Quantum biology in photosynthesis bird navigation and smell
Foundations of Quantum Biological Systems
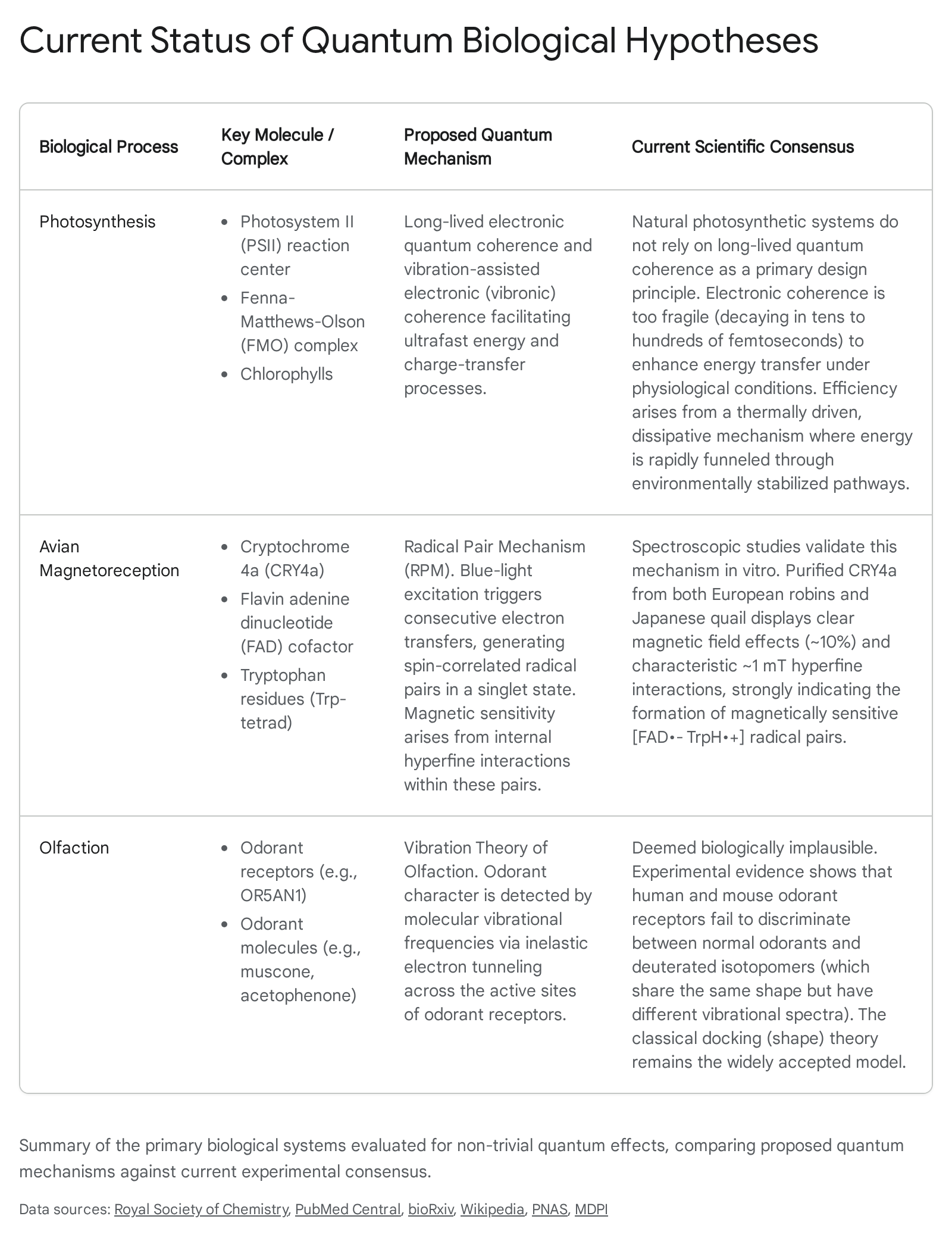
The intersection of quantum mechanics and biological sciences has historically been viewed through the lens of trivial quantum effects. Under this conventional framework, quantum mechanics is acknowledged as the underlying foundation of all molecular structures, governing chemical bonding, electron orbitals, and the structural conformations of macromolecules 12. However, the emerging discipline of quantum biology focuses on non-trivial quantum effects. These involve phenomena such as quantum superposition, entanglement, and quantum tunneling operating dynamically to fulfill specific biological functions 134. Since the early speculations of physicists like Erwin Schrödinger, who proposed that quantum fluctuations produce mutations, and Niels Bohr, who considered complementarity in living systems, the question of whether macroscopic life exploits quantum mechanics has remained controversial 4.
