Social Cognition in Autism Spectrum Disorder
Social cognition encompasses the complex network of psychological and neural processes required to perceive, interpret, and respond dynamically to the social signals of others. Within the context of autism spectrum disorder (ASD), social cognition has historically been framed exclusively through a paradigm of innate deficit, operating under the assumption that the autistic brain lacks the fundamental cognitive machinery necessary to execute empathy and mentalizing. However, the proliferation of advanced structural and functional neuroimaging technologies, integrated with cross-cultural developmental psychology and the emerging neurodiversity paradigm, has substantially revised this perspective. Current research literature demonstrates that social cognition in ASD is characterized by atypical neural network connectivity, intense localized perceptual autonomy, and bidirectional communication mismatches, rather than an absolute biological absence of social computational ability.
Neuroanatomical Correlates of the Social Brain
The "social brain" consists of a widely distributed, highly interacting set of cortical and subcortical regions responsible for face processing, intention attribution, and emotional regulation 12. In typical neurodevelopment, these regions coordinate to facilitate rapid, largely automatic evaluations of social environments. In autistic individuals, however, neuroimaging methodologies - ranging from functional magnetic resonance imaging (fMRI) to voxel-based morphometry (VBM) and diffusion tensor imaging (DTI) - reveal persistent differences in localized activation, cellular composition, and volumetric trajectories 134.
Regional Alterations in Cortex and Subcortex
At the localized anatomical level, variations in brain regions associated with primary social perception represent some of the most robust and replicated findings in ASD research. The amygdala, which is critical for early-stage emotion processing and the rapid evaluation of facial expressions, demonstrates a profoundly atypical developmental trajectory. Structural MRI studies indicate an initial overgrowth and volumetric enlargement of the amygdala in young autistic children, which subsequently tends to normalize, or even diminish in volume, by adolescence and adulthood 56. Functionally, individuals with ASD frequently exhibit hypoactivation of the amygdala when processing complex social judgments, such as evaluating the trustworthiness of human faces 27. Furthermore, postmortem and structural studies have observed smaller local gray matter volumes in the bilateral amygdala, left anterior insula, and medial prefrontal cortex (mPFC) in autistic cohorts relative to neurotypical controls 1.
Downstream from the amygdala, the fusiform gyrus - specifically the lateral region designated as the fusiform face area (FFA) - is integral to the perception of personal identity and facial configuration. Numerous task-based fMRI studies have reported significant hypoactivation of the FFA when autistic individuals view human faces 1238. Early cognitive interpretations posited that this hypoactivation reflected a fundamental structural impairment in social perception. However, studies utilizing non-face social attribution tasks, such as observing interactions among simple geometric shapes, have demonstrated robust FFA engagement in typical adults, challenging the view that the FFA is strictly face-exclusive 8. This suggests that abstract semantic information associated with social computation is encoded in the fusiform gyrus. Therefore, the reduced activation to human faces in ASD may reflect a divergent allocation of visual attention, or alternative developmental scaffolding, rather than an inherent inability of the tissue to process social stimuli 69.
Other critical regions implicated include the temporoparietal junction (TPJ) and the medial prefrontal cortex (mPFC). The TPJ is responsible for encoding "other-relevant" information and predicting behavior during interactions, whereas the mPFC is engaged in monitoring both self and other mental states 101112. Task-based imaging frequently reveals hypoactivation in these regions during explicit theory of mind or mentalizing paradigms 1011. Furthermore, evidence extending to the cerebellum highlights significant cellular anomalies. Postmortem brain tissue analyses demonstrate a striking 79% incidence of substantially decreased Purkinje cell size and density in the posterolateral neocerebellar and archicerebellar cortices of autistic individuals 24. Because Purkinje neurons represent the sole output mechanism of the cerebellar cortex, this anatomical disruption is hypothesized to cascade into higher-order cognitive and motor integration deficits 4.
