Sensory integration and the binding problem in neuroscience
Introduction to the Functional and Phenomenal Binding Problem
The central nervous system continuously processes a relentless, massively parallel influx of sensory information originating from complex environmental and somatic sources. These signals are transduced by specialized sensory epithelia and subsequently routed through anatomically and functionally divergent neural pathways. In the primate visual system, processing strictly bifurcates beyond the primary visual cortex into a dorsal stream - projecting to the posterior parietal cortex to compute spatial navigation and action execution coordinates - and a ventral stream - projecting to the inferior temporal cortex for granular object identification and form representation 1234. Auditory, somatosensory, and proprioceptive modalities undergo similarly distributed, parallel processing across distinct cortical maps 356. Despite this extreme anatomical segregation, humans do not perceive a fragmented, disjointed world of decoupled shapes, colors, pitches, and tactile pressures. Instead, conscious perception manifests as a cohesive, seamlessly unified gestalt. The question of how the brain rapidly and accurately recombines these distributed neural representations into single objects or events without catastrophic interference is known as the binding problem 78.
In cognitive science and theoretical neuroscience, the binding problem is rigorously divided into two interrelated domains: functional binding and phenomenal binding 91011. Functional (or computational) binding refers to the specific biophysical, network, and algorithmic mechanisms by which local neural circuits tag, route, and correlate disparate streams of data so that downstream motor effectors and cognitive evaluations treat them as a single entity 1011. Phenomenal binding refers to the subjective, first-person integration of these features - the metaphysical and neurobiological mechanism by which the brain generates a unified macro-scale conscious experience from billions of localized micro-units of information 91011. Reconciling these two domains requires mapping micro-level synaptic and dendritic computations to macro-level oscillatory networks and global cognitive architectures.
Principles of Multisensory Integration
The foundation of resolving the functional binding problem lies in the brain's capacity for multisensory integration, a process that dramatically enhances detection accuracy, discrimination precision, and processing speed by fusing noisy sensory signals that emanate from a common causal source 1112. The nervous system does not arbitrarily bind stimuli; rather, it relies on strict statistical probabilities and physical parameters to determine whether cross-modal cues share a single origin or constitute independent environmental events.
Spatial and Temporal Contiguity
The probability that the nervous system will bind two or more sensory cues into a single perceptual object depends heavily on spatial and temporal rules. Information arising from the same approximate spatial coordinates and occurring within a specific temporal proximity is inherently more likely to be integrated 141314. This temporal proximity is operationally defined as the "temporal binding window" - a calibrated interval within which physically asynchronous cross-modal stimuli are subjectively perceived by the organism as simultaneous 111314.
The boundaries of the temporal binding window are neither rigidly fixed nor innate; they exhibit high plasticity and undergo a surprisingly protracted developmental trajectory extending well into late adolescence 111315. In neonates, the ability to synthesize paired multisensory cues does not occur rapidly but develops gradually alongside sensory experience, requiring extensive exposure to normal visual scenes and auditory inputs to configure the underlying neural circuits 15. Psychophysical research analyzing audiovisual simultaneity judgment tasks demonstrates that children and young adolescents exhibit significantly wider temporal binding windows compared to adults 1314. For example, children and adolescents are highly likely to bind asynchronous stimuli into a single percept even when auditory cues precede visual cues by extensive stimulus onset asynchronies of 150 to 350 milliseconds - intervals that adults easily segregate into two distinct events 1314. Through continuous interaction with the environment, the brain calibrates the disparate speeds of physical signal propagation (e.g., light traveling faster than sound) against the speeds of internal neural processing (e.g., auditory transduction occurring faster than visual phototransduction), progressively narrowing the temporal binding window to optimize perceptual accuracy 13.
Perceptual learning paradigms demonstrate that even in adulthood, the temporal binding window remains malleable. Simultaneity judgment training paired with trial-by-trial feedback induces a marked, stable narrowing of the temporal binding window, indicating that top-down cognitive knowledge and active error correction can recalibrate low-level sensory binding parameters 11. The baseline prior expectation that two stimuli share a common source is referred to as the "binding tendency," which exhibits significant inter-individual variability across spatial localization, size-weight perception, and speech perception tasks 11.
