Science of Human Handedness and Brain Lateralization
Human handedness - the preferential use of one upper limb over the other for motor tasks and object manipulation - represents one of the most visible and quantifiable manifestations of cerebral lateralization. Across millennia and irrespective of geographic boundaries, the human species has consistently exhibited a profound bias toward right-handedness. This population-level asymmetry, characterized by an approximate 90:10 ratio of right-handers to left-handers, has captivated evolutionary biologists, cognitive neuroscientists, and geneticists 123. The persistent survival of a distinct left-handed minority presents a complex evolutionary paradox: if handedness is heritable and left-handedness historically carried supposed fitness costs or subjected individuals to intense societal stigmas, the trait should have theoretically been extinguished by natural selection 45.
Modern neuroimaging, massive genome-wide association studies, and cross-species comparative zoology have fundamentally reshaped the contemporary understanding of manual lateralization. Research establishes that handedness is not merely a binary motor preference dictated by a single gene, but rather a highly complex, polygenic trait intimately linked with brain morphology, microtubule dynamics during embryogenesis, and androgen exposure in utero 367. Furthermore, the study of handedness provides a critical analytical window into the functional architecture of the brain, offering nuanced insights into how higher-order cognitive processes - particularly language, visuospatial attention, and executive control - are distributed across the cerebral hemispheres 8910.
Epidemiology and the Measurement of Handedness
Establishing the exact global prevalence of left-handedness has historically been complicated by varying definitions of manual preference, ranging from the strict identification of the writing hand to broad assessments of cross-dominance in multiple unimanual and bimanual tasks. However, comprehensive modern meta-analyses have provided highly calibrated statistical estimates that isolate the true phenotypic distribution.
Global Prevalence Rates and Methodological Variance
A landmark 2020 meta-analysis incorporating 262 datasets from 200 published studies, which encompassed over 2.3 million individuals globally, established that the baseline prevalence of left-handedness is approximately 10.6% 111213. The data highlights that variance in reported prevalence rates across the scientific literature is heavily dependent on the stringency of the measurement criteria utilized by researchers. When utilizing the most stringent criterion of exclusive left-handedness across all queried motor tasks, global prevalence falls to roughly 9.34% 11. Conversely, when utilizing the most lenient criterion of "non-right-handedness" - which includes varying degrees of mixed-handedness, cross-dominance, and ambidexterity - the prevalence rate rises to 18.1% 1112.
Cross-dominance, or mixed-handedness, wherein individuals switch their preferred hand depending on the specific task (e.g., writing with the left hand but throwing with the right), accounts for nearly 9% of the population 111. Natural ambidexterity, defined as equal and interchangeable proficiency with both hands across a comprehensive spectrum of tasks, remains exceptionally rare, with an estimated global prevalence of roughly 1% 1. Furthermore, demographic analyses consistently reveal a distinct sexual dimorphism in manual preference: men are approximately 25% more likely to be left-handed than women, resulting in a male-to-female ratio of roughly 5:4 across large cohorts 1415.
Regional Disparities and Cultural Pressures
While the global average stabilizes near 10.6%, regional disparities are highly pronounced. Epidemiological studies consistently indicate that left-handedness is most prevalent in North America, Western Europe, Australia, and New Zealand, where rates frequently reach 10% to 13.1% 131617. Conversely, significantly lower rates are observed in parts of Asia, Africa, and South America, often hovering between 2% and 6% 1317. Within specific nations, internal regional variations also exist; for example, data from Italy indicates higher rates of left-handedness in the northern provinces compared to the south 16.
These geographical fluctuations are largely attributed to societal and cultural mechanisms rather than localized genetic divergence. In many traditional societies, the left hand is historically associated with personal hygiene and is heavily stigmatized in contexts of greeting, eating, or writing 11217. Consequently, children exhibiting natural left-handedness are frequently subjected to forced hand-switching by parents and educators 117. Anthropological frameworks utilizing the Power Distance Index (PDI) to evaluate cultural structures demonstrate that high-PDI cultures - which emphasize rigid conformity and formal adherence to majority behavioral patterns - exhibit the lowest rates of phenotypic left-handedness. Sixty-eight percent of countries categorized as formal cultures are located in regions displaying the lowest prevalence 17.
