Social complexity and the evolution of human intelligence
The evolutionary trajectory of the mammalian brain, particularly within the primate order, is characterized by exceptional encephalization. Brain tissue is metabolically expensive; the adult human brain accounts for approximately 2% of total body mass while consuming up to 20% of the body's total energy intake 1. Given the fundamental principles of evolutionary biology, which dictate that biological adaptations must confer fitness benefits sufficient to outweigh their metabolic costs, the persistent expansion of the primate neocortex requires a robust evolutionary explanation 1. Historically, the consensus view assumed that brains evolved primarily to process information of ecological relevance, enabling organisms to navigate environmental challenges. However, this ecological view often fails to explain why primates require significantly larger brains than other mammalian taxa to perform similar ecological tasks 1.
In response to these anomalies, comparative neuromorphology and behavioral ecology have increasingly focused on the complex structural dynamics of primate societies as the primary selective pressure driving neural expansion. This framework, formalized as the social brain hypothesis, asserts that the distinctively large brains of anthropoid primates evolved primarily to manage the computational demands of living in unusually complex, stable, and differentiated social groups 1234. Unlike the simpler aggregations seen in many vertebrate taxa, primate groups are held together by intricate networks of bonded relationships, coalitions, and dominance hierarchies, which require sophisticated socio-cognitive mechanisms to navigate 3567. Over the past three decades, the core premises of this hypothesis have been rigorously tested, expanded, and challenged, leading to adjacent evolutionary models and deep neuroanatomical investigations.
Evolutionary Context of Primate Brain Size
In foundational studies of brain evolution, researchers argued that brain size scales predictably with body size due to fundamental allometric relationships required to manage the physiological machinery of the body. The metric of interest is not absolute brain size, but rather the spare brain capacity over and above that needed for basic somatic management 1. Evolutionary changes in primate brain volume have been largely mosaic, meaning that the brain did not expand uniformly. Instead, specific components of the neural system grew at varying rates depending on localized selective pressures 14.
Within primates, these changes in brain size have predominantly involved the disproportionate growth of the neocortex. The neocortex is responsible for higher-order cognitive abilities, including reasoning, conscious thought, and complex social processing, and it accounts for 50% to 80% of total brain volume in primates 48. In insectivorous mammals, the neocortex is roughly the same size as the medulla, a primitive brain region controlling respiration and heart rate. In prosimians, the neocortex is ten times larger than the medulla; in monkeys and apes, it is 20 to 50 times larger; and in humans, the neocortex is 105 times larger than the medulla 4.
Neocortical Metrics and Group Size Modeling
To identify the selective pressures driving this mosaic expansion, researchers sought to map anatomical indices of cognitive capacity against environmental and social variables. Utilizing comprehensive neuroanatomical datasets, such as the widely cited morphological measurements compiled by Stephan et al. in 1981, comparative biologists tested numerous variables, including the percentage of fruit in the diet, the size of ranging areas, and the length of day journeys 79. The empirical results demonstrated that ecological variables were generally poor predictors of relative neocortical volume. Instead, the most robust predictor of neocortex size was social group size 9.
The Neocortex Ratio
To standardize comparisons across species with vastly different body masses and energetic requirements, researchers utilized the "neocortex ratio." This metric is calculated by dividing the volume of the neocortex by the volume of the rest of the brain (the total brain volume minus the neocortex) 457. This ratio effectively discounts the hindbrain and subcortical structures tied to basic physiological regulation, isolating the neural hardware available for executive processing.
When the mean social group size for various primate genera is plotted against the neocortex ratio, a strong positive correlation emerges. Analyses utilizing reduced major axis (RMA) regression yield the equation log(N) = 0.093 + 3.389 log(CR), where N represents mean group size and CR represents the neocortex ratio 7910. This equation accounts for approximately 76% of the variance in mean group size among 36 genera of prosimian and anthropoid primates 5710. The reliance on mean group size, rather than maximum observed group size, is intentional. Primate groups are highly structured, and maximum group size typically represents the threshold of social collapse and fragmentation rather than the stable capacity of the species 710.