The Principle of Inverse Effectiveness and Superadditivity
Beyond contiguity, the magnitude of integration is governed by the principle of inverse effectiveness. This principle dictates that the degree of multisensory enhancement is inversely proportional to the salience or effectiveness of the individual unisensory stimuli 141819. When individual sensory cues are weak, degraded, or environmentally ambiguous (e.g., visual cues obscured by fog or auditory cues masked by background noise), their convergence results in a "superadditive" neural response 19161718. In this state, the combined multisensory activation significantly exceeds the linear algebraic sum of the isolated unisensory responses 19161718.
Optimal multisensory integration requires overlapping neural activity patterns rather than mere simultaneous stimulus onset. When evaluated against rigorous benchmark criteria rooted in signal detection theory, multisensory behavioral performance consistently outpaces the most stringent sum of unisensory performance levels, exhibiting an approximate 50% proportional enhancement in accuracy 1416. This superadditive effect shows minimal variance across diverse testing sessions, animal sex, spatial configurations, or trial histories, establishing multisensory binding as a highly reliable neural mechanism essential for survival 16.
Neuroanatomical Hubs of Sensory Convergence
The physical binding of distributed features requires specific anatomical substrates capable of receiving, collating, and modulating multiple unimodal streams. Historically, sensory processing was viewed as strictly modular up to the highest levels of the association cortex. However, contemporary connectomics and electrophysiological recordings reveal a sophisticated network of subcortical and cortical hubs that facilitate both early and late-stage multisensory convergence 19202122.
Single-Cell Integration in the Mauthner Network
While cortical networks illustrate macroscopic integration, definitive proof of functional binding at the extreme micro-scale is established via the Mauthner cell system in teleost fish, such as goldfish. Mauthner cells are a pair of giant reticulospinal command neurons responsible for initiating an explosive startle escape behavior known as the C-start 191723. A single action potential in one Mauthner cell activates contralateral spinal motor circuits, providing a rare, direct 1:1 link between single-neuron computation and behavioral execution 1823.
The Mauthner cell receives visual inputs (e.g., looming stimuli mimicking predators) and auditory inputs (e.g., sound pips mimicking water displacement) through anatomically segregated dendritic trees 1918. In vivo intracellular recordings demonstrate that the Mauthner cell acts as an independent multisensory integrator 1723. The convergence of subthreshold visual and auditory postsynaptic potentials on the Mauthner cell dendrites evokes a supralinear response that directly increases the probability and drastically reduces the latency of the escape behavior 191823. Adding a weak, low-intensity auditory stimulus early in a visual loom sequence yields an enhanced integration magnitude that strictly obeys the inverse effectiveness principle 171823. Mechanistically, this binding relies on the distinct decay dynamics of feed-forward inhibition triggered by auditory and visual stimuli, as well as highly nonlinear dendritic membrane properties, proving that the earliest stages of perceptual binding and behavioral decision-making can occur within the biophysics of a single neuron 171823.
Subcortical Integration in the Superior Colliculus
In mammals, the superior colliculus in the midbrain serves as a primary model for subcortical multisensory integration. It receives direct afferent inputs from the retina, spinal cord, inferior colliculus, and widespread cortical regions, sending efferents to motor centers and the thalamus 1215. Within its intermediate and deep layers, receptive fields from visual, auditory, and somatosensory modalities spatially converge, forming a topographically aligned, two-dimensional multisensory map of external space 1222.
Recordings from over 5,000 neurons across the anatomical axes of the superior colliculus in awake mice demonstrate that multisensory neurons consistently encode temporal delays through the nonlinear summation of inputs. This nonlinearity is particularly pronounced when visual stimuli naturally precede auditory stimuli, actively mirroring the statistical realities of light and sound propagation in the physical environment 28. The superior colliculus exhibits distinct regional functional specializations; cross-correlation analyses indicate high recurrent connectivity in the medial zone, where multisensory neurons preferentially wire to other multisensory neurons, accounting for approximately 50% of their local input 28. This dense recurrent architecture optimizes the population-level decoding of temporal features and facilitates extreme spatial discriminability in the peripheral visual field 28.