| Nation | Estimated Left-Handed Population (Millions) | Total Population (Millions) | Implied Prevalence Rate | Regional Context |
|---|---|---|---|---|
| Netherlands | 2.4 | 18.3 | ~13.1% | Western Europe (High tolerance, non-formal) |
| United States | 44.9 | 342.4 | ~13.1% | North America (High tolerance, non-formal) |
| France | 7.7 | 69.0 | ~11.1% | Western Europe (High tolerance, non-formal) |
| Japan | 5.8 | 123.9 | ~4.7% | East Asia (Historical forced switching, high conformity) |
| Vietnam | 4.1 | 98.0 | ~4.26% | Southeast Asia (Formal culture, conformity pressures) |
| China | 49.5 | 1,410.0 | ~3.5% | East Asia (Formal culture, complex writing systems) |
Large-scale demographic tracking in the United States using the Gilbert and Wysocki data further highlights ethnic variations influenced by ancestral cultural backgrounds. While White, Black, and Native American demographics exhibited historical trendlines mirroring the national average, individuals of Asian and Hispanic descent showed lower overall rates of left-handedness and a slower rate of phenotypic increase over the 20th century, compatible with cultural norms emphasizing right-handed writing 14.
Historical Fluctuations and Demographic Trends
Historical and archaeological data further corroborate the profound influence of societal attitudes on the phenotypic expression of handedness. Archaeological evidence - such as the prevalence of right-hand stencils in Upper Paleolithic cave art (e.g., Cueva de las Manos) and right-handed wear patterns on ancient Pleistocene tools - indicates that the overwhelming right-handed majority has existed for tens of thousands of years 12416.
However, modern demographic history reveals significant short-term fluctuations. Analyses of birth cohorts from the 19th and 20th centuries show a precipitous drop in left-handedness during the Victorian era. Among individuals born between 1880 and 1900, the rate of left-handedness fell to an anomalous nadir of roughly 3% to 4% 1416. The prevalence subsequently rose steadily throughout the first half of the 20th century, reaching an asymptote of 11% to 12% for birth cohorts post-1950 14.
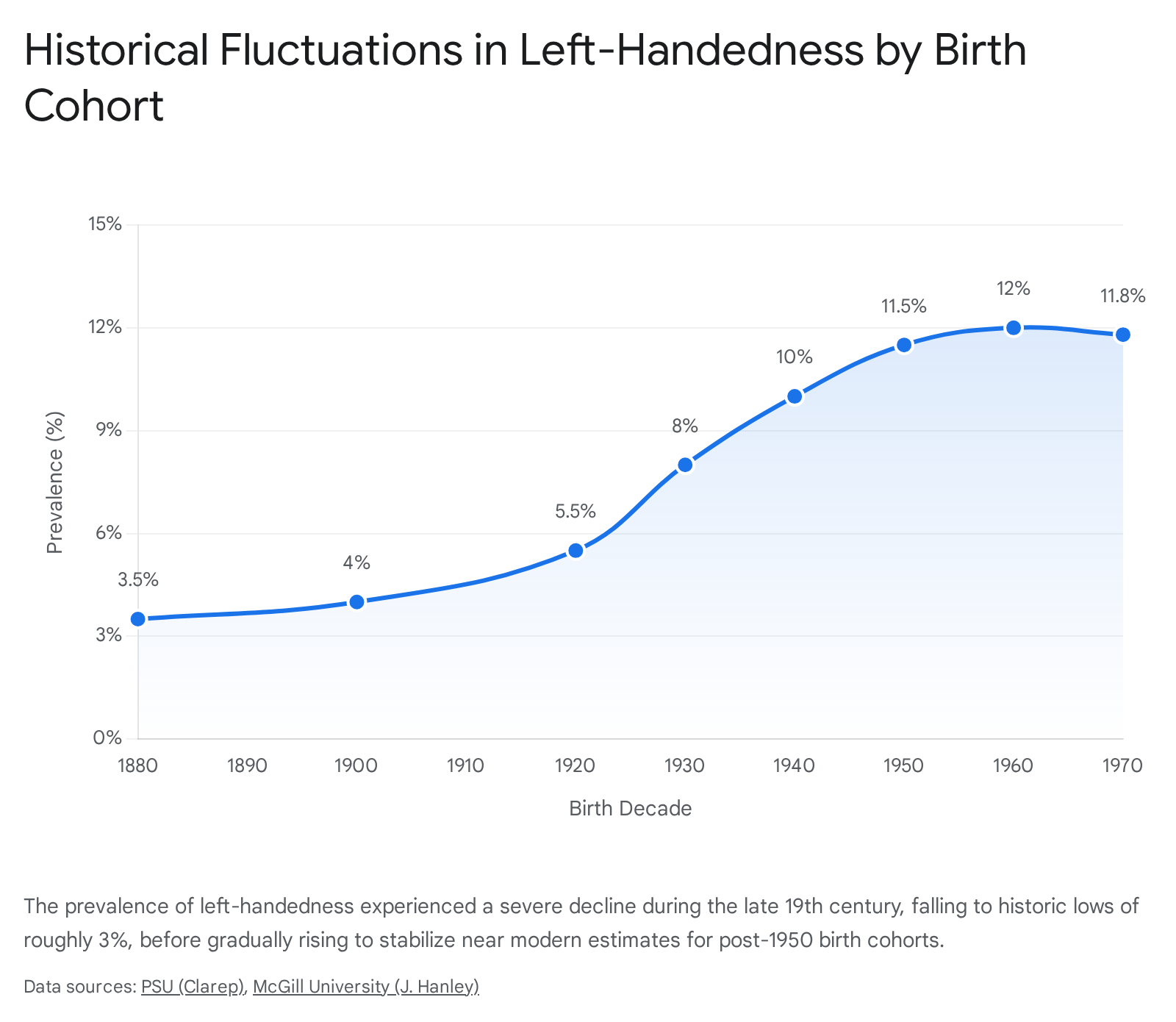
Researchers attribute this historical dip to intense social ostracization. During the 19th century, left-handedness was heavily penalized, which not only led to forced hand-switching but also impacted sexual selection and human mating dynamics 16. Historical data suggests that left-handers became less desirable as mating partners due to social stigmatization. Furthermore, couples where both partners were left-handed exhibited lower reproductive rates during this period (averaging 2.3 children) compared to right-handed couples (averaging 3.1 children), presumably due to a reluctance to subject offspring to systemic societal prejudice 16. The subsequent 20th-century rebound directly correlates with the gradual relaxation of these rigid societal norms, allowing the genetic propensity for left-handedness to express itself phenotypically without immediate reproductive or social penalties 16.
Evolutionary Biology and the Persistence of Left-Handedness