The primary theoretical obstacle to non-trivial quantum biology is the phenomenon of quantum decoherence. Quantum states are notoriously fragile; when a quantum system interacts with a macroscopic environment, the phase relationships that characterize quantum coherence are rapidly destroyed 345. In the cold, highly isolated environments of laboratory quantum systems, such as superconducting qubits or trapped ions cooled to near absolute zero, coherence can be maintained for extended periods 36. Biological systems, conversely, operate in warm, wet, and highly complex environments, typically at or near 300 Kelvin 7. Standard physical models predict that thermal fluctuations in such environments should cause quantum decoherence on the order of femtoseconds ($10^{-15}$ seconds) to picoseconds ($10^{-12}$ seconds) 45. This timescale is generally assumed to be too fast to influence slower biochemical reactions. For instance, early modeling suggested that decoherence times for a nucleotide in a cellular environment at room temperature would be less than $10^{-13}$ seconds 4.
Despite these theoretical constraints, experimental data utilizing ultrafast multidimensional spectroscopy and advanced molecular biology have repeatedly suggested that certain biological complexes can sustain and exploit quantum coherence 28. This raises a fundamental biophysical question: do living organisms possess structural mechanisms capable of shielding quantum states from environmental decoherence, or do they actively utilize environmental noise to assist quantum transport?
Mechanisms of Environmental Shielding
Recent biophysical research has identified several mechanisms by which biological structures might mitigate rapid decoherence. Unlike static laboratory qubits, biological molecules are dynamic, open systems operating far from thermodynamic equilibrium 5. The assumption that the environment acts merely as a destructive thermal bath is being replaced by models in which the environment is structurally integrated into the quantum dynamics.
One primary protective mechanism is quasi-isolation through hydrophobic pockets. Within specific proteins, functional reaction centers are often buried in non-polar regions that exclude water molecules, thereby restricting the degrees of freedom available for interaction with the surrounding thermal bath 510. Additionally, the structured arrangement of water molecules and specific ions near protein surfaces may act as a screening layer, altering the spectral density of the environmental noise 56. Within highly structured environments like microtubules, it has been proposed that layers of ordered water, gel states, and counterion shielding might screen thermal fluctuations, though the efficacy of this shielding for macroscopic quantum computation remains highly contested 56. Studies of specific putative quantum processors, such as Posner molecules (entangled calcium phosphate clusters), have demonstrated that entanglement in these asymmetric ensembles decays on a subsecond timescale, rendering them unlikely candidates for long-term neuronal processing 11.
Furthermore, biological systems may utilize noise-assisted transport. Rather than environmental fluctuations purely destroying quantum phase, specific frequencies of thermal noise can continuously bump a quantum state out of localized traps, effectively driving energy transfer forward 7. In this regime, the system is dynamically driven away from equilibrium, and the interplay between the system and its environment creates decoherence-free subspaces 58. Evolution appears to have optimized certain biomolecular architectures to sit at the "quantum edge," balancing speed, energy dissipation, and quantum coherence to achieve operational efficiencies impossible under purely classical regimes 58.
Photosynthetic Excitation Energy Transfer
Photosynthesis represents one of the most highly efficient energy conversion processes in nature, successfully capturing solar photons and routing the resulting excitation energy to reaction centers with near-unity quantum efficiency 91011. The initial step of this process relies on light-harvesting antenna complexes, such as the Fenna-Matthews-Olson (FMO) complex in green sulfur bacteria and Photosystem II (PSII) in higher plants and cyanobacteria 11121314.
The classical framework for understanding this energy transport is Förster resonance energy transfer (FRET), which models the process as incoherent, stepwise hopping of localized excitons between pigment molecules, driven by a gradient of site energies leading to the reaction center 121415. However, the extreme speed and efficiency of this transfer led physicists to investigate whether excitons might exist in a state of quantum superposition, exploring multiple energetic pathways simultaneously to find the optimal route through the complex energy landscape 111516.