| Brain Region | Primary Social Cognitive Function | Consistent Neuroimaging Findings in ASD |
|---|---|---|
| Amygdala | Rapid emotion processing, threat detection, and early facial evaluation | Early volumetric overgrowth shifting to normalization/reduction; hypoactivation during explicit social judgment tasks 1257. |
| Fusiform Face Area (FFA) | Perception of personal identity and facial structural configuration | Hypoactivation during face-viewing tasks; atypical neurodevelopment highlighting altered visual attention allocation 23813. |
| Temporoparietal Junction (TPJ) | Encoding "other-relevant" information and intention attribution | Hypoactivation during mentalizing tasks; reduced interhemispheric and network connectivity 101112. |
| Medial Prefrontal Cortex (mPFC) | Monitoring self and other mental states, social problem-solving | Reduced activation during self-referential processing; acts as a causal target rather than a causal source in network flow 1101114. |
| Cerebellum | Motor integration and coordination of rapid cognitive sequencing | Marked decrease in Purkinje cell density and size; atypical structural connectivity with the Default Mode Network 24. |
Structural Developmental Trajectories
Neurodevelopmental differences in ASD are not limited to discrete subcortical nuclei but extend to the global architecture of the brain. Prospective neuroimaging studies of infants at high familial risk for ASD reveal that a hyperexpansion of the cortical surface area occurs between 6 and 12 months of age, which subsequently precedes the overall brain volume overgrowth typically observed between 12 and 24 months 515. This accelerated expansion is largely confined to surface area rather than cortical thickness, and is particularly pronounced in the frontal and temporal lobes 4515.
As aging progresses, studies indicate that autistic individuals tend to exhibit thinner cortices and reduced surface area compared to their neurotypical peers 15. This trajectory - early overgrowth followed by arrested growth or premature thinning - suggests a dysregulation in the typical processes of synaptic proliferation and subsequent pruning 416. An overabundance of unpruned synapses inhibits efficient cognition while simultaneously demanding increased cerebral blood flow. Consequently, neuroimaging of high-functioning autistic youths reveals ongoing hyper-perfusion and increased oxygen metabolism in frontal brain areas, indicating a delayed or altered maturation of the socio-emotional circuitry 16.
Functional Connectivity and Network Architecture
While localized structural differences establish the neurobiological substrate, social cognition requires the rapid, dynamic integration of distributed neural networks. Another perspective conceptualizes ASD primarily as a disorder of neural information processing and connectivity.
The Default Mode Network
The Default Mode Network (DMN) is heavily implicated in internal mentation, self-referential thought, autobiographical memory, and mentalizing. Its core nodes consist of the posterior cingulate cortex (PCC), the mPFC, and the TPJ 1011. In typical development, the DMN forms an internally coherent system that deactivates during externally directed tasks.
Resting-state and task-based functional connectivity studies consistently highlight atypical intra- and inter-network communication involving the DMN in autism 1718. A dominant developmental theory suggests an overall shift from neural hyperconnectivity in autistic childhood to pervasive hypoconnectivity in adolescence and adulthood, particularly regarding long-range pathways 1116.
