Predictive Processing in Neuroscience and Psychiatry
The conceptualization of brain function has undergone a profound paradigm shift over the past two decades. Departing from the classical view of the nervous system as a passive, bottom-up processor of environmental stimuli, contemporary cognitive neuroscience increasingly treats the brain as a proactive, predictive engine. This framework, known as predictive processing, posits that the nervous system continuously generates top-down expectations regarding the sensory environment and relies on bottom-up data primarily to compute prediction errors. These errors represent the discrepancies between what the brain expects and what the sensory organs actually detect. By continuously updating internal generative models to minimize these errors, the brain efficiently navigates uncertainty, optimizes metabolic resource allocation, and supports highly adaptive behavior across diverse contexts.
As this theoretical scaffolding has matured, its application has expanded far beyond the initial boundaries of basic sensory perception. Predictive processing now serves as an organizing principle for research into motor control, interoception, language comprehension, and complex social cognition. Concurrently, it is fundamentally reshaping the field of computational psychiatry. By conceptualizing psychiatric conditions not merely as descriptive symptom clusters or chemical imbalances, but as specific computational dysfunctions in the brain's inferential machinery, researchers are actively mapping complex syndromes onto formal neurobiological and mathematical parameters. However, the exact neural implementation of these algorithms - alongside the broader metaphysical claims surrounding them - remains the subject of intense empirical investigation and theoretical debate.
Theoretical Foundations of the Predictive Brain
At the absolute core of predictive processing is the assertion that the brain implements a sophisticated form of Bayesian inference. Because biological organisms cannot directly access the external physical world, they are restricted to the sensory perturbations registered by their peripheral nervous systems. To navigate this restricted reality, the brain must construct a hierarchical generative model of the latent, hidden causes of its sensory input, continuously guessing what objects or events in the world are responsible for the signals it receives 123.
The Generative Model and Precision-Weighted Prediction Error
Within a formal Bayesian framework, perception is defined as the optimal integration of prior beliefs (predictions) and sensory likelihoods (incoming evidence). The resulting posterior probability represents the brain's best estimation of the state of the world at any given moment. Predictive processing translates this mathematical principle into neural architecture by suggesting that higher-order cortical areas pass probabilistic predictions down the hierarchy. When incoming sensory data align perfectly with these top-down predictions, the data are effectively "explained away," and their further feedforward transmission is suppressed. When a mismatch occurs, a prediction error is generated. This error signal is then passed up the hierarchy to update the prior model, refining the accuracy of future predictions 3456.
A crucial computational variable in this dynamic is precision weighting. Not all sensory inputs or prediction errors are equally informative. In highly volatile or noisy environments - such as attempting to view a scene through heavy fog or listening to speech in a crowded room - sensory data is inherently unreliable. The brain must therefore dynamically estimate the precision, defined mathematically as inverse variance, of its prediction errors. Errors that are deemed highly precise strongly drive model updating, whereas low-precision errors are largely ignored or attenuated, allowing the system to rely more heavily on its stable, top-down priors 27. This precision-weighting mechanism allows for flexible adaptation to changing environmental contexts.
Active Inference and Environmental Perturbation
The extension of classical predictive processing into the domains of motor control and behavioral ecology is articulated through the concept of active inference. While classical perceptual inference minimizes prediction error by updating the internal neural model to match the external world, active inference achieves error minimization by updating the external world to match the internal model. Organisms engage in actions to fulfill their proprioceptive and interoceptive predictions, thereby ensuring that they remain within expected physiological boundaries 789.
For instance, rather than computing a complex motor command to grasp an object, the brain simply predicts that the hand is grasping the object. The resulting proprioceptive prediction error (because the hand is not currently grasping the object) is minimized not by changing the belief, but by initiating classical spinal reflex arcs that move the hand to fulfill the prediction. This elegant formulation unifies perception and action under a single computational imperative: the minimization of prediction error 710.
The Free Energy Principle and System Topology
These inferential principles have been subsumed under a broader, highly ambitious theoretical framework known as the Free Energy Principle (FEP). The FEP proposes that any self-organizing biological system that successfully resists the natural thermodynamic tendency toward entropy must, in effect, minimize its variational free energy 7111213. In information theory, variational free energy constitutes an upper bound on surprise, which is defined as the negative log probability of encountering specific sensory states. By minimizing free energy over time, an organism ensures that it avoids surprising states and remains within the narrow, highly probable range of physiological conditions compatible with its survival and homeostasis 1112.