The Electronic Coherence Hypothesis
The hypothesis of quantum-enhanced photosynthesis gained significant empirical support between 2007 and 2014. Experiments utilizing two-dimensional electronic spectroscopy (2DES) at cryogenic temperatures (77 Kelvin) revealed long-lasting oscillatory signals - termed "quantum beats" - in the FMO complex and PSII 12151617. These non-linear spectroscopic signals were initially interpreted as direct evidence of exceptionally long-lived electronic quantum coherence lasting up to 1.5 picoseconds 121518.
Under this interpretation, the exciton is quantum-mechanically delocalized over multiple chromophores simultaneously. The long-lived phase relation between the clusters was thought to render the process of energy transfer less sensitive to the intrinsic structural disorder of the pigment-protein complexes, allowing the system to reach the reaction center without losing energy to the environment 1216. This sparked a paradigm shift, suggesting that biological macromolecular systems could overcome thermal decoherence to maintain purely electronic superpositions.
Rapid Electronic Decoherence
Subsequent experimental and theoretical work published between 2017 and 2025 has fundamentally revised this interpretation, returning the field to a more constrained view of electronic coherence 9121518. Rigorous re-evaluation of 2DES signatures at ambient, physiological temperatures demonstrated that pure electronic decoherence in the FMO complex and other light-harvesting units actually occurs within roughly 60 to 200 femtoseconds 9121518.
This extremely rapid decoherence aligns perfectly with standard physical predictions for open quantum systems embedded in a warm, polar solvent environment 1215. Because the timescale of electronic decoherence (sub-100 fs) is significantly shorter than the overall timescale of energy transfer under physiological conditions (which requires several picoseconds), sustained long-range electronic superposition cannot be the primary evolutionary design principle driving transport efficiency 912. Theoretical calculations confirm that the efficiency of energy and charge transport does not depend significantly on the persistence of pure electronic coherences, and nature instead favors a thermally driven, dissipative mechanism 912.
Vibronic Coupling and Charge Separation
While pure electronic coherence is too fragile to sustain long-range energy transport, the long-lived quantum beats observed in spectroscopic data were not experimental artifacts. Instead of arising from electronic superposition, these coherent oscillations have been conclusively identified as vibrational and vibronic coherences 1618192021.
Vibronic coupling occurs when the electronic transitions of the pigment molecules interact strongly with specific, discrete vibrational modes (nuclear motions) of either the pigments themselves or the surrounding protein scaffold 192122. When the energy of a localized vibrational mode closely matches the energy gap between two excitonic states, the electronic and vibrational degrees of freedom mix to form a vibronic exciton 162023. This mechanism does not require coherent laser excitation and is valid under natural, incoherent sunlight excitation 16.
This resonant electronic-vibrational configuration sustains and regenerates coherence between electronic states, effectively bridging energy gaps that would otherwise impede efficient transfer 162223. In the PSII reaction center, strong excitonic interaction within specific radical pairs (such as the primary donor complex $P_{D1}$ and $P_{D2}$) enables vibronic coherence to persist for approximately 600 to 2000 femtoseconds even at room temperature 1820.