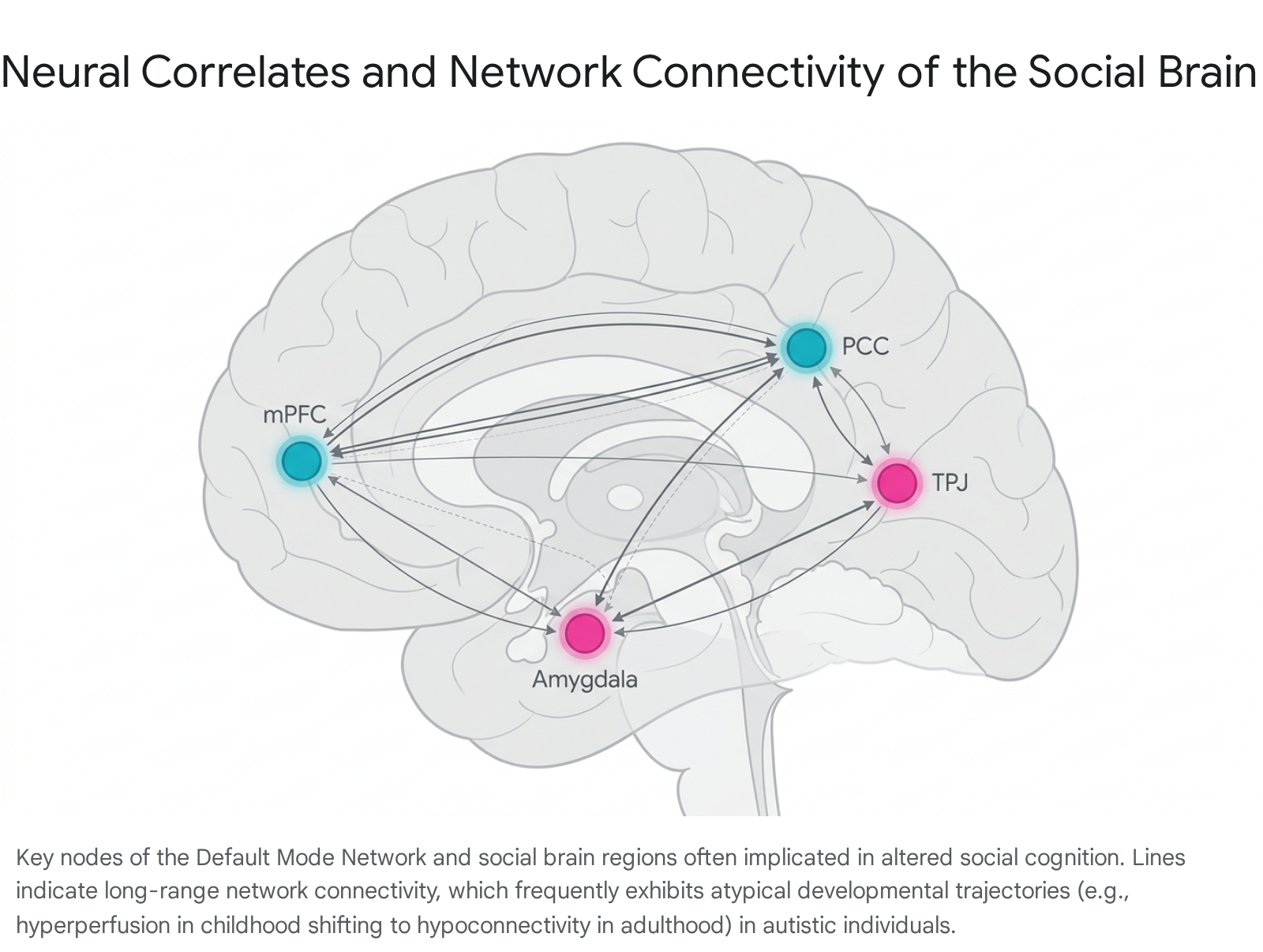
When evaluating intra-DMN function in adolescents and adults with ASD, researchers predominantly identify underconnectivity, such as reduced signaling between the PCC and mPFC 1117.
Recent methodologies applying Granger causality and Liang information flow algorithms have allowed researchers to map directional causal connectivity rather than mere correlation. These analyses demonstrate that in neurotypical populations, the dorsal mPFC acts as a primary causal source of information flow within the DMN. In contrast, in the autistic brain, the mPFC operates predominantly as a causal target 14. This reversal of information hierarchy indicates a fundamental perturbation in how the brain sequences and distributes social computations, leading to cascading delays in subsequent cognitive operations 1419.
Salience and Executive Control Networks
In typical individuals, task-positive networks such as the Executive Control Network (ECN) and the Salience Network (SN) operate with strict functional segregation and are highly anti-correlated with the DMN 18. The Salience Network, anchored in the anterior insula and dorsal anterior cingulate cortex, acts as a switchboard to dynamically engage or disengage the DMN and ECN based on environmental demands 1418.