Mathematical Bounds and Cognitive Constraints
This linear relationship on a logarithmic scale indicates that an individual organism's neocortical processing capacity acts as a strict upper constraint on the number of stable, interpersonal relationships it can simultaneously monitor and maintain 101112. When primate groups exceed the cognitive threshold dictated by their species-specific neocortex ratio, individuals suffer from information overload regarding the structure of relationships. The group experiences increased internal friction, fails to defuse conflicts arising from shared foraging spaces, and inevitably fragments into smaller, more manageable daughter cliques 7912.
Extrapolating this data to modern humans provides a theoretical calculation of human social capacity. The human neocortex volume is approximately 1006.5 cc against a total brain volume of 1251.8 cc, yielding a neocortex ratio of 4.1. This ratio is roughly 30% to 50% larger than the maximum value observed for any other primate species 5710. Interpolating a ratio of 4.1 into the ape-specific regression equation yields a predicted human group size of 147.8 10. Due to the logarithmic transformation and the extrapolation beyond the bounds of non-human primate data, the 95% confidence limits around this prediction are moderately wide, ranging from 100.2 to 231.1 individuals 710. Alternative indices of neocortex size, such as Jerison's Extra Neocortical Neurons Index or Encephalization Quotient residuals, yield predicted group sizes ranging from 107.6 to 189.1, all closely aligning with this core estimate 5710.
Socio-Cognitive Evolutionary Grades
Subsequent structural analyses of these regression models revealed that the relationship between neocortical volume and group size is not a single, continuous evolutionary pathway. Instead, the data partition into distinct evolutionary "grades," indicating progressive step-changes in the computational demands of sociality 813.
Data plotting neocortex ratios against mean social group sizes across primate clades reveals these distinct evolutionary shifts. Specifically, prosimians, monkeys, and apes align along parallel regression trendlines, demonstrating that these clades operate within distinct socio-cognitive grades. Extrapolating the human data point along the ape trendline - using a neocortex ratio of approximately 4.1 - yields a predicted group size near 150. These parallel slopes demonstrate that managing a group of a given numerical size requires proportionately more computational power as a lineage progresses from prosimians through monkeys to apes 1314.
K-Means Clustering of Primate Taxa
Advanced k-means cluster analyses partitioning the residual group sizes identify four primary socio-cognitive grades. The lowest grade (Grade IV) consists of platyrrhines and strepsirrhines that typically live in relatively small, monogamous pairs or solitary arrangements, with a mean group size of 3.2 8. Grades II and III contain monkeys that live in relatively small, stable groups, alongside apes such as Gorilla and Hylobates, averaging 17.4 and 9.0 individuals respectively 8.
The highest tier (Grade I) consists of genera that exhibit extensive fission-fusion flexibility, live in exceptionally large groups compared to related taxa, or form large, stable, multi-male/multi-female bonded groups. Genera in this grade include Ateles, Pan, Papio, and Macaca, with a mean group size of 34.9 8. Notably, chimpanzees (Pan) exhibit a bimodal distribution with community sizes peaking around 32 and 59 individuals, aligning closely with the upper limits of these sophisticated cognitive grades 8. This stratified structure indicates that ape sociality is fundamentally more cognitively demanding than monkey sociality. Apes require a substantially larger "neural computer" to manage a group of a specific size because their alliances rely on complex mentalizing, tracking third-party relationships, and managing fluid fission-fusion dynamics 131414.
Time Budgeting and Grooming Cliques
The absolute size of a primate group is heavily constrained by temporal and ecological realities. Large primate groups are not homogenous masses; they are constructed by welding together sets of smaller, tightly bonded grooming cliques 9. Social grooming serves as the primary mechanism for establishing trust, buffering individuals against social stresses, and cementing coalitions. Grooming triggers the release of endorphins, forming a psychopharmacological platform for social cohesion 61516.