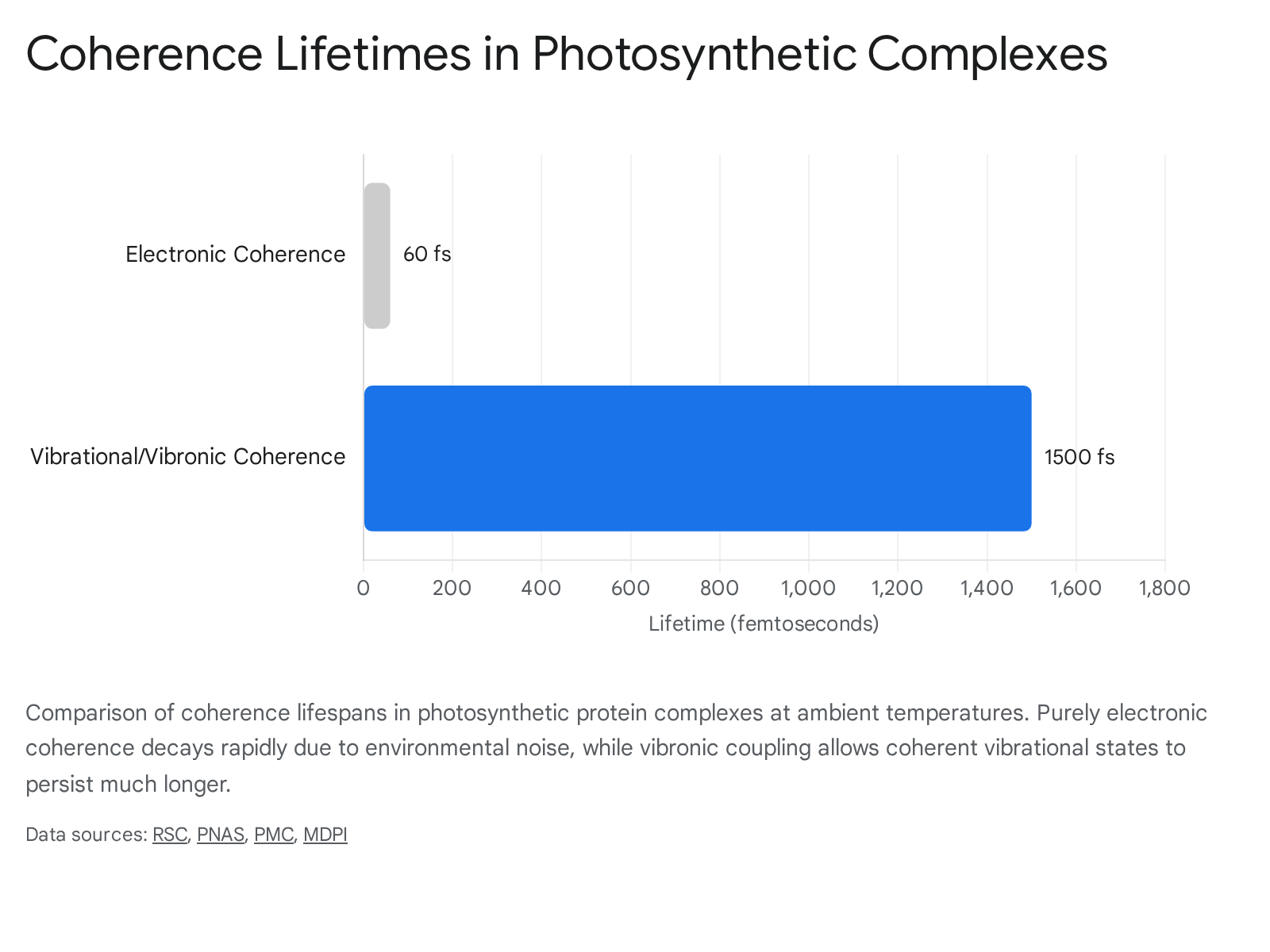
This vibrationally-assisted transport directs energy rapidly and unidirectionally toward the reaction center 91924. By utilizing low-frequency coherent nuclear motions to sample multiple spatial pathways, the system actively drives ultrafast and efficient charge separation before the excitation energy is lost to background noise 1618.
The current scientific consensus maintains that quantum mechanics plays a vital, non-trivial role in photosynthesis, but through the highly optimized mechanism of vibronic coupling and short-lived charge transfer resonances, rather than through global, sustained electronic superpositions 91722. This interplay between electronic and vibrational coherence prolongs the lifetime of vibronic states, enhancing the speed of charge separation 20.
Avian Magnetoreception and Spin Dynamics
While the role of quantum coherence in photosynthesis has been mathematically constrained by thermal decoherence parameters, avian magnetoreception remains the most robust candidate for macroscopic, non-trivial quantum biology 22825. Behavioral studies have long established that migratory birds, such as the European robin (Erithacus rubecula), Eurasian blackcap (Sylvia atricapilla), and garden warbler (Sylvia borin), navigate using a biological compass sensitive to the Earth's magnetic field 302627.
The Earth's magnetic field is exceptionally weak - approximately 50 microteslas - making it theoretically difficult for standard chemical receptors to detect it against the background of thermal noise at physiological temperatures 2829. The classical "magnetite hypothesis," which posits that biogenic iron oxide particles align with the magnetic field to exert mechanical force on ion channels, possesses theoretical viability but has suffered from a persistent inability to identify the actual receptor cells responsible for the compass sense in any animal 2527. Consequently, the prevailing biophysical explanation for this capability is the Radical Pair Mechanism (RPM), a photochemical process intrinsically dependent on the quantum spin states of unpaired electrons 303031.