In ASD, the typical anti-correlation between the task-positive ECN and the task-negative DMN is significantly reduced. This results in broad network "cross-talk," with studies showing extensive overconnectivity between the DMN and the right ECN 18. Concurrently, the SN exhibits widespread intra-network underconnectivity, which correlates heavily with the severity of socio-communicative and sensory symptoms 18. The failure of the anterior insula to properly modulate and segregate these networks may leave the autistic brain suspended in a state of overlapping internal mentation and external processing, rendering rapid social exchanges cognitively exhaustive 131418.
Cognitive Frameworks for Social Processing
The interpretation of raw neuroimaging data heavily relies on cognitive frameworks that conceptualize how autistic individuals process their physical and social environments. Moving away from monolithic deficit models, contemporary cognitive science recognizes multiple overlapping dimensions of variance.
Theory of Mind and Mentalizing
Theory of Mind (ToM) represents the cognitive capacity to attribute mental states, beliefs, and desires to oneself and others. Historically, it has been a focal point of autism research 202122. Early models, tested via paradigms like the Sally-Anne false-belief task, proposed a unilateral "mind-blindness" in autism. Neurotypical children reliably pass standard false-belief tests around age four, demonstrating an intuitive grasp that others can hold beliefs differing from reality; however, autistic children frequently exhibit significant delays, sometimes failing to pass until reaching a verbal mental age of 11 202324.
However, modern research indicates that ToM abilities in ASD are highly nuanced rather than entirely absent. Functional neuroimaging demonstrates that while autistic individuals exhibit reduced activation in traditional mentalizing hubs (such as the mPFC and TPJ) during spontaneous social cognition, high-functioning autistic adults can perform adequately on explicit ToM tasks 2022. To achieve this, they often activate alternative, domain-general brain regions associated with explicit logical reasoning and problem-solving 2024. This suggests that what is an automatic, intuitive social perception in typical development becomes an active, computationally demanding executive task for the autistic individual.
Empathizing-Systemizing Theory
Expanding upon ToM, the Empathizing-Systemizing (E-S) Theory, advanced by Simon Baron-Cohen, organizes human cognitive styles along two independent, orthogonal dimensions 252627. Empathizing is defined as the drive to identify another person's mental state and respond with an appropriate emotion, encompassing both cognitive and affective components 2628. Systemizing is defined as the drive to analyze, deconstruct, or construct physical, mechanical, or abstract rule-based systems 2628.
The E-S framework classifies individuals into distinct "brain types" based on the mathematical discrepancy between their empathizing and systemizing quotients 2527. The Extreme Male Brain (EMB) theory, an extension of the E-S model, proposes that typical females generally score higher on empathizing (Type E), typical males score higher on systemizing (Type S), and autistic individuals display a hyper-masculinized profile. The autistic cognitive profile predominantly features average or elevated systemizing coupled with below-average empathizing (Extreme Type S) 252728.
| Cognitive Profile (E-S Theory) | Defining Characteristic | Distribution Patterns in Research |
|---|---|---|
| Type E (Empathizing) | Empathizing capability dominates Systemizing (E > S) | More frequently observed in neurotypical females 2528. |
| Type B (Balanced) | Empathizing and Systemizing drives are relatively equal (E = S) | Distributed evenly across the general population 2527. |
| Type S (Systemizing) | Systemizing capability dominates Empathizing (S > E) | More frequently observed in neurotypical males 2528. |
| Extreme Type S | Systemizing is profoundly elevated; Empathizing is diminished (S >> E) | Disproportionately characterizes the cognitive phenotype associated with ASD 2728. |
A massive validation study analyzing over 600,000 individuals confirmed these population-level trends, demonstrating that D-scores (the mathematical difference between E and S) account for 19 times the variance in autistic traits compared to other demographic variables like biological sex 28. However, critics note that performance on these measures relies heavily on self-perception and self-reporting tools, which may not accurately capture the lived experience of individuals with intellectual disabilities or atypical language development 26.