However, maintaining relationships via physical grooming requires a massive time investment. Research demonstrates that the percentage of daylight hours devoted to social grooming scales linearly with group size 57. The highest recorded value for grooming time allocation in a single species is 18.9% for a group of Papio papio baboons, representing a practical upper limit for time expenditure 7. If a species were to attempt to maintain the stability of human-sized or exceptionally large ape-sized groups strictly through manual grooming, the time demands would become intolerable, severely compromising the organism's ability to forage, rest, and reproduce 5.
Sex-Specific Drivers of Primate Sociality
A critical refinement to the social brain hypothesis involves the bifurcation of selective pressures based on sex. While foundational studies aggregated species data without regard to sex, primate social evolution is heavily dictated by divergent reproductive demands between males and females 17.
Female reproductive success is primarily linked to the acquisition of nutritional resources and protection from predators, leading females to group according to conditions set by the physical environment. Conversely, male reproductive success is driven by monopolizing access to fertile females; males, therefore, distribute themselves socially in response to female grouping patterns 17. If relative neocortex size limits the number of social interactions an individual can track, this cognitive constraint should exert a stronger selective pressure on female group sizes, simply because female-female bonded networks form the stable core of most primate societies 17.
Phylogenetic comparative analyses of relative neocortex sizes against male and female group sizes validate this sex-specific hypothesis. Using comprehensive datasets (such as Isler et al., 2008), multiple regression models confirm that female group size is significantly and positively correlated with relative neocortex volume (p = 0.006) 17. In stark contrast, male group size shows no significant correlation (p = 0.275), and when analyzed collectively, exhibits a slight negative tendency 17. These findings indicate that brain evolution in primates proceeds primarily in accordance with the social demands of females, establishing that the social intelligence hypothesis is fundamentally a theory of female sociality 17.
The Human Cognitive Limit: Dunbar's Number
Extrapolating the constraints of the neocortex ratio to Homo sapiens yielded the widely cited "Dunbar's number," which proposes a cognitive limit of approximately 150 stable social relationships for modern humans 111618. A stable relationship is strictly defined as one wherein an individual knows the identity of the other person, understands how that person relates to the rest of the network, and shares a history of reciprocal trust, altruism, and obligation 1116.
Fractal Layers of Social Intimacy
Human social networks are not flat distributions; they exhibit a highly distinct, hierarchical fractal structure organized into concentric layers of intimacy. These layers scale by a relatively consistent factor of three, reflecting varying levels of emotional closeness, contact frequency, and willingness to behave altruistically 16192021.
The structural layers are defined as follows: * 1.5 to 5 individuals: The support clique. This innermost layer contains romantic partners and best friends who provide absolute emotional and practical support 161922. * 15 individuals: The sympathy group. This layer encompasses close friends and key confidants 1621. * 50 individuals: The affinity group. This layer consists of good friends and frequent interactants, commonly overlapping with regular social companions 1621. * 150 individuals: The active network. This limit encompasses "just friends" and extended family. It demarcates the boundary of personalized, reciprocal relationships 1621.
Beyond the 150-layer boundary, relationships undergo a qualitative shift. Interactions become increasingly transactional, rely on identity recognition rather than true intimacy, and are governed by restrictive rules, laws, and enforced norms rather than organic social bonding. These outer layers encompass 500 acquaintances, 1,500 familiar faces, and scale upwards to 5,000 known faces 111623.
Digital Networks and the Persistence of Constraints
The proliferation of global social media platforms raised the question of whether technological mediation could bypass biological cognitive constraints. If geographic dispersal and the time costs of physical communication were removed, could humans maintain stable relationships numbering in the thousands?
Analyses of user behavior across platforms such as Twitter and Facebook strongly suggest that biological limits persist in digital environments. Data-driven studies reveal that the vast majority of users maintain an active communication network that closely mirrors the 150 threshold 18202124. While a user may accumulate hundreds or thousands of passive connections or "followers," the volume of reciprocal communication - which requires the deployment of limited cognitive resources and attention - remains bounded by Dunbar's number. Students with an average of 300 Facebook connections, for instance, typically only consider roughly 75 to 150 of them as actual, meaningful friends 20.