The Radical Pair Mechanism in Cryptochromes
The RPM is localized in light-sensitive proteins called cryptochromes (Cry), situated within the double-cone photoreceptor cells of the avian retina 25303732. Specifically, Cryptochrome 4a (CRY4a) has been identified as the primary magnetoreceptor candidate. The operational pathway of this quantum sensor involves a complex sequence of photochemical and physical events beginning with photon absorption and culminating in altered chemical yields.
The process is initiated when blue-green light (approximately 370 - 565 nm) enters the avian eye and strikes the CRY4a protein, exciting a non-covalently bound flavin adenine dinucleotide (FAD) chromophore 303133. This photoexcitation triggers a rapid, sequential transfer of an electron along a highly conserved structural chain of four tryptophan residues (the Trp-tetrad) toward the FAD molecule 303133.
This electron transfer event generates a transient radical pair - specifically, the $[FAD^{\bullet -} TrpH^{\bullet +}]$ complex. This complex consists of two molecules separated by a specific physical distance, each possessing an unpaired electron 2530. Because these two unpaired electrons originated from the same molecular orbital before the transfer, their quantum spins are initially entangled in a correlated singlet state, meaning their spin vectors are strictly antiparallel 2530.
The functional essence of the biological compass lies in the coherent interconversion between this singlet state and a spin-triplet state (where the electron spins are parallel). This oscillation is driven by internal magnetic hyperfine interactions between the electron spins and the magnetic nuclei of the surrounding atoms within the protein 253033. Crucially, the rate and amplitude of this singlet-triplet mixing are highly sensitive to the alignment of any external magnetic field, including the weak geomagnetic field of the Earth 3033.
Because the singlet and triplet radical pairs recombine into distinctly different biochemical signaling products, the inclination of the external magnetic field alters the overall chemical yield of the reaction 25. The bird's visual and neurological systems are hypothesized to interpret these varying chemical concentrations as a directional signal, effectively "seeing" the magnetic field lines 293034.
Spectroscopic and Experimental Evidence
Recent international collaborations, including extensive studies in 2025 and 2026 comparing CRY4a in the night-migratory European robin to that of the Japanese quail (Coturnix japonica), have provided robust in vitro evidence for this mechanism. The Japanese quail, a domesticated bird with wild migratory origins, has proven to be an excellent experimental model with magnetic properties mirroring those of the European robin 283033.
Researchers utilized broadband cavity-enhanced absorption spectroscopy (BBCEAS) and transient absorption spectroscopy to track the kinetics of radical formation in purified quail and robin CRY4a. These experiments confirmed definitive magnetic field effects on the radical pairs, demonstrating changes on the order of 10% when a magnetic field is applied 3033. Furthermore, pulsed electron paramagnetic resonance (EPR) validated that the radicals are indeed formed in a spin-correlated singlet state and that the radical separation distances (approximately 2.09 nanometers for the wild-type quail CRY4a) perfectly match the theoretical requirements for a functional quantum compass 30. Confocal fluorescence microscopy further demonstrated that the steady-state fluorescence intensity of the FAD chromophore rises when an external magnetic field is applied, proving that the magnetic field actively alters the probability of forming long-lived radicals 3033.
Evolutionary Optimization and Behavioral Disruption
Biological evolution appears to have intricately optimized this quantum hardware for specific navigational tasks. Genomic and phylogenetic studies show that CRY4a in night-migratory songbirds exhibits significantly higher magnetic sensitivity and specific mRNA upregulation during migratory seasons compared to the equivalent protein in non-migratory species, such as chickens 302937.
Moreover, alternative splice variants within the same organism lack this functionality. For instance, European robin Cryptochrome 4b (ErCry4b) has been shown through computational simulations and mass spectrometry to lack the ability to bind the FAD cofactor entirely. Without the FAD cofactor, electron transfer cannot occur, rendering ErCry4b photochemically inactive and irrelevant to the radical pair mechanism 3135. This specific functional isolation of ErCry4a underscores its specialized evolutionary role.