The persistence of the left-handed polymorphism demands an evolutionary explanation. Given that left-handedness is heritable, any trait that constitutes a consistent minority must offer a selective advantage in specific contexts; otherwise, it would either be driven to extinction by natural selection or drift to fixation 2518.
The Original Fighting Hypothesis
In 1996, researchers proposed the "fighting hypothesis," postulating that left-handedness is maintained in the human population through negative frequency-dependent selection 2419. This evolutionary model suggests that rare traits provide advantages precisely because they are rare. In the context of early hominin physical combat, right-handers (comprising the vast majority) were accustomed to fighting other right-handers. Conversely, a left-handed combatant possessed a distinct tactical element of surprise, as their striking angles and defensive postures were unfamiliar to the right-handed majority 45.
The primary evidence supporting this hypothesis derives from modern interactive sports, which serve as an observable proxy for ancient combat. In sports requiring direct, high-speed interaction and spatial calculation - such as fencing, boxing, tennis, and baseball - left-handers are vastly overrepresented, occasionally comprising up to 40% to 50% of the elite sporting echelons 121920. This tactical advantage diminishes only when left-handers become too common in a local population, at which point right-handers adapt to their fighting style, thereby stabilizing the polymorphism at a distinct minority ratio 1821. Furthermore, the fighting hypothesis provides a plausible explanation for the sex difference in handedness: men are roughly 25% more likely to be left-handed than women, aligning with the evolutionary reality that intra-sexual physical combat has historically been overwhelmingly male-dominated 241419.
Evaluation of Health Costs and Fitness Penalties
The original fighting hypothesis relied on a critical secondary assumption: that left-handedness carried inherent physiological or neurodevelopmental "health costs" (e.g., reduced longevity, increased susceptibility to autoimmune disorders, or elevated psychopathology), which kept the trait from expanding beyond roughly 10% despite its combat advantages 522. Early literature heavily suggested that left-handedness possessed Darwinian fitness costs that acted as a counterbalance to the fighting advantage 52123.
However, exhaustive modern meta-analyses have largely dismantled these supposed health penalties. Studies encompassing tens of thousands of participants reveal no reliable links between consistent left-handedness and reduced lifespan, diminished fertility, or increased baseline disease profiles 522. A massive meta-analysis evaluating handedness and depression, which aggregated data from 87 studies and over 35,000 participants, found zero evidence linking left-handedness to increased levels of depression 22. While some data indicate minor increases in cognitive deficits or health issues among mixed-handers or ambidextrous individuals (potentially due to pathological origins of their atypical handedness), consistent left-handers are objectively as healthy as consistent right-handers 522. The absence of these health costs in both industrial and non-industrial societies significantly weakens the original framework of the hypothesis 522.