Furthermore, psychological literature indicates an adverse effect to social overexposure. Operating digital networks that vastly exceed human cognitive boundaries without cultivating deep, proximal ties has been correlated with increased social dissatisfaction and a higher probability of exhibiting symptoms of depression 2324.
Methodological Critiques and Cross-Cultural Variance
The precision and universality of Dunbar's number are highly contested within certain sectors of evolutionary biology and sociology. Critics assert that human networks are governed by cultural and technological factors, and that relying on regression extrapolations from non-human primate datasets is statistically flawed 1226.
When researchers re-analyzed the foundational brain data using Bayesian and generalized least-squares (GLS) phylogenetic methods, the results yielded wildly different estimates. Bayesian models generated approximate average group sizes ranging from 69 to 109, while GLS models predicted sizes between 16 and 42 12. More critically, these alternative analyses produced enormous 95% confidence intervals (ranging from 4 to 520 individuals), leading critics to argue that deriving any single, specific cognitive limit on human group size is mathematically futile 12.
Additionally, cross-cultural studies reveal significant global variance in the composition of intimacy circles. While the overarching fractal structure remains somewhat consistent, the cultural perception of friendship differs deeply. In a survey of over 10,000 global participants, individuals in Saudi Arabia reported an average of 6.6 best friends, while individuals in the UK averaged 2.6, and a significant portion of Americans reported having only one or no best friends 22. These findings suggest that while the neocortex sets a hard ceiling on total tracking capacity, cultural frameworks deeply modulate the internal distribution of human social networks.
Competing and Complementary Evolutionary Frameworks
The assertion that social complexity is the sole or primary driver of primate encephalization is continuously weighed against other ecological and developmental selective pressures. If brain expansion demands massive caloric intake, the extra neural tissue must "pay for itself" energetically through enhanced survival or foraging efficiency 125. This has led to the formalization of alternative evolutionary frameworks.
| Evolutionary Framework | Primary Selective Pressure | Predicted Neurological Outcome | Core Mechanisms | Species Extrapolation |
|---|---|---|---|---|
| Social Brain Hypothesis | Group size and social complexity. | Expansion of neocortex, specifically DMN and mentalizing networks. | Coalition building, reciprocal altruism, tracking alliances, grooming cliques. | Explains the quantitative neocortex-group size link in primates; predicts Dunbar's number in humans. |
| Ecological Brain Hypothesis | Foraging efficiency, diet quality, and spatial navigation. | Expansion of domain-general intelligence, spatial memory, and motor integration. | Extractive foraging, mental mapping of ephemeral resources, overcoming intense seasonality. | Explains the shift from folivory to frugivory; emphasizes energetic benefits of increased caloric intake. |
| Cultural Brain Hypothesis | Accumulation of adaptive knowledge and social learning. | Enhancement of regions linked to imitation, information storage, and extended plasticity. | Autocatalytic feedback loop: social learning drives cultural complexity, demanding greater neural capacity. | Distinguishes hominins and great apes; links extended juvenile development to cultural acquisition. |
The Ecological and Environmental Brain Hypotheses
The ecological brain hypothesis proposes that the extraordinary evolutionary costs of large brains were offset by the enhanced cognitive ability to solve complex environmental problems. This includes extracting hidden food sources, predicting the fruiting cycles of trees, and navigating efficiently through vast, unpredictable home ranges 12526.
Phylogenetic comparative analyses incorporating dietary variables suggest that the shift toward high-quality, energy-dense diets - specifically the transition from leaf-eating (folivory) to fruit-eating (frugivory) - is a stronger predictor of relative brain size across primates than social group size 127. The necessity of building complex mental representations to forage ephemeral fruit requires substantial spatial memory and executive function 2628.
Recent neuroanatomical analyses mapping specific brain regions indicate a synthesis of social and ecological pressures. The frontal pole (FP), vital for metacognitive skills, and the dorsolateral prefrontal cortex (DLPFC), vital for working memory, scale positively with both population density (a proxy for social interaction) and daily traveled distance (a proxy for foraging complexity) 262829. This dual correlation suggests that primate brain evolution is a mosaic process where the cognitive machinery required to track migrating resources heavily overlaps with the circuitry required to track fluctuating social alliances 2829.