Thalamic Synchronization via the Pulvinar Nucleus
The pulvinar is the largest nucleus in the primate thalamus. Scaling in evolutionary expansion with the neocortex, it serves as a higher-order, extrageniculate relay that maintains widespread, reciprocal cortico-thalamo-cortical connections with the occipital, parietal, temporal, and frontal lobes 29242526. Unlike first-order thalamic relays that simply pass raw peripheral data to the primary sensory cortex, the pulvinar occupies a strategic hub position to modulate, synchronize, and bind communication between discrete cortical zones during selective attention tasks 29242526.
The pulvinar organizes integration through a distinct topographical connectivity gradient. The anterior pulvinar communicates predominantly with highly spatiotopic regions like the early visual and parietal cortices, while the posterior pulvinar connects to less spatially organized regions like the inferior temporal cortex 24. Lesion studies in humans reveal that damage to the anterior pulvinar selectively disrupts the ability to bind features to a specific location in space, whereas posterior pulvinar damage produces deficits in binding features across time 24.
Crucially, the pulvinar actively controls cortical binding through rapid state-switching between tonic and burst firing modes 25. During selective, covert attention tasks, specific electrical microstimulation of the pulvinar triggers high-frequency bursting that rapidly synchronizes cortical spiking across distributed networks, biasing the cortical representation toward an integrated, attended target 25. Thalamic bursting functions as a dynamic routing mechanism, allowing the brain to transiently bind disparate neural ensembles to meet moment-to-moment cognitive demands, effectively coordinating multi-scale incremental feature binding 2533.
Cortical Convergence in the Parietal and Frontal Lobes
At the highest levels of cortical processing, the posterior parietal cortex operates as a central hub for integrating extrapersonal spatial arrays and sensory associations. The posterior parietal cortex, specifically the inferior parietal lobule and the intraparietal sulcus, acts as the terminus for the dorsal visual stream while maintaining dense connections to the ventral stream, the somatosensory cortex, and the prefrontal cortex 124619.
The posterior parietal cortex largely resolves the functional binding problem by anchoring decoupled object features (e.g., color, shape, motion) to specific, ego-centric spatial coordinates 720. Functional magnetic resonance imaging studies demonstrate that the spatial attention networks of the parietal lobe are preferentially activated during feature conjunction tasks - specifically when multiple objects are presented simultaneously at different locations, requiring active spatial mapping to avoid conjunction errors 7. Furthermore, memory load studies demonstrate that neural activity in the intraparietal sulcus scales directly with the number of bound features required for nonspatial working memory, with load sensitivity strengthening along a caudal-to-rostral gradient from IPS0 to IPS5 34.
The integration architecture extends into the frontal lobe via long-range anatomical association tracts 120. The right ventral inferior frontal gyrus receives convergent projections from both the dorsal stream (via the intraparietal sulcus) and the ventral stream (via the fusiform gyrus), allowing it to incorporate localized spatial representations with object identification matrices to maintain short-term feature binding 1. Additionally, anatomical tract-tracing in macaques and resting-state functional connectivity MRI in humans identify a highly centralized connectional hub located in the medial rostral dorsal caudate. This striatal hub receives dense, convergent inputs from the caudal inferior parietal lobule and multiple prefrontal networks, mediating the delicate balance between visual attentional bias, reward association, and cognitive control over bound stimuli 19.