The Modified Fighting Hypothesis
The lack of a confirmed health cost leaves a profound theoretical gap: if left-handers win more fights and suffer no evolutionary health penalties, why did they not become the majority? To resolve this, researchers proposed the Modified Fighting Hypothesis in 2023, shifting the focus from the minority left-handers to the biomechanical vulnerabilities of the majority 522.
This revised evolutionary model focuses on cardiovascular anatomy during armed combat among early hominins. Approximately 75% of the human heart, and the critical ascending aorta, is located in the left hemithorax 2224. In mortal combat utilizing sharp, stabbing implements, a right-handed unilateral grip naturally rotates the vulnerable left side of the chest away from the opponent, presenting the less critical right hemithorax. Conversely, a left-handed fighter inherently exposes their vital left hemithorax directly to their adversary's weapon 52224.
Under this model, right-handedness became the overwhelming population-level baseline because it offered a profound, immediate survival advantage by shielding the cardiovascular system from mortal wounds in early human history. Left-handedness persists today solely because, as a rare phenotype, its frequency-dependent tactical surprise advantage in physical conflicts manages to counterbalance its inherent, fatal anatomical vulnerability 522.
Genetic Architecture and Gestational Mechanisms
Historically, theoretical models attempted to explain handedness through simple Mendelian inheritance or single-gene alterations, such as the Right-Shift theory or the Geschwind-Behan-Galaburda (GBG) model 125. Modern genomic research has rendered these perspectives obsolete, demonstrating that handedness is a highly complex, polygenic trait shaped by early developmental environments and cellular mechanics.
Genome-Wide and Exome-Wide Associations
Recent breakthroughs utilizing massive datasets from the UK Biobank, 23andMe, and the International Handedness Consortium have isolated the distinct genetic architecture of manual preference. A 2020 genome-wide association (GWAS) meta-analysis evaluating 1,766,671 individuals identified 41 distinct common genetic loci significantly associated with left-handedness and 7 loci associated with ambidexterity 626. Crucially, the genetic correlation between left-handedness and ambidexterity is remarkably low ($r_G = 0.26$), indicating that they are fundamentally distinct phenotypes driven by disparate genetic mechanisms rather than varying degrees of a single lateralization continuum 626.
Tissue-enrichment analyses of these loci heavily implicate the central nervous system, with specific enrichment in genetic pathways regulating brain morphology and microtubule dynamics 626. Subsequent 2024 exome-wide association scans analyzing rare protein-altering variants (frequencies $\le 1\%$) isolated specific genes, notably the beta-tubulin gene TUBB4B and the microtubule-associated protein MAP2 272829. In the UK Biobank exome data from 351,314 individuals, the TUBB4B gene showed an exome-wide significant association, with a rate of rare coding variants 2.7 times higher in left-handers than right-handers, including specific frameshift mutations found exclusively in left-handed cohorts 29.
Microtubules constitute the cellular cytoskeleton and play a pivotal role in neuronal migration, axonal growth, and the function of primary cilia during embryogenesis. Researchers posit that early left-right asymmetrical development in the human embryo is driven by fluid flow generated by microscopic ciliary motion; thus, minute genetic variations in tubulin structures may shift the organism's baseline asymmetrical development, subtly altering brain laterality and manual preference 62829.
Furthermore, while the exome-wide heritability of left-handedness due to rare coding variants was found to be just 0.91% (with general heritability across all variants estimated around 24%), the genes identified overlap with neurodevelopmental profiles 129. Variants in genes such as DSCAM and FOXP1, previously implicated in autism spectrum disorder and schizophrenia via exome screening, show evidence for rare coding variant associations with left-handedness 29. However, researchers emphasize that these genetic links are highly polygenic and non-deterministic, functioning as broad indicators of atypical neurodevelopment rather than direct, causal pathological linkages 626. Previous theories linking visceral asymmetry genes to handedness have also been largely disproven in the general population 27.
The Twin Testosterone-Transfer Hypothesis
Beyond pure genomics, the intrauterine environment exerts a profound influence on manual lateralization, primarily through fetal hormone exposure during critical developmental windows. The "twin testosterone-transfer hypothesis" provides compelling, large-scale evidence for this mechanism 3031. In human gestation, fetuses are exposed to varying hormonal environments based on the sex of their co-twins due to hormone diffusion through shared amniotic fluid or maternal bloodstreams 730.
A comprehensive study analyzing 4,736 Finnish twins (representing roughly 70% of all twins born in Finland between 1983 and 1987) measured the prevalence of left-handedness in females sharing the womb with a female co-twin versus those sharing the womb with a male co-twin 73132. The data revealed that females from opposite-sex pairs exhibited a significantly lower prevalence of left-handedness (5.3%) compared to females from same-sex pairs (8.6%) 732. This outcome occurs because the female fetus in a male-female pairing is exposed to elevated levels of androgens - specifically testosterone - produced by the male co-twin 73031.