| Cryptochrome Variant / Species | Migratory Status | FAD Binding | Magnetic Sensitivity (In Vitro) | Proposed Role in Magnetoreception |
|---|---|---|---|---|
| ErCry4a (European Robin) | Migratory | Yes | High | Primary magnetosensor candidate 3031 |
| CjCry4a (Japanese Quail) | Migratory Origins | Yes | High | Primary magnetosensor candidate 3033 |
| GgCry4a (Chicken) | Non-Migratory | Yes | Low / Moderate | Baseline circadian/photoreceptor function 302937 |
| ErCry4b (European Robin) | Migratory | No | None | Irrelevant to radical pair mechanism 3135 |
Behavioral experiments further corroborate the quantum hypothesis in vivo. When migratory birds are placed in testing funnels and exposed to exceedingly weak, broadband radio-frequency (RF) electromagnetic fields in the megahertz range, their navigational abilities are entirely disrupted 2930. Classical magnetic particles, such as magnetite, are entirely unaffected by such low-intensity RF fields. However, these specific frequencies precisely match the energetic resonance required to scramble the delicate singlet-triplet spin dynamics of entangled electrons 29. This phenomenon, often referred to as the "quantum needle" effect, is widely considered the most compelling behavioral proof that birds rely on quantum resonance to navigate, directly linking molecular quantum physics to macroscopic animal behavior 29.
Olfaction and the Vibration Theory Debate
The sense of smell relies on the physical interaction between airborne volatile molecules (odorants) and specialized G-protein coupled receptors (GPCRs) located in the olfactory epithelium within the nasal cavity 3637. The classical and widely accepted model for this interaction is the docking theory, historically termed the shape theory 3638. This model posits that an odorant is recognized by a specific receptor through a complementary structural fit, governed by a complex constellation of weak non-covalent interactions. These interactions include van der Waals forces, hydrogen bonding, dipole attraction, pi-stacking, metal ion interactions, and hydrophobic effects 3638.
Inelastic Electron Tunneling in Odorant Perception
An alternative hypothesis, the Vibration Theory of Olfaction (VTO), originally proposed by Malcolm Dyson in 1928 and substantially refined by Robert H. Wright in 1954 and Luca Turin in 1996, posits a fundamentally different, quantum mechanical mechanism for smell 3639. The VTO suggests that olfactory receptors do not merely recognize the spatial geometry and charge distribution of a molecule, but actively detect its molecular vibrational frequencies in the infrared range via inelastic electron tunneling spectroscopy (IETS) 363740.
According to the Turin model, the odorant molecule must first fit into the receptor's binding site. Once docked, the odorant acts as a bridge between an electron donor site and an electron acceptor site within the protein. If the vibrational energy of the odorant's specific chemical bonds matches the energy gap between the donor and acceptor, an electron can quantum mechanically tunnel across the molecule 3640. This inelastic tunneling event supposedly triggers the conformational change in the GPCR necessary for the signal transduction pathway 36.
Proponents of VTO argue that a pure shape theory struggles to explain why molecules with vastly different geometric shapes can possess nearly identical odors - such as the structurally diverse range of synthetic musk compounds - or why similarly shaped molecules can smell differently 36. Furthermore, they highlight the perceived failure of the shape theory to predict the odor of novel synthesized molecules reliably, a major hurdle for the fragrance industry 41.
Isotopomer Testing and Experimental Refutations
To definitively test the VTO, researchers rely on isotopomers - molecules in which specific atoms are replaced with their heavier isotopic counterparts, most commonly substituting hydrogen with deuterium 4041. Because deuterium has twice the atomic mass of hydrogen, carbon-deuterium (C-D) bonds vibrate at significantly lower frequencies than carbon-hydrogen (C-H) bonds. Crucially, this isotopic substitution does not meaningfully alter the molecule's physical shape, atomic radius, or electrostatic surface properties 363841.
If the quantum vibration theory is functionally correct, an organism or an isolated receptor should be able to differentiate between a hydrogenated odorant and its fully deuterated isotopomer, as their vibrational spectra are entirely distinct. Initial psychophysical tests in the 1990s and early 2000s yielded mixed but highly publicized results, with some studies claiming that human subjects could easily smell the difference between deuterated and undeuterated musk molecules, such as cyclopentadecanone 4041. Similarly, studies on Drosophila suggested behavioral differentiation based on isotopomeric composition 3637.