| Anatomical Structure | Scale of Operation | Primary Binding Function | Key Mechanism of Action |
|---|---|---|---|
| Mauthner Cell | Single Neuron / Micro-circuit | Cross-modal threat detection and motor-escape initiation. | Dendritic summation of segregated visual and auditory inputs; inverse effectiveness via feed-forward inhibition decay 171823. |
| Superior Colliculus | Midbrain Nucleus | Spatial orientation and early audiovisual temporal alignment. | Topographic multisensory maps; non-linear summation of temporally delayed inputs matching physical propagation statistics 1228. |
| Thalamic Pulvinar | Diencephalic Relay | Cortical network synchronization; spatial and temporal feature coupling. | Cortico-thalamic burst firing; anterior-to-posterior connectivity gradients for spatiotopic vs temporal processing 242526. |
| Posterior Parietal Cortex | Cortical Association Area | Spatial feature binding; integration of dorsal and ventral processing streams. | Ego-centric coordinate anchoring; maintenance of feature conjunctions in visual and working memory 1272034. |
Temporal Dynamics and Neural Oscillations
While anatomical convergence pathways explain where sensory signals physically meet, they do not fully resolve how millions of distributed neurons represent a single object simultaneously without their signals catastrophically interfering. This interference, often referred to in computational literature as the superposition catastrophe, occurs when the disentangled representations of independent generative factors bleed into one another during parallel processing, leading to perceptual ambiguity 827.
A leading biological resolution to this issue relies on temporal coordination, formalized as the temporal binding theory or the correlation hypothesis 8. This framework posits that neurons encoding different features of the same object fire in precise temporal synchrony, phase-locking their oscillatory cycles. By coupling their firing rates and phase angles - particularly in the beta (13 - 30 Hz) and gamma (30 - 100 Hz) frequency bands - disparate neural ensembles signal to downstream readers that their respective features belong to a unified object 8142829.
Recent theoretical developments highlight the critical role of cortical traveling waves in managing this phase-locking 29. Slower traveling waves propagate physically across the surface of the cortex, dynamically phase-locking separate neuronal populations across disparate hierarchical levels 29. If shape is processed in the temporal lobe and color in the occipital lobe, the synchronous alignment of their oscillatory peaks essentially opens a shared temporal integration window, solving the spatial distance problem through transient temporal coherence 1429.
High-Dimensional Computing and Dendritic Architectures
The biological binding problem has direct parallels in artificial intelligence, where artificial neural networks suffer from the "Reversal Curse" - an inability to properly disentangle, bind, and generalize conceptual logic in reversible factual associations 27. When multiple generative factors interfere, conventional deep learning architectures fail to achieve the compositional understanding typical of human cognition 827.
To replicate the brain's success in conceptual and perceptual binding, computational neuroscientists have begun modeling specific biological algorithms utilizing High-Dimensional Computing and Vector Symbolic Architectures. In these models, assemblies of concept cells are represented by highly sparse, high-dimensional binary vectors 38. Information retrieval and feature binding are achieved through Behavioral Time Scale Plasticity (BTSP), an asymmetric synaptic learning rule natively observed in the CA1 region of the hippocampus 3830. Unlike classical Hebbian plasticity or Spike-Timing-Dependent Plasticity, which require repeated, highly correlated input pairings to alter synaptic weights, BTSP allows networks to form complex, conjunctive representations of diverse content in a single shot 3830.
In silico modeling demonstrates that individual pyramidal neurons equipped with active, highly nonlinear dendritic compartments can independently solve the nonlinear feature binding problem - a computational task traditionally assumed to require massive, multi-layered neuronal networks 30. This confirms that the central nervous system's resolution to the binding problem operates across multiple structural scales simultaneously, from the supralinear voltage dynamics of a single dendritic spine to the global phase-locking of the entire cerebral cortex 30.
Predictive Processing and Bayesian Causal Inference
The modern shift in understanding the binding problem moves away from purely bottom-up feature synthesis toward top-down, hierarchical inference. The Predictive Processing (or Active Inference) framework postulates that the brain functions as a continuous, proactive prediction machine. Rather than passively waiting to assemble sensory fragments arriving from peripheral nerves, the central nervous system maintains an internal, hierarchical generative model of the world and generates continuous top-down predictions regarding likely incoming sensory data 403132.
Within this framework, sensory inputs serve primarily as error signals. When bottom-up sensory data mismatches the top-down internal prediction, it generates a "prediction error," which travels up the neural hierarchy to update the generative model 40313233. The brain must determine which errors represent reliable environmental changes and which are merely statistical noise. It achieves this via precision weighting - an attentional mechanism that assigns Bayesian confidence scores to incoming signals based on contextual reliability 31333435.