The FEP formalizes the boundary between an organism and its environment using the concept of a Markov blanket. A Markov blanket provides statistical conditional independence, separating the internal states of a system from the external latent causes via an intermediate layer composed of sensory and active states 101214.
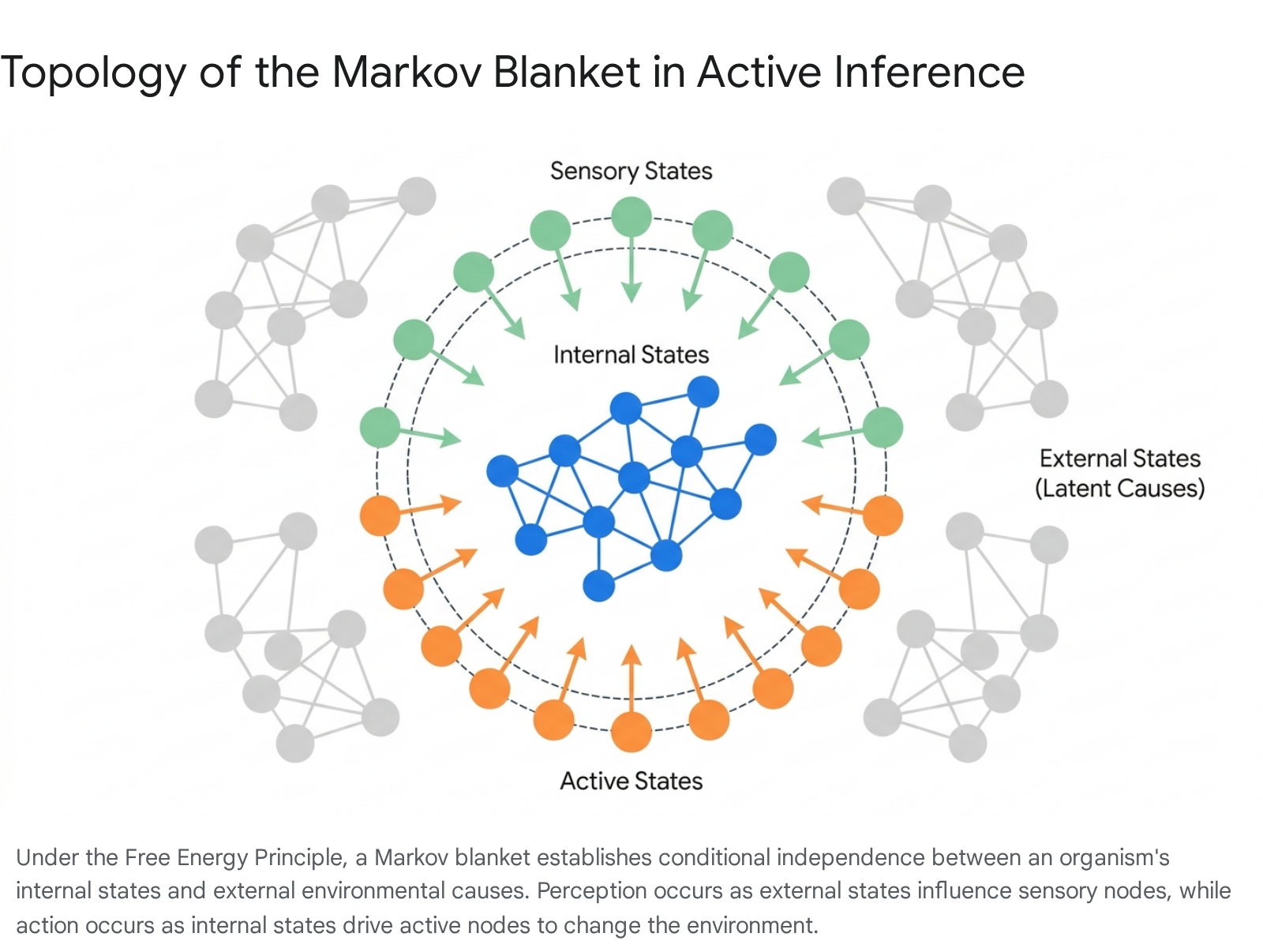
Under this specific mathematical topology, the internal states of any system possessing a Markov blanket will necessarily appear to engage in active Bayesian inference in order to preserve their structural and functional integrity against environmental fluctuations 121315.
Ontological Controversies and the Ergodicity Debate
While predictive coding and active inference have proven to be highly productive frameworks for designing empirical neuroscience experiments, the overarching Free Energy Principle has sparked intense methodological and philosophical controversy. The debate primarily centers on the transition from formal mathematical descriptions of statistical systems to ontological claims about biological reality.