The Cultural Intelligence Hypothesis
A highly specialized extension of the social brain framework is the cultural intelligence hypothesis, alternatively framed as the Cumulative Cultural Brain Hypothesis. This theory asserts that human cognition, and the cognition of certain great apes, evolved specifically to acquire, store, and manage cultural knowledge via sophisticated social learning 303132.
The hypothesis posits an evolutionary autocatalytic feedback loop. Initial ecological pressures required individual problem-solving, generating a pool of adaptive non-genetic information within a population, such as tool use or extractive foraging techniques. As this cultural capital became essential for survival, natural selection favored individuals with the neural architecture necessary for high-fidelity social learning 313334.
Larger brains enabled superior social learning, which fostered the accumulation of even more complex adaptive information. This escalating complexity drove selection for even larger brains and heavily extended juvenile periods to provide offspring sufficient time to absorb the cultural knowledge 2731. Empirical developmental studies support this theory; human children demonstrate superior mentalizing abilities that are heavily associated with targeted cultural learning - the selective imitation of relevant actions - rather than merely utilizing social information to outcompete peers in zero-sum interactions, as predicted by pure "Machiavellian" intelligence models 34.
Taxonomic Anomalies and Non-Primate Brain Evolution
While the quantitative relationship between neocortex size and social group size is robust within anthropoid primates, attempts to generalize the social brain hypothesis across all vertebrate taxa have encountered severe limitations. Brain evolution is deeply contingent on the phylogenetic history of the clade, and diverse selection pressures trigger varied architectural responses.
Pair-Bonding in Non-Primate Mammals
In mammalian lineages outside the primate order - such as carnivores, ungulates, and cetaceans - the relationship between encephalization and sociality takes a distinct qualitative form. Rather than scaling linearly with group size, large brains in these clades are categorically associated with specific mating systems. Species characterized by monogamous, pair-bonded mating systems consistently exhibit the largest relative brain sizes within their respective families 313.
This dynamic suggests that the primary driver of encephalization across broader mammalian evolutionary history was the intense cognitive demand of selecting an optimal mate, coordinating behavior, and maintaining a durable pair-bond over a lifespan. Anthropoid primates appear to represent an evolutionary anomaly; they co-opted the neuroendocrinological pathways originally evolved to maintain monogamous pair-bonds and generalized these processes to non-reproductive group members. This generalization gave rise to the concept of platonic "friendship," facilitating the formation of the massive, cohesive groups unique to primates 313.
Marsupial brain evolution presents an entirely separate paradigm. In a comprehensive, phylogenetically imputed Bayesian model evaluating 175 marsupial species, ecological and social predictors completely failed to explain relative brain size variation. Instead, life-history traits - specifically litter size - emerged as the sole significant predictor, underscoring how reproductive parameters can severely confound broad socio-cognitive hypotheses 35.
The Ectotherm Challenge
Further challenging the universality of the social brain hypothesis is the comparative study of ectotherms, particularly teleost fishes. Endothermic mammals possess brains roughly ten times larger than ectotherms of similar body size 36. However, despite their small absolute and relative brain volumes, numerous fish species exhibit highly advanced social behaviors, individual facial recognition, and complex decision-making architectures 363738.
Intra-species comparisons of highly social fish demonstrate that populations engaging in frequent social interactions exhibit larger total brain volumes. However, this expansion is highly localized, showing a relative enlargement of the telencephalon (associated with social decision-making in fish) and a corresponding reduction in the hypothalamus 37. This mosaic adaptation demonstrates that complex social processing does not intrinsically require the massive absolute neural tissue seen in endotherms. Consequently, researchers argue that macro-evolutionary theories of brain expansion must expand to incorporate sensory-motor adaptations, which more accurately explain the massive neural tissue differences between mammals and fishes 3036.
