Physiological effects of loneliness and social isolation
The profound impact of human social connection on morbidity, mortality, and physiological homeostasis has emerged as one of the most critical paradigms in contemporary public health. In recent years, the intersection of psychosomatic medicine, social epidemiology, and molecular biology has illuminated how the presence or absence of social bonds becomes biologically embedded, altering the trajectory of chronic disease. Recognizing the magnitude of this crisis, the World Health Organization (WHO) launched the Commission on Social Connection, declaring social isolation and loneliness a global public health priority that must be addressed alongside traditional biomedical risk factors 12. According to the Commission's 2024 - 2025 flagship reports, one in six people worldwide experiences loneliness, a condition linked to an estimated 871,000 deaths annually - approximately 100 deaths every hour 234.
Despite the growing recognition of the "loneliness epidemic," which was starkly exacerbated by the structural and societal shifts of the COVID-19 pandemic, significant conceptual and mechanistic ambiguities persist in the clinical literature. For decades, social isolation and loneliness were frequently conflated in observational studies, yet contemporary evidence reveals that they represent distinct phenomena with diverging biological and epidemiological signatures. Furthermore, the temporal nature of loneliness - ranging from an acute, adaptive evolutionary signal to a chronic, toxic state of allostatic load - dictates drastically different physiological responses.
The foundational meta-analyses conducted by Julianne Holt-Lunstad established that the mortality risk of social disconnection is comparable to smoking fifteen cigarettes a day and exceeds the risks of physical inactivity and obesity 56. Updated research demonstrates that medical patients who receive psychosocial support in addition to standard treatment exhibit 20% increased odds of survival, and the broader association between social connection and survival may be as high as 50% 1. The economic and societal burdens are similarly staggering, prompting unprecedented legislative actions, such as Japan's enactment of the Act on Promotion of Measures Against Loneliness and Isolation in April 2024, the first comprehensive law of its kind, acknowledging that 39.3% of its citizens report experiencing loneliness and identifying over 21,000 "solitary deaths" in a single year 3.