These findings suggest that prenatal testosterone exposure directly modulates the neurodevelopmental pathways dictating motor lateralization 73132. While testing circulating hormone levels in these subjects at age 14 showed no lingering differences, the prenatal exposure leaves an indelible mark on brain organization 732. Although the exact neurobiological cascade remains partially obscured, the data strongly align with theories positing that intrauterine masculinization affects hemispheric growth rates, subsequently shifting the probabilistic outcome of handedness toward right-hand preference in females exposed to higher testosterone 731.
Neuroanatomy and Hemispheric Specialization
Handedness serves as an observable behavioral proxy for the invisible, large-scale functional organization of the brain. The prevailing rule in cognitive neuroscience is contralateral motor control: the left hemisphere dictates the motor functions of the right side of the body, and the right hemisphere controls the left 13. However, the relationship between handedness and higher-order cognitive lateralization - specifically language processing, spatial attention, and executive function - is highly nuanced and anatomically complex.
Language Dominance and Atypical Lateralization
Language processing is historically recognized as the most highly lateralized cognitive function in the human brain. Functional magnetic resonance imaging (fMRI) reveals that in 94% to 96% of right-handers, the primary language hubs (such as Broca's area in the inferior frontal gyrus and Wernicke's area in the posterior temporal lobe) are heavily localized to the left hemisphere 91033. Because the vast majority of the human population is both right-handed and left-hemisphere dominant for language, early neurological dogma assumed the two traits were inextricably hardwired by a single genetic mechanism 1.
However, the neural architecture of left-handers displays significantly greater variance. While the majority of left-handers (approximately 70% to 80%) maintain typical left-hemisphere dominance for language, they exhibit much higher rates of "atypical lateralization" than their right-handed counterparts 333536. Meta-analyses of whole-brain fMRI studies indicate that roughly 20% to 30% of left-handers process language atypically, either bilaterally (utilizing both hemispheres relatively equally) or with distinct right-hemisphere dominance 13637.
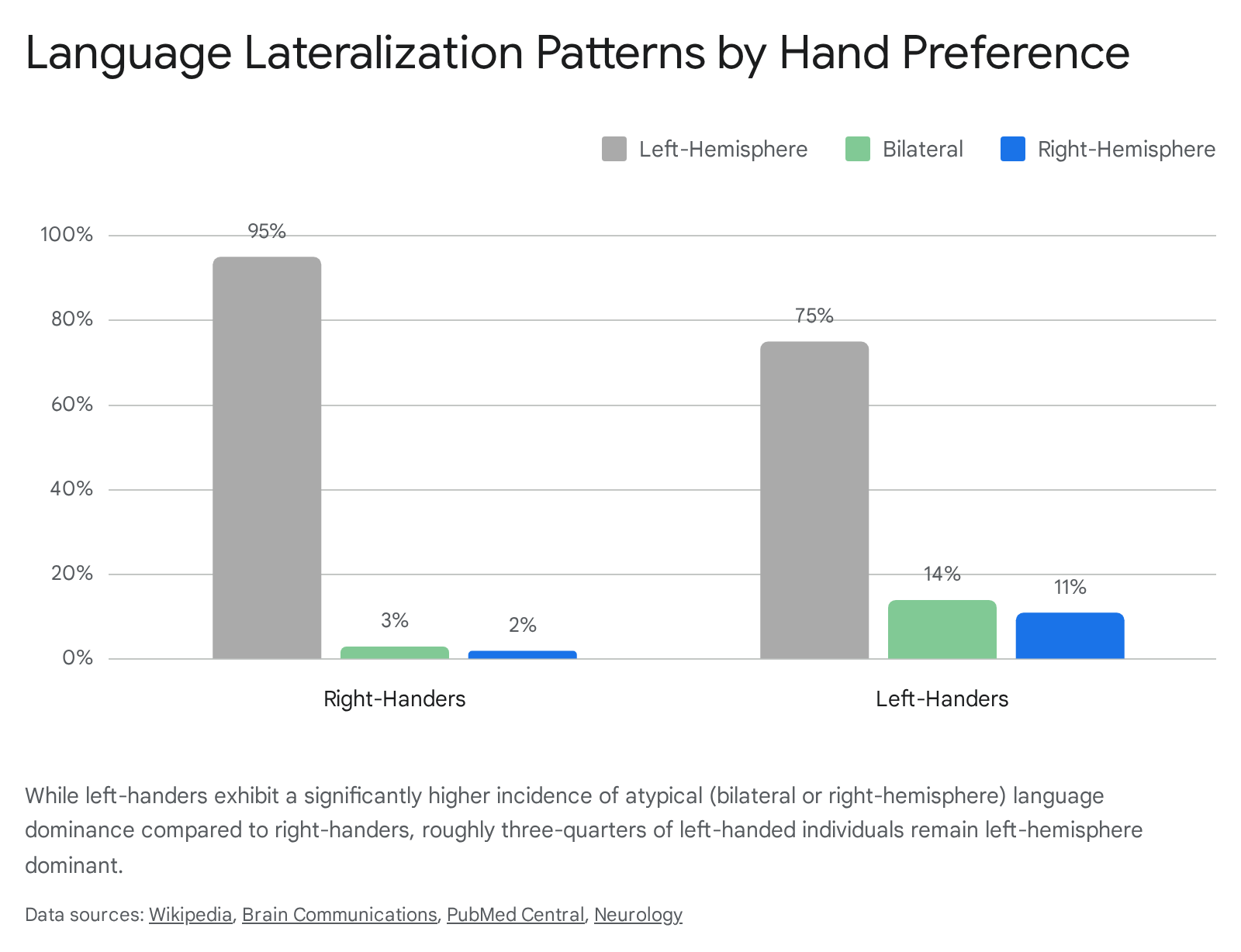
Crucially, atypical language organization is not merely a pristine mirror image of standard neuroanatomy 10. In individuals with bilateral or right-dominant language profiles, functional processing is often more spatially diffuse. During verbal fluency tasks, these individuals frequently recruit right-hemisphere components of the default mode network (DMN), including the angular gyrus and middle temporal gyrus, which are rarely engaged by typically lateralized individuals during similar tasks 1037.