Functional Neuroimaging of the Social Brain
The integration of functional neuroimaging (fMRI) and resting-state connectivity analyses with evolutionary anthropology has provided a precise biological locus for the social brain hypothesis. If social complexity drove primate encephalization, specific neural networks should demonstrate dedicated processing for social information, and their structural integrity should correlate with an individual's social capacity 143940.
The Default Mode Network and Theory of Mind
Neuroimaging confirms that the human brain possesses highly specialized, distributed networks attuned to navigating social dynamics. Chief among these is the Default Mode Network (DMN), a cortical system comprising the medial prefrontal cortex (mPFC), posterior cingulate cortex (PCC), precuneus, angular gyrus, and the right temporoparietal junction (rTPJ) 144142.
While the DMN exhibits high metabolic activity when the brain is at conscious rest and unengaged with external tasks, it is functionally intrinsic to self-referential thought and mentalizing. Mentalizing - synonymous with Theory of Mind (ToM) - is the cognitive capacity to infer, evaluate, and model the mental states, beliefs, and intentions of others 144143. This capacity is the operational prerequisite for empathy, tactical deception, and the maintenance of large social alliances 4144.
Structural MRI studies reveal that the social brain hypothesis explains individual differences within the human species just as effectively as it explains inter-species variations. Among modern humans, the gray matter volume in regions linked to mentalizing - such as the orbitofrontal cortex (OFC), anterior cingulate cortex (ACC), ventromedial prefrontal cortex (vmPFC), and the superior temporal sulcus (STS) - correlates significantly with an individual's self-reported social network size, as well as their proficiency in recognizing facial cues 143944.
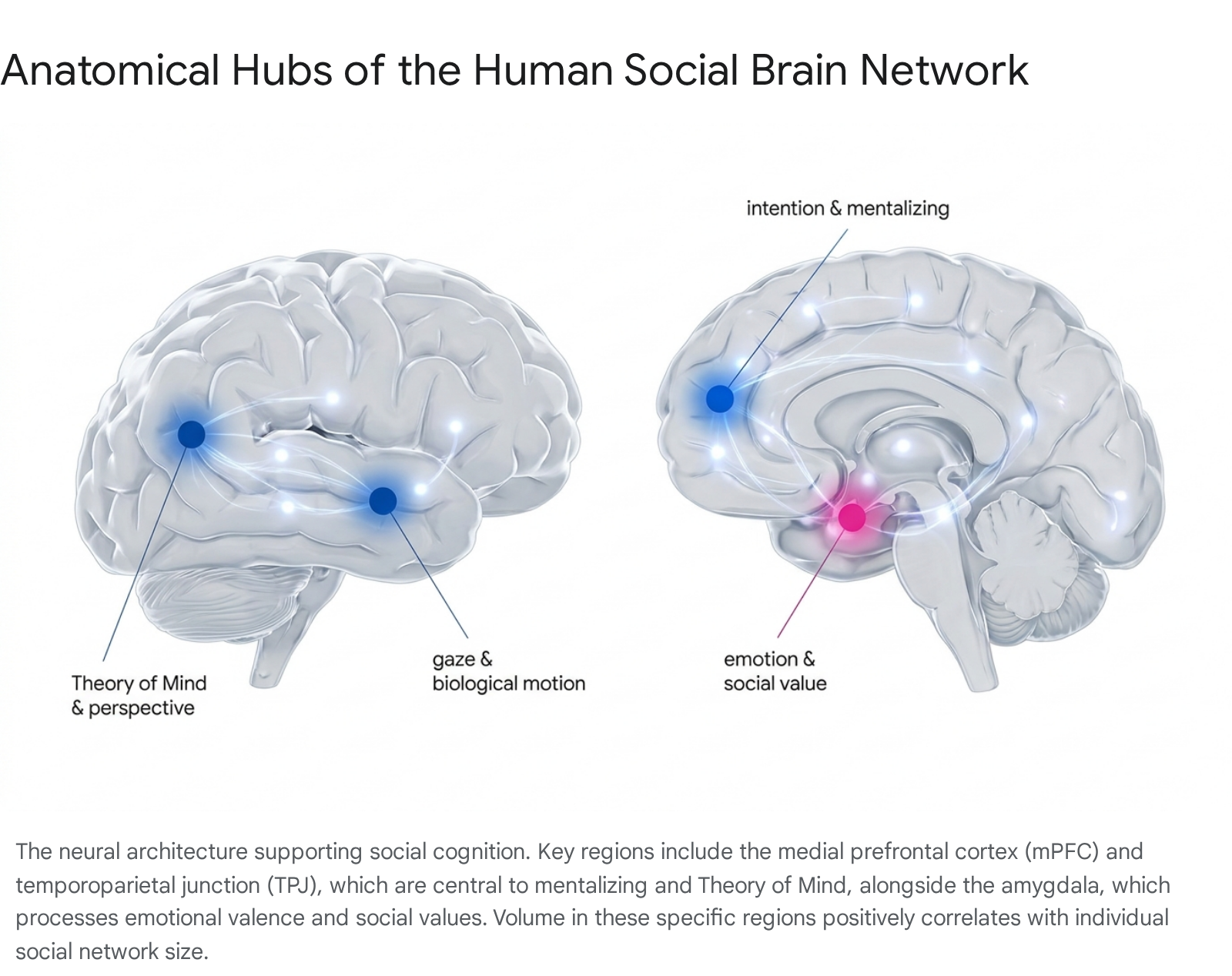
Crucially, parallel structural correlations between amygdala volume, theory of mind networks, and network size have been empirically demonstrated in macaque monkeys, establishing a direct evolutionary continuum between non-human and human primate social neurology 44.
The Amygdala as an Integrative Social Hub