The Enhanced Perceptual Functioning Model
An alternative and highly influential approach to cognitive variance in autism looks past high-level social reasoning to foundational sensory perception. The Enhanced Perceptual Functioning (EPF) model, proposed by Laurent Mottron, suggests that autistic perception is locally oriented and maintains high autonomy from top-down conceptual processing 293031.
The EPF model outlines several core principles of autistic perception, noting that the autistic brain allocates a disproportionate share of neural resources to low-level primary perceptual cortices 2930. This allocation manifests in superior low-level discrimination, heightened pitch processing, enhanced perception of static stimuli, and superiority in visual search tasks 293334. Rather than indicating a deficit in global processing - as suggested by earlier "weak central coherence" theories - EPF posits that localized processing is simply robust, mandatory, and highly autonomous in autism 2931.
Because high-level fluid social processing (such as tracking the holistic, complex movement of facial muscles during a conversation) relies on seamless, top-down neural integration, the autistic brain's intense autonomous processing of raw, micro-level sensory details may inadvertently overwhelm or bypass conventional social computational pathways 293134. For clinicians and educators, the EPF model highlights the necessity of reducing downstream task loads; autistic perception is highly efficient at the raw-signal stage, meaning sensory sensitivities and localized details must be accommodated before demanding complex social integration 34.
Confounding Variables in Social and Emotional Processing
The assertion that autism is fundamentally characterized by an empathy deficit has faced intense scrutiny as methodological precision has improved. Current neuroscience indicates that supposed social deficits in autism are frequently conflated with co-occurring psychological conditions, demanding a higher degree of granularity.
Alexithymia and Interoception
A critical distinction in behavioral science separates cognitive empathy (the ability to accurately identify and track another's emotional state) from affective empathy (the instinctual emotional resonance or response to that state). A comprehensive 2024 systematic meta-analysis authored by Mottron, Cusson, and colleagues revealed a large effect size (g = -0.85) indicating differences in cognitive empathy among autistic populations. However, differences regarding affective empathy were remarkably small (g = -0.17) and became non-significant when restricted solely to high-quality, ecologically valid studies 32.
The primary source of inconsistency in historical empathy research is the confounding presence of alexithymia. Alexithymia is defined as a subclinical difficulty in identifying and describing one's own emotional states, and it co-occurs in approximately 50% of the autistic population 33. The "alexithymia hypothesis" proposes that the empathy and emotion-recognition deficits traditionally attributed to ASD are actually driven by this co-occurring condition. Hierarchical regression models demonstrate that once alexithymia severity is controlled for, autism diagnostic group status ceases to significantly contribute to variance in personal distress or emotional empathic responsiveness 33.
Similarly, research into interoception - the neural perception of internal bodily and physiological states - is heavily confounded. While early theories suggested interoceptive deficits were core to autism, studies matching autistic individuals and typically developing youth find no significant differences in interoceptive sensibility once alexithymia is accounted for 33. Consequently, blending measures of "empathic concern" and "personal distress" into a single monolithic score of "emotional empathy" has historically distorted the literature, perpetuating a false narrative that affective empathy is unilaterally absent in ASD 3233.
The Relational Turn and Double Empathy
For decades, the standard assumption across clinical science was that communication breakdowns between autistic and non-autistic individuals were entirely the result of the autistic person's innate social impairment. The "Double Empathy Problem," formulated by autistic sociologist Damian Milton in 2012, challenges this unilateral premise, reframing the communication barrier as a bidirectional relational mismatch 3435.
Bidirectional Communication Mismatch
The Double Empathy theory posits that empathy and mutual understanding require shared dispositions, communication styles, and social-cognitive experiences. Because autistic and neurotypical individuals possess vastly different neurological and developmental landscapes, they will naturally struggle to accurately mentalize each other 343637. Therefore, the theory predicts that neurotypical individuals should exhibit reciprocal, corresponding difficulties in reading autistic mental states 3435.