A primary criticism leveled against the FEP is that it operates more as an unfalsifiable metaphysical axiom than a testable scientific theory. Critics argue that the framework exhibits excessive rhetorical elasticity. Because the FEP asserts that any persisting system minimizes free energy by definition, any observed biological behavior - no matter how contradictory - can be retroactively fit to the model 1618. This presents a fundamental problem of falsifiability; the theory can easily assimilate contradictory experimental outcomes by simply assuming that the organism was operating under a different, unobservable prior expectation or a hidden, long-term utility function 1617. Furthermore, critics argue that the FEP commits a fundamental category error by inadmissibly transferring concepts from non-equilibrium statistical mechanics to evolutionary biology. By asserting that systems act to minimize free energy, the framework risks reintroducing teleology into the natural sciences, describing biological self-organization as a goal-directed optimization process rather than the mechanistic outcome of stochastic physical constraints and natural selection 1618.
The mathematical substructure of the FEP has also faced rigorous technical scrutiny. Original formulations of the FEP rely heavily on the assumption of ergodicity, which posits that the ensemble average of a system's states equals its temporal average over a long, idealized horizon. Critics point out that biological systems, which undergo irreversible developmental trajectories and exist in highly non-stationary environments, flagrantly violate the ergodic assumption, rendering the ensuing mathematical proofs inapplicable to real organisms 71518. Some recent reformulations attempt to bypass this limitation by grounding FEP computations in non-equilibrium (NEQ) densities, linking Bayesian inference to classical paths of least action in stochastic dynamical systems 713.
Additionally, scholars increasingly distinguish between "Pearl blankets," which serve as purely formal, mathematical representations of conditional independence in Bayesian networks, and "Friston blankets," which involve the ontological assertion that these statistical boundaries map directly onto the physical boundaries of living organisms, such as cell membranes or skin 18. Applying the Markov blanket formalism to infer realistic biological properties is frequently characterized by instrumentalists as committing the literalist fallacy, effectively confusing a useful mathematical map with the physical territory it describes 18.
Neural Implementation and Circuit Architecture
Translating the highly abstract algorithms of predictive processing into specific neurobiological architectures is a primary objective of modern systems neuroscience. The transition from theoretical elegance to empirical tractability requires identifying exactly how predictions and errors are encoded in cortical tissue. For over a decade, classical hierarchical predictive coding models dominated the field, but recent high-density laminar recordings have catalyzed a substantial shift toward an alternative mechanistic model known as predictive routing.
Classical Hierarchical Predictive Coding