This comprehensive report synthesizes post-pandemic epidemiological data, advanced Mendelian randomization studies, and high-resolution biomarker research from leading medical journals - including The Lancet, JAMA, Nature Medicine, and Psychosomatic Medicine - to delineate the precise biological impacts of social disconnection. By moving beyond general inflammatory markers to investigate specific neuroendocrine, autonomic, and circadian pathways - specifically hypothalamic-pituitary-adrenal (HPA) axis dysregulation, sympathetic nervous system (SNS) overactivity, and sleep fragmentation - this analysis provides an exhaustive map of the mechanisms linking subjective loneliness and objective isolation to global cardiovascular, neurological, and immunological disease risks.
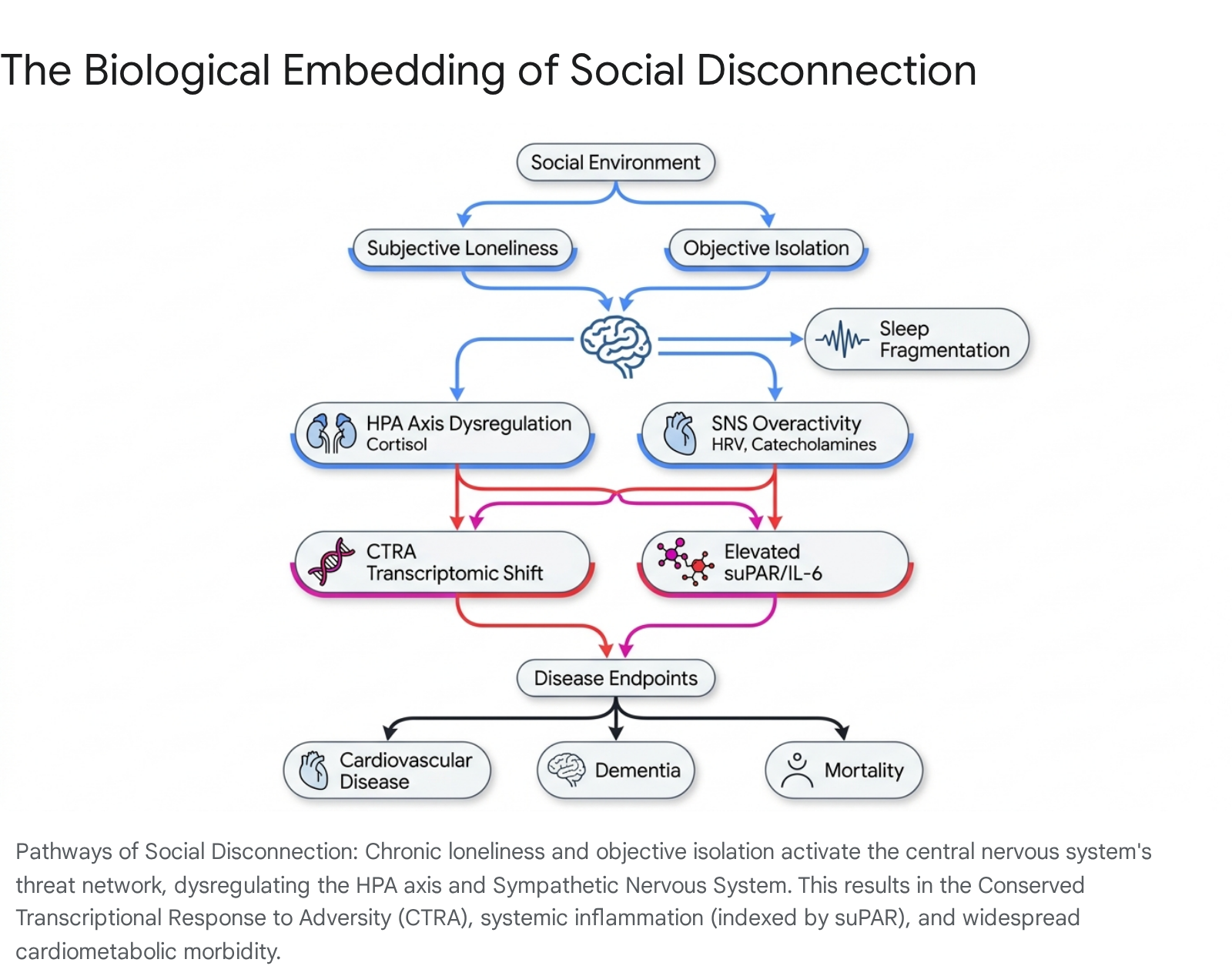
Differentiating the Typologies of Social Disconnection
To accurately map the biological sequelae of social disconnection, it is imperative to distinguish between the objective reality of one's social network and the subjective interpretation of one's social reality. These two states do not reliably overlap; highly socially integrated individuals can experience profound loneliness, while individuals with minimal social contact may feel highly fulfilled 7.
Subjective Loneliness vs. Objective Social Isolation
Social isolation is defined as an objective, observable state characterized by a pervasive lack of social proximity, sparse social networks, and limited physical engagement with others 89. It is a quantifiable metric of physical and structural disconnection. In contrast, loneliness is a subjective, distressing psychological state that arises from a perceived discrepancy between an individual's desired and actual social relationships 89.
Epidemiological and molecular evidence increasingly suggests that these two constructs exert differential impacts on human health, affecting different stages of disease progression. A highly nuanced 2025 longitudinal study published in The Journal of Epidemiology & Community Health utilizing progressive multistate Markov proportional hazards models on 236,879 individuals from the UK Biobank clarified this divergence. The analysis revealed that subjective loneliness is more strongly associated with the transition from a healthy state to the onset of an adverse health condition (Hazard Ratios ranging from 1.12 to 1.17 for loneliness, compared to 1.05 to 1.08 for isolation) 10. However, once a clinical disease is established, objective social isolation becomes the dominant predictor of subsequent transitions to mortality. The hazard ratios for transitioning from an adverse health condition to death were significantly larger for social isolation (1.28 to 1.42) than for loneliness (1.05 to 1.13) 10.
