, and retrieve visual information 273637. Meta-analyses further suggest that the strength of the correlation between working memory capacity and elements of divergent thinking may be significantly stronger in Asian cultural contexts compared to Western ones, implying that the specific strategies used to manipulate memory are culturally contingent 60.
Consequently, advancing a truly comprehensive understanding of human cognitive architecture requires integrating non-WEIRD populations to differentiate biological invariants from culturally learned cognitive strategies.