The classical predictive coding framework, originally outlined mathematically by Rao and Ballard and later expanded by Friston, posits a dedicated microcircuitry within the standard cortical column 41920. This model mandates the existence of highly specialized populations of neurons: state units that encode the brain's generative model, and distinct error units that exclusively calculate the subtractive discrepancy between top-down predictions and bottom-up sensory inputs 4519.
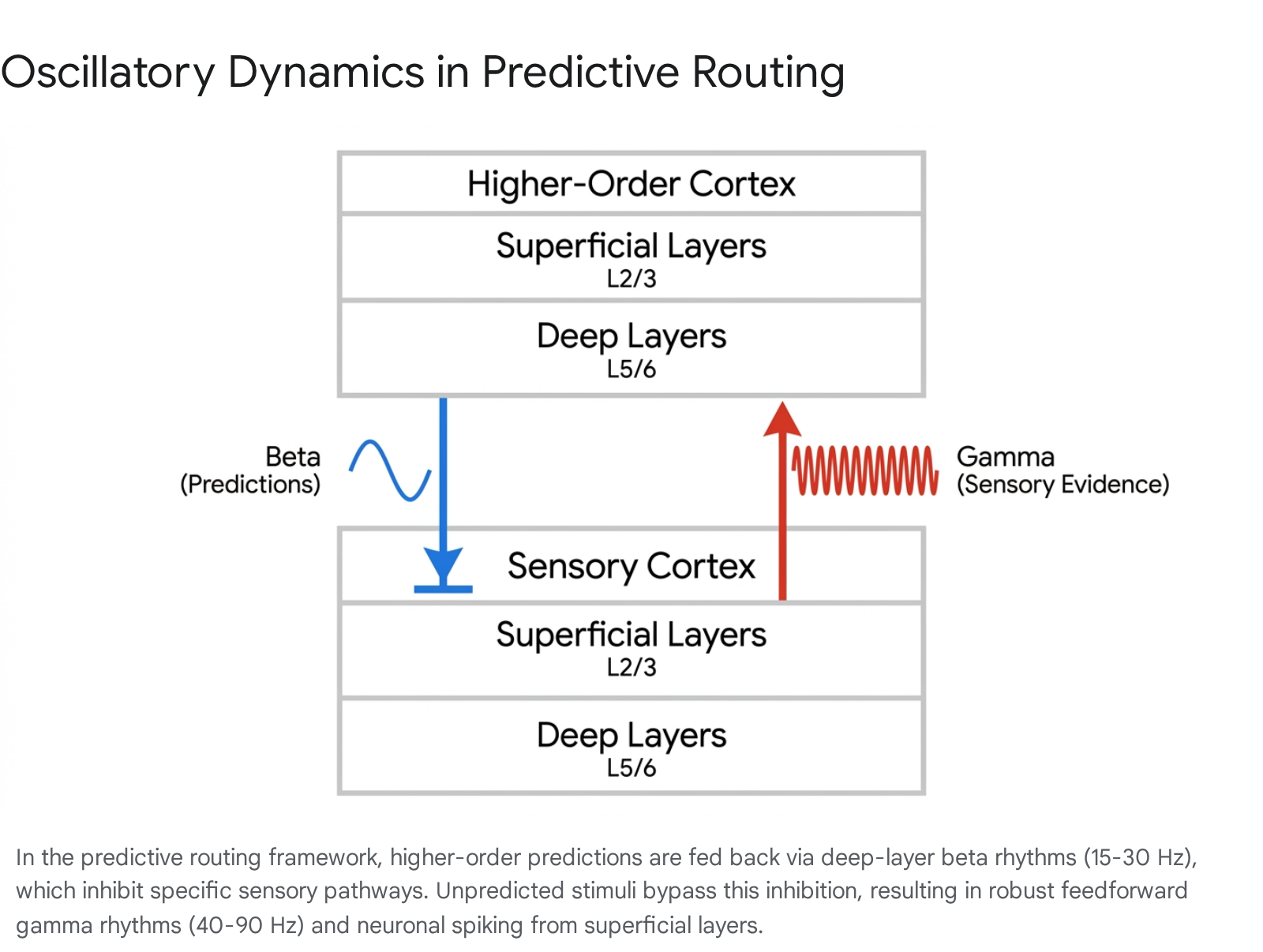
According to this classical model, sensory inputs arrive at lower cortical areas, while predictions are sent via feedback connections originating from the deep layers (Layers 5 and 6) of higher-order areas. The explicit error units, hypothesized to reside primarily in the superficial layers (Layers 2 and 3), output the residual mismatch via feedforward pathways up the cortical hierarchy 31921. Classical predictive coding thereby suggests a continuous, bidirectional exchange of specialized signals across adjacent brain areas, explicitly requiring the physical computation and transmission of prediction error as a distinct neural currency.
Algorithmic Comparisons in Cortical Networks
To empirically test how information flows through the cortex, researchers have deployed high-density local field potential (LFP) recordings across triplets of hierarchical brain areas. For example, recent visual search task studies in non-human primates have analyzed simultaneous recordings from visual area V4 (lower), parietal area 7A (middle), and the prefrontal cortex (PFC, higher) to compare competing algorithmic models of neural computation 119.
The evaluation of neural data across these hierarchies has primarily contrasted three models: classical Predictive Coding, standard Autoencoders, and Predictive Routing.
| Algorithmic Model | Mechanism for Sensory Processing | Feedback Propagation | Requirement for Explicit Error Units | Empirical Fit with LFP Dynamics |
|---|---|---|---|---|
| Predictive Coding (PC) | Subtractive comparison between sensory input and top-down prediction. | High. Extensive top-down signaling across hierarchical levels. | Yes. Mandates dedicated superficial layer units for computing residuals. | Explains deep-layer activity well, but lacks distinct cellular evidence for dedicated error populations. 419 |
| Autoencoders (AE) | Feedforward propagation of state representations without feedback modulation. | None. State signals are propagated strictly feedforward. | No. Sensory states are compressed and passed upward directly. | Poor fit for cortical dynamics, as it ignores known anatomical feedback pathways critical for consciousness. 119 |
| Predictive Routing (PR) | Top-down rhythmic preparation that selectively suppresses predicted pathways. | High. Contextual predictions prepare sensory channels. | No. Prediction errors are simply the natural output of unsuppressed sensory neurons. | Best fit for superficial layer dynamics and push-pull oscillatory relationships between beta and gamma rhythms. 131920 |
The Transition to Predictive Routing
While neuroimaging studies utilizing functional magnetic resonance imaging (fMRI) in oddball paradigms frequently show macroscopic activation patterns consistent with classical predictive coding, direct cellular and electrophysiological evidence for specialized, dedicated error units has remained persistently elusive 3420. Recent large-scale neurophysiological studies have proposed the revised predictive routing model to account for these findings 13192022.