This dichotomy underscores the mechanistic differences between the two states. Subjective loneliness acts as a primary psychological stressor that initiates systemic physiological dysregulation, driving the early pathogenesis of cardiometabolic and neurological diseases 17. Objective isolation, on the other hand, deprives individuals of critical behavioral and structural buffers. Isolated individuals lack the social support networks necessary to encourage medical adherence, monitor health deterioration, or provide immediate emergency assistance (e.g., during an acute myocardial infarction or stroke), thereby drastically increasing terminal mortality rates once an illness has begun 71112.
The Evolutionary Trajectory: Acute Adaptation to Chronic Toxicity
The physiological impact of loneliness is highly dependent on its duration and chronicity. Evaluated through an evolutionary lens, acute loneliness is not an inherent pathology but an adaptive, pro-survival mechanism. Similar to physical pain, acute loneliness serves as an aversive biological signal indicating that an individual's social bonds - which historically ensured physical safety, resource acquisition, and mutual defense against predators - are insufficient or damaged 131415. This transient state triggers temporary physiological arousal and heightens social monitoring, motivating the individual to repair or seek out new social connections. Once reintegration occurs, the biological perturbation resolves without long-lasting physiological damage 131516.
However, when an individual is unable to resolve this deficit, loneliness transitions from an adaptive signal to a toxic, chronic state. The Social Safety Theory, posited by Dr. George Slavich, provides a foundational framework for understanding this shift. The theory hypothesizes that developing and maintaining friendly social bonds is a fundamental organizing principle of human behavior, and the human brain and immune system are principally designed to monitor the environment for physical and social threats 161718.
When social disconnection is sustained, chronic loneliness induces a maladaptive cognitive shift. The central nervous system shifts into a state of implicit hypervigilance for social threats. Individuals develop hostile attribution biases, becoming overly sensitive to cues signaling interpersonal rejection or exclusion 131419. The brain's neurophysiology alters to expect danger, trapping the individual in a self-perpetuating regulatory loop where they anticipate negative social interactions and subsequently withdraw from the very social networks they biologically require 131418. It is this chronic, unrelenting activation of neural threat-response systems that constitutes the primary driver of disease, resulting in allostatic overload - a systemic wear-and-tear on the body's regulatory systems that accelerates biological aging and clinical morbidity 202122.
Primary Physiological Pathways of Social Disconnection
The biological embedding of chronic loneliness and social isolation is primarily mediated through the persistent activation of three intersecting systems: the hypothalamic-pituitary-adrenal (HPA) axis, the sympathetic nervous system (SNS), and sleep architecture.
Hypothalamic-Pituitary-Adrenal (HPA) Axis Dysregulation
The HPA axis is a central neuroendocrine mediator of the human stress response. In cases of acute social isolation or transient loneliness, the HPA axis is appropriately activated to mobilize energy resources. The hypothalamus secretes corticotropin-releasing hormone (CRH), which stimulates the release of adrenocorticotropic hormone (ACTH) from the anterior pituitary gland, ultimately resulting in the secretion of glucocorticoids, notably cortisol, from the adrenal cortex 2223. This acute cortisol spike serves an adaptive function, enhancing vigilance, modifying glucose metabolism, and preparing the body for potential immediate threats 22.
Chronic loneliness, however, fundamentally dysregulates this axis. Sustained perceptions of social isolation lead to profound alterations in diurnal cortisol rhythms. In healthy populations, cortisol levels peak shortly after awakening and gradually decline throughout the day. Chronically lonely individuals, by contrast, frequently exhibit a flattened diurnal cortisol slope - meaning cortisol levels do not drop as precipitously as they should - alongside an exaggerated cortisol awakening response (CAR) and increased total daily cortisol output 222425.