Recent behavioral experiments provide robust empirical support for this hypothesis. In studies utilizing diffusion chains (passing a narrative story from person to person), researchers found that information degraded rapidly in mixed-neurotype chains containing both autistic and allistic (non-autistic) participants. Conversely, information transfer remained highly accurate, and interpersonal rapport was rated significantly higher, within homogenous chains composed solely of autistic individuals or solely of non-autistic individuals 3538. Furthermore, empathic accuracy tasks reveal that neurotypical observers have significantly lower accuracy scores when attempting to read the emotional states of autistic narrators recounting autobiographical events, particularly regarding expressions of happiness and sadness 38. Observer studies also note that neurotypical evaluators frequently misinterpret autistic directness as negative affect, demonstrating that communication mismatch alone is sufficient to generate negative social perception 3537.
Cultural Context and Social Performance
Further supporting the relational nature of social cognition, cross-cultural studies indicate that autistic social performance is not biologically fixed, but is highly sensitive to cultural and contextual expectations. A recent comparative cross-cultural study of mentalizing tasks involving moving geometric shapes investigated socio-cognitive abilities across UK and Japanese cohorts 39.
The findings revealed a stark cultural divide. British neurotypical adults struggled significantly to interpret animations created by their autistic peers, demonstrating the classic double empathy gap. However, in the Japanese cohort, autistic and non-autistic adults interpreted each other's animations with comparable, high accuracy. Furthermore, Japanese autistic participants outperformed both British groups 39. This implies that Western diagnostic criteria and social assessments may capture specific cultural manifestations of communication mismatch rather than universal biological deficits. The Double Empathy paradigm, therefore, recontextualizes autistic social cognition as a different, not inherently deficient, mode of processing that resembles cross-cultural communication challenges 3639.
Methodological Constraints in Neuroscience Evidence
The rapid evolution of autism research is closely tied to the recognition of severe structural limitations in how neurobiological and behavioral data are collected, processed, and generalized.
The Ecological Validity of Neuroimaging
While functional MRI has successfully mapped the topography of the social brain, standard neuroimaging paradigms suffer from chronically low ecological validity 4041. The MRI scanner environment requires participants to remain entirely static, isolated in a confined tube, while viewing highly controlled, two-dimensional, simplified stimuli. Because real-world human social cognition is inherently dynamic, bidirectional, multimodal, and situation-dependent, static fMRI assessments inherently strip away the naturalistic variables that define genuine social exchange 404142.
Consequently, there is a risk that neuroimaging captures isolated cognitive mechanics rather than true social application. Furthermore, preclinical neuroscience highlights the "shiftability paradigm" - the principle that the only way to establish how neurotransmitters modulate information processing is to physically change them and observe the shift. Human neuroimaging relies heavily on correlational association rather than direct experimental neurochemical manipulation, leaving a substantial translational gap in understanding the precise neurobiology of ASD 43.
To bridge the ecological gap, modern study designs are increasingly supplementing static neuroimaging with Ecological Momentary Assessment (EMA). By prompting participants to record their feelings, physiological states, and social interactions via mobile technology in their naturalistic, daily environments, researchers capture the nuanced, shifting reality of lived autistic experiences 4044. Systematic reviews indicate that EMA is highly feasible and acceptable for autistic adults, providing a necessary counterweight to sterile laboratory findings 44. Furthermore, advancements in computational modeling, such as Activity Flow Mapping (AFM), are beginning to predict task-evoked social brain activity directly from non-invasive resting-state functional connectivity, offering more flexible data collection paradigms 45.
Demographic Bias and Sampling Representation
Compounding the limitations of ecological validity is a persistent, systemic bias in participant sampling. A disproportionate volume of cognitive neuroscience research historically relies on WEIRD (White, Educated, Industrialized, Rich, Democratic) populations 46. Weighting pediatric MRI datasets to match the actual demographics of the general U.S. population dramatically shifts the established timelines of normative brain development - moving the peak of brain surface area maturation forward by over two years. This underscores the reality that our foundational maps of brain development are severely skewed by non-representative data 46.