The Mechanism of Bayesian Binding
Recent extensions of the Active Inference model propose a formal mechanism for resolving the binding problem termed "Bayesian Binding" 35363738. According to this theory, the generation of a unified conscious experience requires nested levels of binding. At each hierarchical stage, the system attempts to synthesize prior expectations with sensory evidence to form an approximate posterior belief 38.
Because the brain processes a chaotic, massively parallel stream of sensory data, there is an intense "inferential competition" among possible explanations for what is occurring in the external environment 36373940. Bayesian Binding dictates that the inferences that win this precision-weighted competition are those that most effectively reduce long-term uncertainty and logically cohere with the overarching global reality model 35363738.
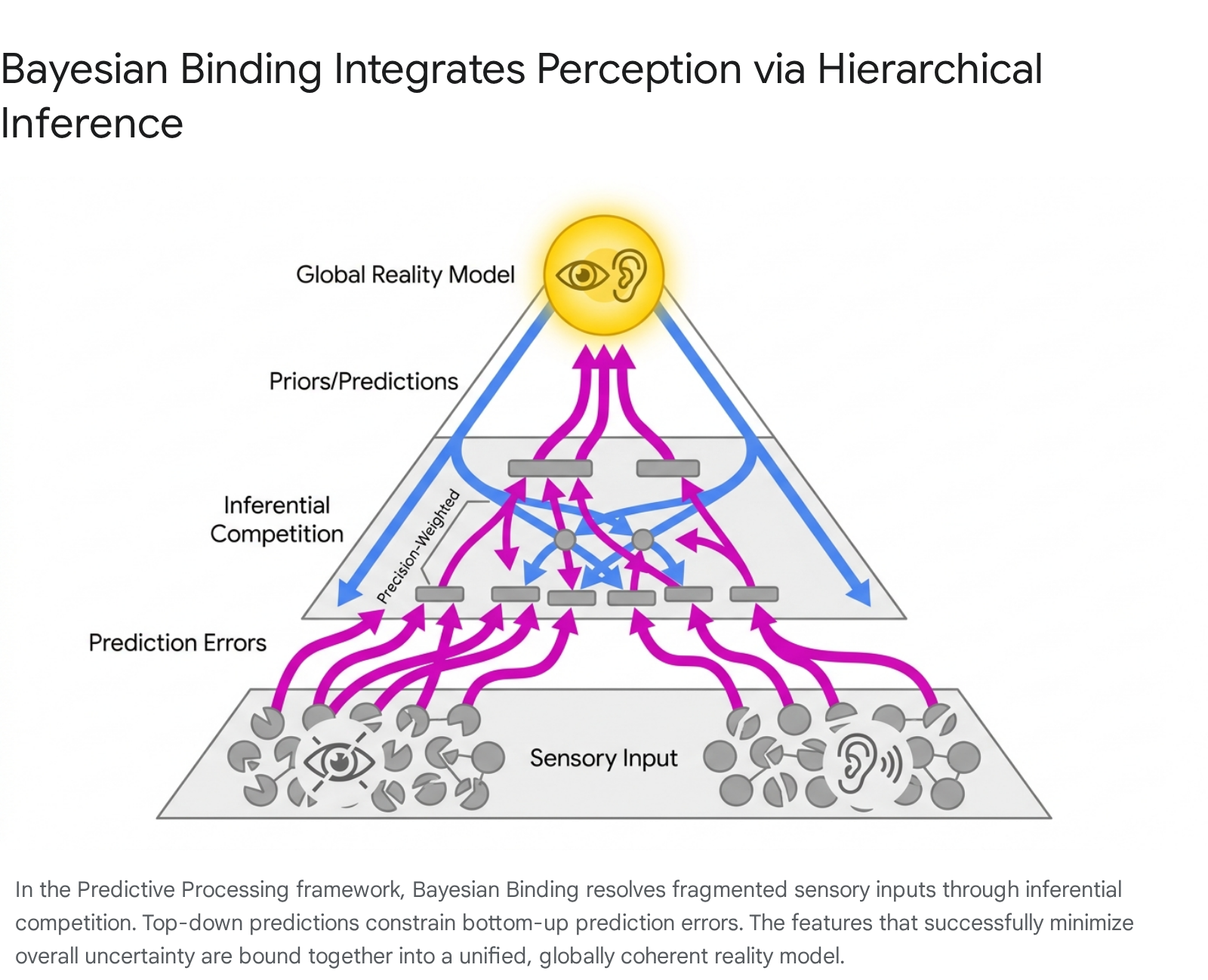
The global unity of a percept (e.g., binding the smell, shape, and color of a physical object) naturally emerges from a basic thermodynamic imperative to minimize prediction error; an internally incoherent perceptual field would exponentially accumulate uncertainty, paralyzing adaptive motor action 3738.