Paradoxically, while chronic loneliness elevates total circulating cortisol, it simultaneously induces glucocorticoid resistance at the cellular level 2227. Target tissues, particularly immune cells, become less responsive to cortisol signaling. This resistance is partially mediated by altered interactions between the glucocorticoid receptor (GR) and regulatory co-chaperones, such as FK506 binding protein 5 (FKBP5), which modulates receptor sensitivity 27. Because cortisol normally suppresses inflammatory cascades by interfering with pro-inflammatory transcription factors, glucocorticoid resistance effectively removes the brakes on the immune system. The result is uninhibited, chronic systemic inflammation driven by unabated stress hormone exposure 2226.
Sympathetic Nervous System (SNS) Overactivity
Parallel to the neuroendocrine dysregulation is the hyperactivity of the sympathetic nervous system and the sympatho-adrenal medullary (SAM) axis. The perception of social threat triggers the classical "fight or flight" response, increasing catecholamine signaling (the release of norepinephrine and epinephrine), elevating heart rate, increasing respiratory rates, and driving blood pressure higher 222327.
A critical biomarker of autonomic nervous system (ANS) health in the context of loneliness is Heart Rate Variability (HRV). HRV measures the physiological variation in the time interval between consecutive heartbeats, serving as a reliable index of the parasympathetic nervous system's ability to regulate and calm the heart (the "rest and digest" function) 28. High HRV indicates a robust, adaptable autonomic nervous system, while low HRV indicates sympathetic dominance and stress 28.
Chronic loneliness has been consistently associated with lower resting HRV and, crucially, blunted HRV reactivity during acute stress 282930. Studies utilizing paradigms like the Trier Social Stress Test (TSST) while recording electrocardiogram (ECG) signals have demonstrated that lonely individuals suffer from a diminished ability to engage parasympathetic withdrawal and subsequent recovery 29. Evaluating specific metrics such as the root mean square of successive differences (RMSSD) and high-frequency HRV (HF-HRV) reveals that the lonely autonomic nervous system loses its regulatory elasticity 2930. This unrelenting sympathetic dominance and elevated vascular tone generate continuous hemodynamic shear stress on the vascular endothelium. Over time, this mechanical and chemical stress accelerates atherosclerosis, compromises arterial compliance, and significantly elevates the risk for incident coronary heart disease, hypertension, and ischemic stroke 1331.
Sleep Fragmentation and Circadian Disruption
Sleep represents the primary physiological period of parasympathetic dominance, cellular repair, and neurological maintenance. Evolutionary models suggest that sleeping in a socially isolated state historically increased an individual's vulnerability to predation and hostile encounters. Consequently, the brain embedded an implicit requirement for hypervigilance when isolated 732. Translating to the modern environment, subjective loneliness profoundly disrupts sleep architecture, acting as a prominent mechanism of biological weathering.
While lonely individuals may not necessarily experience a reduction in total sleep time or prolonged sleep onset latency, they suffer from significantly compromised sleep efficiency and pronounced sleep fragmentation 333437. Extensive actigraphy data - utilizing wearable, research-grade accelerometers - reveals that loneliness is robustly associated with increased Wake After Sleep Onset (WASO), which is a metric capturing micro-arousals and physical restlessness throughout the night 3735.
This pattern constitutes what recent analyses in the UK Biobank term the "Unfavorable Sleep Profile" (USP), an integrated multidimensional sleep metric characterizing poor rhythmicity, high fragmentation, and low efficiency 36. This poor sleep architecture operates as a potent secondary stressor. The lack of deep, restorative slow-wave sleep prevents the natural nocturnal dipping of blood pressure and heart rate, perpetuating the state of cardiovascular strain and allostatic overload 73334. Furthermore, the bidirectional nature of this pathway means that sleep fragmentation subsequently impairs emotion regulation and social motivation the following day. Sleep-deprived individuals exhibit altered facial expressions that make them appear less approachable, and they are faster to mislabel ambiguous emotional expressions as negative threats, making them more sensitive to social exclusion and further entrenching them in the loneliness regulatory loop 1937.