Developmental studies utilizing large pediatric cohorts (273 children ages 4-16) demonstrate that strong left-hemisphere language bias in activation volume and response magnitude is fully evident by age 4 38. This confirms that atypical lateralization in certain individuals is a distinct neurodevelopmental trajectory laid down in early childhood, rather than a generalized delay in lateralization that solidifies later in life 38.
Despite these differences in spatial organization, comprehensive cognitive testing reveals no broad relationship between left-handedness and diminished cognitive efficiency in the general population . However, within the highly specific sub-population of left-handers possessing strongly atypical right-hemisphere language lateralization, functional connectivity studies suggest moderately poorer overall performance in tasks requiring rapid processing speeds and extensive interhemispheric connectivity, such as spatial processing and verbal reasoning under temporal pressure 936.
Structural Morphology and the Corpus Callosum Debate
The search for macroscopic structural biomarkers of handedness has historically focused on the corpus callosum (CC), the massive white-matter tract connecting the two cerebral hemispheres. Originating from early morphometric studies in the 1980s and 1990s, a widely entrenched neuroscientific belief held that left-handers and mixed-handers possessed a larger or thicker corpus callosum than consistent right-handers, theoretically facilitating greater interhemispheric cross-talk to accommodate their atypical lateralization profiles 3435.
However, the advent of massive neuroimaging datasets and rigorous analytical controls has fundamentally challenged this consensus. A definitive 2022 meta-analysis utilizing data from 1,057 participants in the Human Connectome Project (HCP 1200) achieved unprecedented spatial resolution by analyzing 100 specific measuring points along the midsagittal corpus callosum 34. By strictly controlling for forebrain volume (FBV) as a covariate and comparing groups based on both the consistency and direction of hand preference, the researchers found no statistically significant differences in the macroscopic size, relative area, or subregional thickness of the corpus callosum between left- and right-handers 34. The calculated effect sizes were exceptionally small (Cohen's d < 0.1), suggesting that historical findings of larger CC volumes in left-handers were likely false positives driven by underpowered sample sizes and lower-resolution segmentation strategies 34.
Conversely, microstructural analyses utilizing Diffusion Tensor Imaging (DTI) present a slightly different, highly localized picture. Studies measuring fractional anisotropy (FA) - a sensitive metric of white-matter microstructural integrity, water diffusion, and axonal coherence - indicate subtle differences 353637. Some high-resolution DTI reports find slightly increased FA and diminished mean diffusion across all callosal subregions in left-handers compared to right-handers 35. This suggests that while the macroscopic perimeter and volume of the structure are identical, the molecular diffusion properties and internal axonal packing within the tract may possess subtle, handedness-related microstructural variations 3538.