Bayesian Unbinding and Clinical Disruptions
Conversely, the deconstruction of this hierarchical framework results in "Bayesian unbinding" 33383941. By deliberately altering attention to lower the precision weighting of top-down predictions - such as through advanced meditative states prioritizing minimal phenomenal selfhood - the inferential competition fails to reach global coherence 3539. In the absence of top-down conceptual binding, sensory information remains raw, temporally volatile, and uncompressed, leading to a profound dissolution of the unified perceptual field 333841.
Pathological disruptions in predictive processing and Bayesian causal inference offer robust explanations for several severe psychiatric conditions. In schizophrenia, a failure to appropriately weight prediction errors can lead to a breakdown in the binding of self-generated actions to their sensory consequences, generating hallucinatory perceptions 283134. A deficit in extinction learning and erratic prediction error updating may generate the profound sense of "unreality" or dual-reality bookkeeping often observed in clinical delusions 34.
Despite its explanatory power, Predictive Processing faces significant criticism in philosophical circles. Critics argue that the framework is overly expansive, bordering on unfalsifiability, as almost any cognitive phenomenon can be post-hoc modeled as a form of error minimization 4031. Furthermore, phenomenological theorists argue that reducing all subjective experience to an extrinsic evolutionary cost-function fails to account for the intrinsic, subjective value of conscious feeling, leaving the "hard problem" of phenomenal binding untouched 42.
Theoretical Frameworks of Consciousness
The phenomenal binding problem is inextricably linked to the search for the Neural Correlates of Consciousness. Two dominant theories have historically provided competing, mutually exclusive explanations for how physical neural matter instantiates bound, conscious awareness: Global Neuronal Workspace Theory and Integrated Information Theory.
Global Neuronal Workspace Theory
Global Neuronal Workspace Theory posits that consciousness arises when highly processed sensory information is broadcast widely across the brain via a densely connected fronto-parietal network 384344. Under this model, binding is achieved through a sudden, nonlinear "ignition" - a widespread activation of neural coalitions, heavily reliant on the prefrontal cortex, which makes information globally available for working memory, verbal report, and action execution 384445. In this framework, binding is essentially a computational broadcasting event triggered when a threshold of activation is crossed.
Integrated Information Theory
Integrated Information Theory approaches the binding problem axiomatically, arguing that consciousness is a fundamental, intrinsic property of physical systems that possess a specific type of causal architecture. Consciousness exists precisely to the degree that a system is both highly differentiated (informative) and highly integrated (bound together), mathematically quantified as $\Phi$ 43565746.
Integrated Information Theory identifies the "posterior hot zone" (encompassing the occipital, temporal, and parietal cortices) as the anatomical substrate of maximal $\Phi$, demoting the prefrontal cortex to a secondary, non-essential role 445647. The theory resolves the phenomenal binding problem by asserting that a "complex" - the entity with maximum $\Phi$ - literally defines objective existence; individual micro-units within the complex cease to exist as independent entities and are intrinsically, metaphysically bound into a singular phenomenal state 1011.
Critics of Integrated Information Theory point to severe ontological difficulties, particularly the "dynamic entity evolution problem." They question how a bound self maintains psychological contiguity as the maximal complex physically shifts across the biological neural network over time 1011. Furthermore, critics challenge the theory's "intrinsicality 2.0 problem," arguing that by equating true existence solely with phenomenal existence, the theory erroneously excludes all unconscious, extrinsic physical entities from objective reality, bordering on idealism or unworkable panpsychism 574648.