However, rigorous, multidisciplinary research published between 2015 and 2024 has systematically dismantled these claims. A major collaborative effort shifted the debate from subjective human psychophysics to rigorous in vitro biophysics by testing heterologously expressed olfactory receptors directly 4041. When specific, known olfactory receptors - such as the human musk-recognizing receptor OR5AN1 and the mouse thiol-recognizing receptor MOR244-3 - were challenged with purified isotopomers, they exhibited entirely identical dose-response activation profiles to both hydrogenated and deuterated versions of their cognate ligands 384041. The receptors completely failed to distinguish between the vibrational differences. Furthermore, contrary to VTO predictions that specific infrared bands (e.g., the 1,380- to 1,550-cm - 1 range) are essential for musk odor, tests revealed that muscone-d30 lacks these bands yet still robustly triggers the human OR5AN1 musk receptor 40.
Perireceptor Events and Trace Impurities
The failure of the VTO at the receptor level necessitated a re-evaluation of the earlier behavioral studies that seemingly supported it. Subsequent gas chromatography and nuclear magnetic resonance (NMR) analyses revealed that many of the deuterated synthetic samples used in prior experiments contained trace impurities not present in the hydrogenated control samples 4042. These trace impurities were sufficient to activate different receptor profiles, causing a perceived difference in smell that was erroneously attributed to vibrational frequencies 4042.
Additionally, differences observed in live animal testing can be fully explained by classical perireceptor events. Isotopomers are known to diffuse through the nasal mucus at slightly different rates and can be metabolized differently by biotransformation enzymes in the mucosal layer prior to reaching the receptor epithelium 40. These secondary isotope effects alter the concentration of the odorant reaching the receptor, generating a differential neural response entirely independent of quantum electron tunneling 4042.
Finally, theoretical biophysics models have demonstrated that the proposed inelastic electron tunneling mechanism is highly implausible within the standard environment of a GPCR. The necessary signal from the odorant's subtle molecular vibrations would be easily masked or suppressed by the overwhelming thermal noise and the massive quantum vibrational modes of the surrounding protein environment (such as peptide bonds and water molecules) 3640.
Consequently, the current scientific consensus unequivocally rejects the Vibration Theory of Olfaction. There is no reliable experimental evidence at the receptor level to suggest that electron tunneling or vibrational frequency detection triggers olfactory responses 404142. The classical, shape-based docking theory - incorporating the full suite of electrostatic and structural interactions - remains the sole predictive framework for olfaction, rendering quantum effects in smell perception trivial 3637.
Synthesis
The rigorous investigation into quantum biology over the past two decades reveals a highly nuanced biophysical landscape where the boundary between classical thermodynamics and functional quantum mechanics is tightly dictated by the precise timescales of environmental decoherence and structural optimization.
In the realm of photosynthesis, the initial hypothesis that natural light-harvesting complexes utilize long-lived electronic quantum superposition to perfectly route energy has been conclusively revised. The sub-100 femtosecond collapse of pure electronic coherence in physiological temperatures precludes such a mechanism. Instead, the consensus indicates that nature optimizes energy transfer through vibronic coupling, exploiting resonances between electronic transitions and the structured vibrations of the protein scaffold. This represents a highly sophisticated evolutionary tuning of molecular dynamics, functioning as a dissipative, vibration-assisted process rather than a pure electronic quantum computer.
Conversely, avian magnetoreception stands as the most robust demonstration of non-trivial quantum biology to date. The evidence supporting the Radical Pair Mechanism in retinal cryptochromes is substantial. The intrinsic sensitivity of spin-correlated singlet and triplet states to ultra-weak magnetic fields, combined with in vitro verification of radical distances and the behavioral disruption caused by resonance-matching radio frequencies, provides compelling evidence that migratory birds navigate by interpreting functional quantum spin dynamics.
Finally, the mechanisms governing olfaction highlight the boundaries of the quantum biological paradigm. Despite the theoretical elegance of the Vibration Theory of Olfaction, exhaustive empirical evidence rules out quantum inelastic electron tunneling as a functional mechanism for odor recognition. The absolute inability of isolated odorant receptors to distinguish between identical molecular shapes with different vibrational signatures firmly re-establishes the classical docking theory.
Ultimately, biological systems do not broadly circumvent the laws of quantum physics. Instead, through billions of years of selective pressure, specific macromolecular complexes have been structurally optimized to exploit transient quantum phenomena - such as vibronic coupling and spin entanglement - precisely within the brief temporal windows before environmental decoherence prevails.