Cortical Asymmetry, Thalamic Morphometry, and Clinical Plasticity
On the broader cortical surface, large-scale structural MRI analyses investigating tens of thousands of participants (N = 31,864) have identified widespread, albeit slight, grey matter volume (GMV) asymmetries. Right-handers and left-handers differ in average brain asymmetry across ten specific cortical regions associated with working memory, language, and executive hand control 3940. In these regions, left-handers generally exhibit relatively larger GMV in the right hemisphere, anatomically reflecting the right hemisphere's dominant functional role in controlling their preferred left limb 39.
Subcortical structural variability has also been linked to manual preference. Automated parcellation of thalamic nuclei using test-retest fMRI datasets reveals interhemispheric differences in specific regions - namely the ventral anterior and ventral posterolateral thalamic nuclei - between left- and right-dominant individuals, highlighting that motor lateralization relies heavily on deep-brain relay mechanisms 41.
The flexibility of these lateralized structures is best demonstrated in clinical pathology and specialized learning. In individuals with chronic stroke-induced aphasia, voxel-based morphometry (VBM) reveals that recovery of language function is often mediated by the contralesional right hemisphere. Aphasic patients demonstrate that increased right-hemisphere GMV in the middle temporal gyrus (MTG) correlates with improved language comprehension, indicating that the right hemisphere can support functions previously performed by damaged left-hemisphere hubs 42. Similarly, studies of sequential bilingual individuals demonstrate structural adaptations to lateralization, such as a steeper age-related GMV decline in the left inferior parietal lobule (IPL) alongside higher overall bilateral IPL volume, acting as a form of "bilingual brain reserve" 43.
Deconstructing Neuromythology
The comparative rarity of left-handedness, combined with early rudimentary misunderstandings of functional brain lateralization, spawned numerous cultural and scientific myths that persisted in the public consciousness for decades. Modern empirical scrutiny has systematically deconstructed these narratives.
Creativity and Divergent Thinking
A pervasive modern stereotype holds that left-handed individuals are innately more creative, artistic, and innovative. This belief stems from the accurate neurological observation that divergent thinking - the capacity to rapidly explore multiple, non-linear solutions to a problem and forge unexpected conceptual connections - is heavily supported by processing hubs within the brain's right hemisphere 4445. Because left-handers utilize the right hemisphere to control their dominant hand, popular psychology deduced a direct, biological link to enhanced overall creativity.
A comprehensive 2025 meta-analysis published in the Psychonomic Bulletin and Review systematically dismantled this claim. Researchers sifted through nearly 1,000 scientific papers published over more than a century regarding handedness and creative output, ultimately utilizing rigorous laboratory tests of divergent thinking 444551. The data revealed no consistent advantage in creative thinking for left-handers. In standardized metrics, such as the Alternate Uses Test, handedness made virtually no difference; when statistically significant deviations did occur, right-handers frequently demonstrated a slight advantage 444551.
The endurance of the "creative lefty" myth is largely attributed to statistical coincidence and confirmation bias. Because both extreme creative genius and left-handedness are relatively rare (affecting roughly 10% of the population), overlapping occurrences are highly memorable and culturally romanticized as the "tortured artist" archetype 444551. While left-handers may be anecdotally overrepresented in specific artistic and musical fields, broad empirical data reveals that right-handers dominate professions requiring the highest objective, standardized metrics of creativity 444551.
The Left-Brain Versus Right-Brain Fallacy
The creativity myth is a sub-component of a much larger neuromyth: the concept of binary "brainedness" or the "left-brain vs. right-brain" personality dichotomy 2535. Popular culture aggressively characterizes "left-brained" individuals as logical, scientific, and analytical, while painting "right-brained" individuals as creative, artistic, and intuitive - routinely correlating left-handedness with the latter profile 2535.
This simplistic paradigm was definitively debunked by a rigorous 2013 neuroimaging study conducted at the University of Utah. Utilizing resting-state functional connectivity magnetic resonance imaging (rs-fcMRI), researchers analyzed the brains of 1,011 individuals between the ages of seven and 29. The team divided each cortex into over 7,000 localized regions to map lateralized connectivity networks 35. The analysis proved that while specific functions (like language production or spatial attention) are lateralized to one hemisphere, whole-brain networks are not. Individuals absolutely do not exhibit a globally stronger, more active, or more highly connected left- or right-hemisphere network. Consequently, psychological personality profiles are entirely divorced from global hemispheric dominance, rendering the concept of "right-brained" individuals scientifically obsolete 2535.