The Cogitate Consortium Adversarial Collaboration
To break the theoretical deadlock between these frameworks, a massive open-science adversarial collaboration known as the Cogitate Consortium tested Global Neuronal Workspace Theory and Integrated Information Theory directly against each other. Published in 2025, the landmark study recorded brain activity from 256 human participants using functional magnetic resonance imaging, magnetoencephalography, and intracranial electroencephalography while they viewed suprathreshold stimuli for variable durations 44454749. The researchers preregistered strict, divergent predictions for both theories to eliminate confirmation bias 44.
The results were highly mixed, delivering substantial empirical challenges to key tenets of both overarching frameworks 44474950.
Global Neuronal Workspace Theory predicted a massive ignition in the prefrontal cortex at both the onset and offset of a conscious stimulus. While information could indeed be decoded in the inferior frontal cortex, there was a general lack of the predicted ignition at stimulus offset. Furthermore, the prefrontal cortex exhibited highly limited representation of certain conscious dimensions - such as specific stimulus categories or visual orientations - suggesting the prefrontal cortex is far less central to raw perceptual binding than the theory hypothesized 44474951.
Conversely, Integrated Information Theory predicted sustained, unbroken synchronization exclusively within the posterior hot zone for the entire duration a stimulus was consciously perceived. The rigorous data failed to show this sustained posterior synchronization, directly contradicting the core claim that continuous, static posterior network connectivity specifies ongoing conscious binding 4447. The only clear victory for GNWT over IIT was the finding of high-frequency oscillatory synchronization between early visual cortical regions and the front of the brain, a connection IIT predicted would not exist for simple visual awareness 49.
| Theory | Proposed Substrate of Binding | Primary Mechanism | Empirical Challenges (Cogitate 2025) |
|---|---|---|---|
| Global Neuronal Workspace Theory | Fronto-parietal network (emphasizing Prefrontal Cortex) | Nonlinear "ignition" and global computational broadcasting 4445. | Lack of offset ignition; poor decoding of specific stimulus categories in prefrontal cortex 444951. |
| Integrated Information Theory | Posterior hot zone (Occipital, Temporal, Parietal lobes) | Maximal $\Phi$ complexes defining intrinsic existence 115746. | Lack of sustained synchronization within posterior regions during continued perception 4447. |
| Predictive Processing (Bayesian) | Global hierarchical generative models | Precision weighting and inferential competition minimizing error 363738. | Debated falsifiability; struggles to address the purely subjective "hard problem" of experience 3142. |
Ultimately, the Cogitate study concluded that neither theory fully accounts for the neural realization of binding and consciousness 434750. The lack of a decisive victor underscores the sheer computational and anatomical complexity of phenomenal binding, suggesting that the integration of experience likely relies on transient, multi-regional, cross-frequency synchronizations that do not neatly fit either the pure prefrontal broadcasting model or the static posterior maximal-complex model.
Conclusion
The binding problem remains one of the most profound inquiries in contemporary neuroscience, spanning the molecular biophysics of single neurons to the highest levels of human consciousness. Extensive research demonstrates that the unification of separate sensory streams into a single experience is not achieved by a single, monolithic "Cartesian theater" in the brain. Instead, integration is a highly dynamic, scale-invariant process governed by strict statistical principles of spatiotemporal contiguity and inverse effectiveness.
Functional binding begins at the micro-level with non-linear dendritic summation in extreme hubs like the Mauthner cell and the superior colliculus. It ascends to the macro-level via cortico-thalamic pacing driven by the pulvinar nucleus and spatial coordinate anchoring in the posterior parietal cortex. The mechanism bridging these anatomical gaps is likely temporal synchrony - the transient phase-locking of distributed oscillations across cortical traveling waves. Ultimately, these synchronized signals are subjected to precision-weighted inferential competition. Through Bayesian Binding, the brain suppresses incoherent noise and elevates reliable predictions, weaving isolated sensory fragments into a single, highly optimized generative model of reality. While dominant theories of consciousness continue to aggressively debate the precise cortical boundaries of this integration, the physical and computational principles underlying multisensory unification provide a robust, increasingly clear map of how physical matter generates coherent perception.