In autism specifically, research samples are overwhelmingly white, male, and drawn from middle-to-high socioeconomic strata 5051524748. While CDC surveillance estimates indicate an overall autism prevalence of 1 in 36 (or 2.8%) among eight-year-olds in the U.S., significant demographic disparities remain hidden beneath the average 495051. Prevalence estimates for profound autism, intellectual disability co-occurrence, and age of diagnosis vary radically by race. For example, 50.8% of Black autistic children are classified with intellectual disability compared to 31.8% of White children, and Hispanic and Black children are vastly overrepresented in "suspected" rather than "confirmed" diagnostic categories due to a lack of complete healthcare records 495052.
Furthermore, standard clinical assessment tools act as an artificial bottleneck for research inclusion, systematically excluding female participants. The Autism Diagnostic Observation Schedule (ADOS), considered the gold standard for clinical classification, was originally developed and calibrated using predominantly male samples 53.
Analyses of large public datasets reveal that studies requiring an ADOS screening for participation feature a severe male-to-female ratio of roughly 8:1. In contrast, studies accepting community-level clinical diagnoses report ratios much closer to 2:1 or 1:1 53.
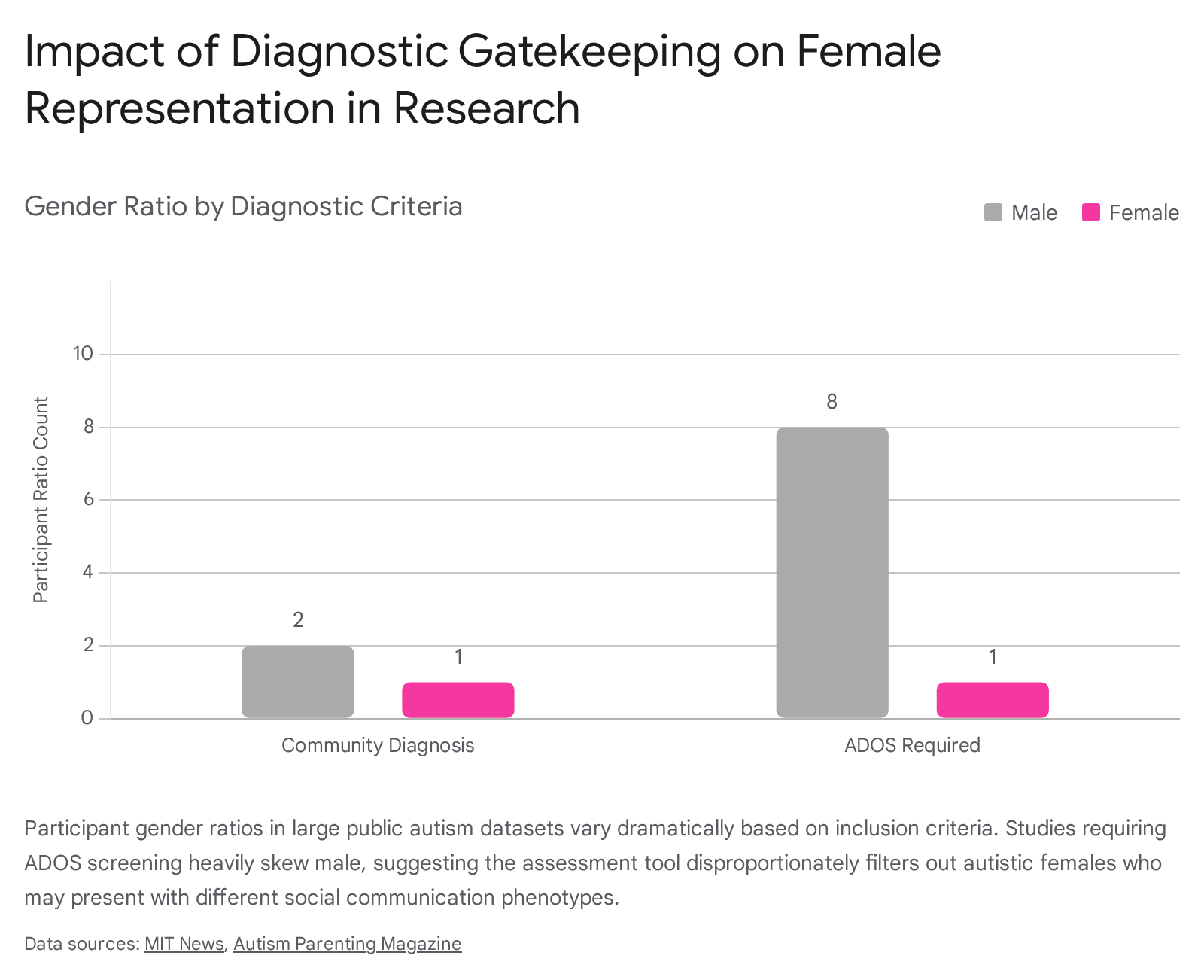
This diagnostic gatekeeping results in profound epistemic injustice. The female presentation of autism - which frequently features high levels of "camouflaging" or "masking," where social difficulties are suppressed to blend into neurotypical environments - is routinely excluded from the data pools used to map the neurobiology of the condition 51535455. Consequently, existing literature risks defining a distinctly white, male expression of autism as the universal neurological standard 515255.
The Neurodiversity Paradigm and Systems Theory
In direct response to the limitations of medicalized, deficit-based models and the biases inherent in historical sampling, the neurodiversity paradigm has gained substantial theoretical and practical traction in cognitive science and public policy.
Reconceptualizing Deficit and Difference