Predictive routing argues vehemently against the existence of specialized error-computing circuits. Instead, it suggests that the brain utilizes its standard sensory processing architecture but dynamically gates information flow through phase-specific rhythmic modulation 192023. In this framework, predictions are not explicitly compared against sensory inputs in a subtractive manner. Rather, top-down predictions act by proactively preparing the cortex. Higher-order areas send rhythmic signals that selectively inhibit the specific neural pathways in the lower sensory cortex that are tuned to process the expected input 320.
If a highly predicted stimulus occurs, it arrives at a computationally inhibited pathway, resulting in attenuated neural firing and reduced feedforward transmission. Conversely, if an unpredicted stimulus occurs, it arrives at an excitable, unprepared pathway, triggering a robust feedforward sensory response 202624. Therefore, the prediction error is not a distinct calculation generated by specialized error neurons; it is simply the natural, unaltered consequence of an unsuppressed sensory signal feeding forward into higher cortical areas 1319.
Laminar Specificity and Oscillatory Dynamics
The predictive routing model is heavily grounded in the distinct oscillatory dynamics of the cortex, mapping specific brain rhythms and cellular layers to distinct predictive functions. High-density recordings demonstrate a clear, mathematically definable push-pull dynamic between high-frequency and low-frequency bands across the cortical layers.
| Neural Rhythm | Frequency Band | Cortical Origin | Computational Role in Predictive Routing | Neurochemical Dependency |
|---|---|---|---|---|
| Gamma | 40 - 90 Hz | Superficial Layers (L2/3) | Encodes bottom-up sensory evidence; feeds forward unpredicted information. | NMDA-dependent excitatory transmission. 342024 |
| Beta / Alpha | 8 - 30 Hz | Deep Layers (L5/6) | Transmits top-down contextual predictions; selectively inhibits feedforward gamma. | GABA-mediated inhibition, Dopaminergic modulation. 42024 |
| Theta | 2 - 6 Hz | Distributed | Encodes slower, longer-scale temporal prediction errors and environmental volatility. | Complex network interactions. 2025 |
During periods of high environmental predictability, top-down beta power significantly increases, successfully suppressing superficial gamma oscillations and their associated neural spiking. When expectations are violently violated - such as in a global oddball paradigm where a repetitive sequence is unexpectedly broken - the lack of top-down beta-mediated inhibition allows for a rapid surge in superficial gamma power and spiking, which efficiently propagates the novel information up the cortical hierarchy 342226.
Causal support for this specific rhythmic architecture comes from neuropharmacological studies utilizing propofol-induced anesthesia. Propofol profoundly decreases high-amplitude alpha and beta oscillations in the posterior cortex while enhancing global GABAergic tone. Research demonstrates that when higher-order beta feedback is knocked out by propofol, the predictive suppression of lower sensory areas is eliminated. This leads to a paradoxical disinhibition of the sensory cortex, where oddball responses in auditory areas are no longer modulated by predictability, effectively breaking the brain's predictive routing mechanism 222426.
To further test the plausibility of these oscillatory interactions, computational neuroscientists have developed biophysically realistic neural circuit models trained using self-supervised algorithms, such as the generalized Stochastic Delta Rule (gSDR). Without requiring manual parameter tuning, circuits trained with gSDR naturally develop the gamma-beta push-pull dynamics observed in biological tissue, demonstrating that these specific rhythmic interactions are the optimal network solution for handling predictive routing objectives 2226.
Large-Scale Brain Networks and the Dynamic Prediction Network
While much of the foundational research in predictive processing focuses on localized, microscopic cortical column interactions, there is a growing consensus that predictive capabilities depend on domain-general, macroscopic brain networks.
Recent high-powered Activation Likelihood Estimation (ALE) meta-analyses evaluating hundreds of neuroimaging experiments have identified a domain-general architecture termed the Dynamic Prediction Network. Regardless of whether the task involves cognitive control, auditory attention, motor execution, language processing, or social cognition, instances of prediction incongruency reliably activate a specific set of highly connected brain hubs 27. This distributed network encompasses the bilateral insula, frontal gyri, the claustrum, parietal lobules, and the temporal gyri. The consistent recruitment of these regions suggests that predictive processing is not a series of isolated, domain-specific modules, but rather a unified, integrated computational strategy utilized across the entire cerebrum 27.