The functional architecture of the social brain relies fundamentally on the amygdala, a subcortical structure located in the medial temporal lobe. The amygdala is central to interpreting emotional valence, decoding facial expressions, and regulating autonomic responses to social threats 144245. The amygdala functions as a critical integrative hub across multiple large-scale brain networks, interacting dynamically with the DMN.
Under conditions of social stress or perceived threat, intra-network communication within the DMN decreases, while inter-network connectivity surges, allowing the amygdala to orchestrate a rapid, integrated response across cognitive domains 45. The strength of resting-state functional connectivity between the amygdala and the ventromedial prefrontal cortex (vmPFC) is heavily implicated in emotional regulation and the successful maintenance of long-term social bonds 48.
Hyperscanning and Developmental Neuroplasticity
Recent advancements in dual-brain imaging, or "hyperscanning," have elucidated how the social brain develops dynamically through interaction. Hyperscanning allows for the simultaneous collection of neural activation data from multiple interacting individuals. Studies assessing infant-to-parent dyadic communication reveal that real-time social interactions - mediated by variations in speech prosody, eye gaze, and joint attention - result in the temporal alignment and synchronization of neural oscillations between the parent and child 1439. This synchronization in reward circuit areas, including the nucleus accumbens and OFC, indicates that the human brain requires reciprocal, high-fidelity social stimulation during critical developmental windows to successfully mature the cognitive hardware required for language and social bonding 1439.
Neuroplasticity within the social brain is not restricted to development. Longitudinal studies on adults ranging from 19 to 94 years old demonstrate that cognitive decline is not inevitable. Proactive engagement in social purpose and cognitive exercises measurably improves holistic brain fitness indices, establishing that the neural networks underlying social connectedness remain highly responsive and trainable across the entire human lifespan 49.
Clinical and Neurodivergent Manifestations
The social brain hypothesis provides a powerful framework for understanding neurodevelopmental dysfunctions and psychiatric conditions marked by profound social impairment. By establishing that the brain is evolutionarily optimized for social cognition, clinical deviations can be analyzed as disruptions within these specific evolved networks.