Championed by sociologists, philosophers, and autistic scholars such as Judy Singer, Nick Walker, and Robert Chapman, the paradigm frames autism, ADHD, dyslexia, and other forms of neurocognitive variation not as pathological dysfunctions, but as natural, evolutionarily valid expressions of human biodiversity 54566357. The neurodiversity paradigm heavily aligns with the social and relational models of disability, as well as ecological systems theory 5765. From this perspective, the social and cognitive disability experienced by autistic individuals is predominantly generated by systemic marginalization and an environmental mismatch with neuro-normative expectations, rather than a broken biological mechanism 19565758.
This paradigm entails a rejection of species-standard normative framing. Instead of defining the neurotypical brain as the "correct" baseline against which the autistic brain is measured as a "deficit," it views neurotypes much like variations in gender or cultural background 545663. This theoretical shift is accompanied by a deliberate linguistic move toward "identity-first" language (e.g., "autistic person" rather than "person with autism"), validating the neurotype as an intrinsic, inseparable component of selfhood and identity rather than an external disease 575859.
Clinical and Educational Implications
Modern neurodiversity-affirming practices do not reject the reality of autistic struggles, nor do they dismiss the value of medical and psychological support 566061. Instead, they reorient the ultimate goal of intervention. Traditional clinical models, heavily influenced by the deficit perspective, often aimed for "normalization" - training autistic individuals to mask their traits, suppress repetitive behaviors (stimming), and simulate neurotypical social cognition 19365961. Such masking is now closely associated with severe longitudinal mental health consequences, including high rates of burnout, anxiety, and depression 3637.
Conversely, neurodiversity-affirming interventions focus on modifying the sensory environment, building autonomous self-advocacy, and establishing cross-neurotype communication bridges. Clinical supports are viewed as essential mechanisms for participation and skill development (such as teaching adaptive coping skills) rather than threats to autistic identity 365960. By moving beyond the monolithic narrative of social deficit, the integration of cutting-edge neuroimaging, nuanced behavioral science, and the neurodiversity paradigm provides a holistic, rigorous, and profoundly more accurate understanding of the autistic mind.