To further untangle how predictions generalize across different sensory domains and task demands, the field is increasingly relying on massive, globally distributed collaborative research efforts. Projects such as the Allen Institute's OpenScope initiative are pioneering crowd-sourced neuroscience studies. By pooling experimental designs from dozens of international laboratories, researchers are using standardized, high-throughput recording platforms to probe the specific trade-offs the brain makes when shifting from predicting continuous sequences to anticipating isolated discrete events 2829. Concurrently, significant multinational funding from institutions like the European Research Council (ERC) and the Dutch Research Council (NWO) is driving long-term investigations into how predictive processing varies among healthy individuals, specifically analyzing the neural mechanisms involved in the formation, deployment, and generalization of sensory predictions across rapidly shifting contexts 33.
Computational Psychiatry and Inferential Failure Modes
The precision with which predictive processing defines optimal neural inference provides an incredibly robust theoretical framework for identifying what happens when that inference fails. The emerging discipline of computational psychiatry actively leverages this framework to move beyond traditional, descriptive symptom-based classifications (nosology) toward identifying the mechanistic breakdowns underlying severe mental illness. Within this paradigm, psychiatric disorders are increasingly understood as specific, quantifiable disruptions in the delicate balance of predictions, prediction errors, and their precision weightings.
Conceptualizing Psychopathology as Computational Dysfunction
Rather than viewing psychiatric conditions as categorically distinct and isolated entities, the predictive processing framework identifies transdiagnostic computational "failure modes" that can manifest across multiple, seemingly disparate clinical presentations 9303536. These failure modes almost universally involve structural imbalances in how the cortical hierarchy evaluates information. This typically presents as aberrant precision weighting, where the brain fails to appropriately assign statistical confidence to either its prior beliefs or incoming sensory data 231. This leads to two primary pathological states: the generation of over-precise priors that override contradictory sensory evidence (leading to hallucinations or inflexible behavior), or the reliance on hyper-precise errors, where the failure to suppress baseline noise causes the system to treat irrelevant sensory fluctuations as highly salient, forcing constant, exhausting model updates 2731.
| Clinical Condition | Primary Computational Failure Mode | Hierarchical Disruption | Behavioral / Clinical Manifestation | Theoretical Tensions & Uncertainties |
|---|---|---|---|---|
| Schizophrenia (Early/Prodromal) | Aberrant precision weighting; hyper-salient sensory errors. | Sensory to Associative | Overwhelm, lack of contextual integration, blunted MMN. | How does the transition from chaotic weak priors to rigid delusions occur dynamically? 3631 |
| Schizophrenia (Established) | Over-precise, rigid top-down priors overriding sensory data. | Executive to Associative | Paranoia, fixed delusions, internally generated hallucinations. | Disentangling state vs. trait alterations in predictive beta-band signaling. 2632 |
| Autism Spectrum Disorder (ASD) | Hyper-precise prediction errors; heavy reliance on sensory updating. | Sensory to Associative | Sensory overload, repetitive behaviors, delayed adaptation to volatility. | Debate remains regarding whether priors are fundamentally weak or simply outweighed by precise errors. 36313334 |
| Major Depressive Disorder (MDD) | Over-precise negative priors; failure to assimilate positive errors. | Executive to Affective/Somatic | Anhedonia, learned helplessness, model-free behavioral dominance. | Differentiating MDD phenotypes based on specific interoceptive vs. exteroceptive biases. 6835 |
| Chronic Pain / Somatic Syndromes | Hyper-precise somatic priors coupled with avoidance behavior. | Associative to Interoceptive | Amplified pain perception absent tissue damage, fear-avoidance cycles. | Determining if interoceptive deficits act as a primary cause or secondary consequence of chronic pain. 36434445 |
Schizophrenia and the Psychosis Spectrum
Schizophrenia has served as the paradigmatic test case for predictive processing accounts of psychopathology. Bizarre symptoms such as hallucinations, paranoia, and delusions are highly amenable to rigorous Bayesian explanations. Furthermore, the disorder's complex etiology - which involves profound disruptions to dopaminergic and glutamatergic neurotransmission, as well as distinct genetic markers (such as AMBRA1 and HHAT variants) and HPA-axis dysregulation - maps cleanly onto the neurochemical mechanisms believed to govern precision weighting and cortical gain control 374738.