Autism Spectrum Disorder and Network Connectivity
Autism Spectrum Disorder (ASD) is historically characterized by significant differences in social communication and behavioral flexibility. Systematic reviews of task-based and resting-state fMRI studies on autistic populations consistently reveal hypo-activity and altered functional connectivity within the core nodes of the social brain 4348. Reduced functional connectivity within the DMN, particularly between the dorsomedial prefrontal cortex (dmPFC) and the posterior cingulate cortex (PCC), is tightly correlated with difficulties in conventional mentalizing and social cognition 4148.
Furthermore, atypical structural growth trajectories of the amygdala in early childhood, coupled with reduced activation in the amygdala, fusiform gyrus, and STS during the implicit processing of emotional facial expressions, highlight the deep structural roots of social processing divergence in ASD 4348. The social motivation hypothesis posits that this hypoactivation, specifically in reward circuitries like the bilateral caudate and ACC, results in autistic individuals experiencing social stimuli as less inherently rewarding than neurotypical individuals 43.
The Double Empathy Problem
While classical neuroscience framed these differences purely as deficits in a universal "theory of mind," contemporary models such as the "double empathy problem" and the "dialectical misattunement hypothesis" challenge this anthropocentric perspective. These frameworks suggest that the social friction experienced by autistic individuals arises not from an inherent inability to connect, but from a mismatch in cognitive strategies and predictive coding algorithms between neurotypical and neurodivergent brains 50. When the sensory processing and communicative styles of two individuals differ significantly, mutual misunderstanding occurs, meaning neurotypical individuals are equally deficient at "reading the minds" of autistic individuals 50.
Subclinical Anxiety and the Anterior Temporal Lobe
Dysfunctions in social processing also manifest in mood disorders. Recent neuroimaging research highlights the role of the anterior temporal lobe (ATL) in processing social hierarchies and semantic social concepts. During the interpretation of social hierarchies - such as making decisions based on the emotional facial expressions of winners or losers - the ATL activates strongly. However, in individuals with high levels of subclinical anxiety, the ATL exhibits hyper-activation, disrupting the smooth collaboration between semantic interpretation and the emotional regulation systems of the limbic network 46. This neurobiological disruption helps explain why anxiety disorders are intrinsically linked to negative affect during the evaluation of social status and peer relationships.
Furthermore, environments characterized by extreme social stigma can short-circuit the social brain entirely. When individuals view photographs of highly stigmatized out-groups, the mPFC - the hub for mentalizing - fails to activate. Instead, regions associated with disgust are engaged 52. This suggests that dehumanization is partly a cognitive lapse wherein the brain's default social processing pathways are bypassed, reinforcing the evolutionary premise that deep social tracking is resource-intensive and reserved primarily for the in-group 52.
Coevolution of Language and Social Cohesion
A final, critical pillar of the social brain hypothesis addresses the evolutionary emergence of human language. In non-human primate societies, group cohesion is maintained by physical grooming, an activity with an inflexible time constraint. As hominin group sizes expanded - whether driven by shifting ecological landscapes, predation pressures, or the compounding benefits of cultural accumulation - the necessity for a highly efficient, time-saving method of social bonding became an evolutionary imperative 5.
Vocalizations initially served as a form of "vocal grooming," allowing an individual to pacify and bond with multiple group members simultaneously across physical distances, thereby bypassing the strict one-to-one temporal constraint of manual grooming 5. Over subsequent evolutionary epochs, this vocal system expanded into complex, grammatically structured language.
The primary adaptive function of language, within this evolutionary model, was not the communication of ecological facts or hunting strategies, but the rapid, high-volume exchange of social information - gossip. Language enables individuals to learn about the behavioral characteristics, dominance alliances, and trustworthiness of group members without requiring direct, time-consuming observation of every interaction 5.
Analyses of modern human conversational dynamics confirm this hypothesis: irrespective of cultural or geographic context, approximately 60% of conversational time in unconstrained environments is devoted to discussing personal experiences, relationships, and third-party behavior 5. Consequently, the evolution of human language is inextricably linked to the neuroanatomical demands of navigating a massively expanded, highly bonded social network 1440. The primate brain is ultimately an organ forged by the pressures of community, where the continuous requirement to interpret, predict, and collaborate with peers has fundamentally shaped the architecture of human cognition.