Furthermore, attempts to link handedness to specific speech pathologies, such as stuttering, have also failed to produce evidence of atypical global lateralization. Functional neuroimaging meta-analyses of people who stutter (PWS) versus typical fluent speakers (TFS) reveal no significant difference in the distribution of handedness between the groups, nor does handedness alter the typical pattern of left-hemispheric cerebral dominance for speech in individuals who stutter 46.
Comparative Lateralization in Non-Human Species
For decades, robust population-level handedness was considered a uniquely human trait, theorized to have evolved synchronously with human-specific evolutionary milestones such as advanced tool making, obligatory bipedalism, and syntactic language 3114748. However, advancements in comparative zoology and behavioral ecology have radically altered this anthropocentric view, establishing that behavioral and morphological lateralization is a ubiquitous feature of vertebrate neural organization 484950.
Lateralization Across Diverse Vertebrate Orders
A comprehensive 2025 meta-analysis updating data across all non-extinct vertebrate orders analyzed 172 different species for limb and motor preferences 4951. The integration of robust empirical evidence revealed that 39.53% of studied vertebrate species show clear evidence of population-level asymmetries (where the vast majority of the species prefers one side), 32.56% show individual-level asymmetries (where individuals are highly lateralized, but the population is split 50/50), and 27.91% show no asymmetry 4951.
| Taxonomic Group | Examples | Prevailing Asymmetry Type | Specific Findings |
|---|---|---|---|
| Primates | Chimpanzees, Spider Monkeys | Individual-Level | Strong individual preference; ~50/50 population split. Arboreal species display stronger lateralization than terrestrial species. |
| Carnivora | Domestic Cats, Dogs | Individual-Level | 78% of cats and 68% of dogs show individual preference. Female cats have higher odds of right-paw preference. |
| Squamata | Geckos | Individual-Level | Strong individual preference; ~50/50 population split. No population-level asymmetry. |
| Marsupialia (Bipedal) | Red Kangaroos, Wallabies | Population-Level (Left) | Up to 95% left-forelimb preference for unimanual tasks (feeding, grooming). |
| Marsupialia (Quadrupedal) | Sugar Gliders, Opossums | No Asymmetry | Ambidextrous motor profiles; no population or strong individual bias. |
Table 1: Summary of limb preference and asymmetry across various non-human taxa based on recent meta-analyses 4749515852.
Primate Phylogeny and the Uniqueness of Human Handedness
In an attempt to trace the direct phylogenetic origins of human right-handedness, zoologists have heavily analyzed our closest evolutionary relatives. A massive 2022 comparative study collected standardized manual manipulation data (the "tube task") from 1,786 individuals across 38 species of anthropoid primates 475361.
The results indicated that while many non-human primates display strong individual-level lateralization, population-level uniformity is exceedingly rare 4761. In species like spider monkeys, the ratio of left-handed to right-handed individuals hovers near an equal distribution. The phylogenetic analysis confirmed that human lateralization patterns do not align with trends found among other anthropoids, confirming that the universal ~90% right-handedness seen in Homo sapiens is an unparalleled evolutionary extreme, likely driven by unique selective pressures in the hominin lineage rather than a shared primate inheritance 8475361. Interestingly, the study noted that ecological environment dictates the strength of hand preference: arboreal primates require complex, stabilizing limb coordination and exhibit stronger individual lateralization, whereas terrestrial primates tend to display weaker overall hand preferences 4753.
Marsupial Bipedalism and Population-Level Asymmetry
While primates fail to mirror human population-level lateralization, recent discoveries in Australian marsupials have unveiled striking, convergent parallels. A groundbreaking 2015 study published in Current Biology demonstrated that multiple species of bipedal marsupials exhibit extreme population-level left-handedness 58525455.
In comprehensive field observations, bipedal species such as eastern grey kangaroos, red kangaroos, red-necked wallabies, and brush-tailed bettongs displayed a distinct left-hand preference in up to 95% of recorded instances for unimanual tasks, such as feeding, grooming, and manipulating objects 525556. Conversely, closely related quadrupedal marsupials - such as the sugar glider, the gray short-tailed opossum, and the arboreal Goodfellow's tree-kangaroo - exhibit ambidextrous profiles with no population-level handedness 58525456.
This stark dichotomy among marsupial families strongly suggests that posture and gait, specifically the evolutionary shift to obligate bipedalism, act as potent catalysts for the development of population-level limb lateralization 5855. Because the marsupial brain entirely lacks a corpus callosum - relying instead on alternative commissural pathways for interhemispheric communication - their development of pronounced, population-wide handedness proves that the emergence of lateralized motor control is an ancient, convergent evolutionary strategy deeply tied to bipedal biomechanics, rather than a unique byproduct of placental mammalian neuroanatomy 345255.