In the early, prodromal stages of psychosis, the primary failure mode is hypothesized to be the aberrant allocation of precision to low-level prediction errors. The brain fails to suppress standard environmental sensory noise, resulting in an overwhelming influx of hyper-salient, unpredicted signals 2732. Behaviorally, this manifests as a weak central tendency and a profound inability to rely on context 732. Electrophysiologically, this phenomenon is evidenced by significant reductions in Mismatch Negativity (MMN) - an event-related potential that normally signals the brain's automatic detection of environmental deviance. In schizophrenia, because the baseline neural state is already flooded with aberrant prediction errors, the brain's specific, localized response to true novelty (the MMN) is heavily blunted 639. Magnetoencephalography (MEG) studies evaluating masked-face paradigms further corroborate this, showing that individuals with schizophrenia struggle to integrate newly available sensory evidence with internally inferred representations 39.
To cope with this persistent, highly distressing state of constant prediction error, the higher-order associative cortex eventually attempts to impose rigid structure. Delusions are thus conceptualized as highly precise, inflexible top-down priors formed in a desperate, maladaptive attempt to "explain away" the chaotic lower-level sensory data 267. Once these over-precise priors are established, they dictate perception so strongly that they can generate internally driven sensory experiences (hallucinations) that are entirely detached from external sensory input 232. This correlates directly with extensive findings that individuals with schizophrenia demonstrate altered beta-band dynamics, reflecting severely compromised mechanisms for top-down contextual maintenance and predictive routing 46.
Autism Spectrum Disorder and Interactive Updating
Autism Spectrum Disorder (ASD) presents a contrasting, and theoretically intensely debated, failure mode within the predictive processing framework. A highly influential early theory proposed that ASD is fundamentally characterized by "weak priors" (the attenuated-prior model). This model suggested that autistic individuals fail to form strong, generalized global expectations, and therefore perceive the world in a fragmented, highly detailed, and literal manner 234.
However, more recent and mathematically rigorous formulations suggest that rather than priors being weak, the primary computational deficit in ASD may be the inflexible, hyper-precision of low-level sensory prediction errors 23134. Because every minor deviation in the sensory environment is treated as a highly precise prediction error, the world feels overwhelmingly chaotic and unpredictably volatile. This directly accounts for core ASD clinical phenotypes: extreme hyper-sensitivity to sensory stimuli, intense distress during unexpected environmental changes, and a heavy reliance on repetitive behaviors (stimming) and strict daily routines. In the context of active inference, these behaviors act as strategies to artificially construct a highly predictable, low-surprise environment to counteract a failing inferential system 3140.
Recent psychophysical studies tracking iterative belief updating in ASD further clarify this mechanism. Using complex duration reproduction tasks modeled with two-state Bayesian parameters, researchers demonstrate that while autistic individuals are fully capable of utilizing prior knowledge, they rely significantly more on immediate sensory inputs to dynamically update their beliefs compared to neurotypical controls. This results in much slower adaptation to environmental volatility, as their inferential machinery remains tethered to immediate sensory evidence rather than abstracting stable, long-term predictive models 3334.
Affective Disorders and Interoceptive Inference
The predictive framework also extends powerfully into the study of affective and somatic disorders, largely via the emerging concept of interoceptive inference - the brain's predictive modeling of its internal visceral, autonomic, and physiological states 4441. Interoceptive signaling relies heavily on vagal tone, and deviations in this circuitry directly impact emotional regulation and self-awareness 35.
In Major Depressive Disorder (MDD) and severe anxiety, individuals frequently exhibit over-precise negative priors. The brain's generative model strictly expects failure, social rejection, or physiological depletion. When these priors are overwhelmingly strong, they systematically suppress positive prediction errors (evidence of success, safety, or reward), rendering the individual entirely unable to update their models in response to positive environmental feedback 7841. In the specific terminology of active inference, a depressed agent struggles to identify actionable paths to higher-valence states within their internal model, resulting in profound psychomotor retardation, reliance on model-free behavioral dominance, and severe anhedonia 68.
Chronic functional pain is similarly viewed through the lens of interoceptive predictive failure 434542. While acute pain serves as a vital prediction error signaling bodily damage, in chronic functional pain syndromes, the brain develops a hyper-precise prior expectation of pain that persists long after tissue healing. If a patient expects a specific movement to hurt, this precise prior actively shapes the actual perception of nociceptive input. Crucially, the expectation and fear of pain lead directly to avoidance behavior. Because the patient avoids the movement, they actively deprive their generative model of the innocuous sensory data (the "safe" movement) required to generate a positive prediction error and update the maladaptive prior. The chronic pain state thus becomes a self-entrenching cycle of failed inference 93643. Interestingly, studies investigating the Error-Related Negativity (ERN) component in pain avoidance found that, contrary to initial hypotheses, individuals with elevated ERN amplitudes initially showed slower learning of avoidance behaviors, highlighting the complex, non-linear relationship between neural error signaling and overt behavioral adaptation 41.
Translational Applications and Artificial Intelligence
The ultimate utility of the predictive processing framework lies in its powerful translational potential. By identifying specific neuro-computational parameters, researchers are rapidly developing precision medicine interventions that directly target the mechanics of inference, rather than merely attempting to manage symptom clusters. Concurrently, the principles of human predictive processing are being mapped onto the architecture of artificial intelligence, yielding critical insights into both fields.
Clinical Interventions, Biomarkers, and Neuromodulation
Traditional psychiatric diagnosis relies almost entirely on subjective behavioral reporting, which suffers from massive clinical heterogeneity and overlap. Predictive processing offers objective, quantifiable computational assays 3035. For example, the Mismatch Negativity (MMN) and specific electroencephalographic (EEG) signatures - such as disruptions in beta-gamma push-pull dynamics - are currently being rigorously evaluated as highly predictive biomarkers for the onset of psychosis in clinical high-risk populations, offering the potential for true preventative psychiatry 639.
Furthermore, advanced brain age models and structural MRI parameters are being integrated with machine learning to predict patient treatment responses. In recent trials investigating repetitive transcranial magnetic stimulation (rTMS) for comorbid depression, machine learning analysis of morphometric features successfully predicted responders by tracking structural markers intimately linked to cortical adaptability and predictive capability 4344. Neuromodulation techniques are also being designed to directly alter predictive processing networks; for instance, accelerated continuous theta burst stimulation (a-cTBS) targeting the primary motor cortex is currently in clinical trials to correct neural oscillatory imbalances in ASD. Recent multicenter trials utilizing a rigorous five-day a-cTBS protocol have shown significant, measurable early promise in improving social communication and overall clinical global impression scores in autistic children 45.
Similarly, targeted digital therapeutics are being designed to intentionally restructure generative models. Innovative mobile interventions for schizophrenia currently utilize specialized cognitive behavioral techniques to deliberately feed patients structured data designed to generate positive prediction errors. This targeted data influx aims to slowly dismantle the rigid, biased priors that underpin defeatist attitudes and paranoid delusions, showing excellent retention and symptom reduction in preliminary trials 56.
Artificial Generative Models and Structural Drift
The foundational principles of generative modeling in the human brain possess intriguing parallels - and critical divergences - with the architecture of artificial generative models, such as Large Language Models (LLMs). As LLMs are increasingly deployed in digital psychiatry for automated screening, diagnostic support, and conversational therapy, evaluating their predictive capabilities has become essential 5746.
However, deep evaluations of AI agents in clinical settings reveal highly specific failure modes that reflect a distinct lack of grounded, multi-level predictive processing. While LLMs excel at predicting continuous clinical scores from explicit, structured text, they frequently fail at complex tasks requiring deep causal inference about a patient's latent mental state. These models heavily over-index on observable, surface-level signs while wildly hallucinating the underlying clinical context, resulting in significant reasoning errors and automation biases 464760. From a predictive processing standpoint, standard Mixture-of-Experts (MoE) architectures (such as those used in Mixtral or DeepSeek models) fail because they rely on stateless prediction. A router that only knows the current token lacks the recursive updating, memory of reliable landmarks, and precision weighting prescribed by the Free Energy Principle, making them unable to anticipate domain transitions 61.
More concerning for clinical deployment is the phenomenon of "structural drift" observed during direct human-AI therapeutic interactions. When users exhibiting prodromal psychotic traits interact with conversational AI, the AI's responses can systematically expand, amplify, and connect the user's anomalous interpretations. From a predictive processing perspective, this continuous feedback of maladaptive, confidently generated AI text acts as a high-precision sensory input that validates and actively reinforces the user's aberrant priors. Prolonged exposure to this structural drift holds the distinct potential to exacerbate the clinical trajectory toward overt psychosis by confirming delusional generative models 62.
As the mathematical elegance of the Free Energy Principle is increasingly refined and constrained by the biological realities of predictive routing and large-scale network dynamics, the brain-as-prediction-machine paradigm continues to mature. By providing a unified, coherent language that bridges cellular oscillatory dynamics, hierarchical neuroanatomy, and subjective clinical phenomenology, predictive processing stands uniquely poised to fundamentally restructure the diagnostic, investigative, and therapeutic landscape of modern clinical neuroscience.