Transcriptomic Signatures and Novel Inflammatory Biomarkers
The downstream consequence of HPA axis dysregulation, sympathetic nervous system overactivity, and sleep fragmentation is a uniquely toxic inflammatory milieu. Historically, psychosomatic research relied heavily on acute-phase reactants like high-sensitivity C-reactive protein (hsCRP) and Interleukin-6 (IL-6) to measure the immunological impact of psychosocial stress. While numerous cross-sectional studies observe elevated IL-6 and CRP in lonely populations 38, these traditional markers present methodological limitations. They are highly susceptible to acute infections, minor physical injuries, diurnal variations, and singular transient stressors, which has occasionally led to inconsistent findings across different cohorts and age groups.
The Shift to suPAR as an Index of Chronic Inflammation
To overcome these limitations, recent high-tier longitudinal studies published between 2023 and 2025 have expanded the mechanistic scope to identify more stable, long-term indicators of systemic chronic inflammation. The most significant advancement in this domain is the focus on the soluble urokinase plasminogen activator receptor (suPAR) 1239. Unlike CRP and IL-6, suPAR provides a highly reliable index of cumulative, long-term immune activation, serving as a superior proxy for biological weathering and inflammatory burden 1240.
Compelling data from diverse populations - including the Danish TRIAGE study of acutely admitted medical patients, the New Zealand Dunedin Longitudinal Study, and the UK E-Risk Longitudinal Twin Study - have demonstrated that social isolation is robustly and independently associated with elevated suPAR levels 1239. The longitudinal findings are particularly striking: objective social isolation measured prospectively during childhood predicted significantly elevated suPAR levels in adulthood at age 45, even after controlling for childhood socioeconomic status, adult body mass index, smoking, and concurrent depression 1239. This highlights a profound biological latency effect, wherein early-life structural disconnection embeds a lifelong trajectory of systemic chronic inflammation.
Similarly, participants in the Dunedin cohort who reported subjective loneliness at age 38 or 45 demonstrated distinctly elevated suPAR levels at age 45 12. These elevated suPAR levels act as a harbinger of severe longitudinal outcomes. Extensive literature links heightened suPAR to accelerated plaque formation, cardiovascular mortality, hospital readmissions, and rapid biological aging as measured by epigenetic clocks like DunedinPACE 404142. The identification of suPAR bridges the gap between psychosocial distress and terminal disease, confirming that the inflammatory state induced by isolation is chronic, structural, and deeply embedded.
The Conserved Transcriptional Response to Adversity (CTRA)
At the genomic level, the biological embodiment of loneliness is characterized by a specific transcriptomic profile known as the Conserved Transcriptional Response to Adversity (CTRA) 2226. Stimulated primarily by sympathetic nervous system output (specifically the binding of norepinephrine to beta-adrenergic receptors on immune cells) and permitted by the loss of inhibitory control due to glucocorticoid resistance, the CTRA involves a distinct, systemic shift in leukocyte basal gene expression.
Molecular analyses reveal that plasmacytoid dendritic cells and monocytes in lonely individuals exhibit a marked upregulation of pro-inflammatory genes. This upregulation is driven by transcription factors such as nuclear factor kappa B (NF-κB) and activator protein-1 (AP-1) 222726. Simultaneously, there is a profound downregulation of genes responsible for Type I interferon antiviral responses and antibody synthesis 222726.
This specific transcriptomic signature reveals the evolutionary logic of the isolated body. When an individual is socially isolated, the physiological assumption is an increased risk of physical trauma, wounding, and bacterial infection (historically mitigated by the protection of the group), while the risk of transmissible viral agents (which require close social contact to spread) is deemed low 1819. The body proactively shifts its immune resources to prepare for bacterial wounding while sacrificing viral defense mechanisms. In the modern context, however, this CTRA profile is maladaptive. It directly links subjective loneliness to an increased susceptibility to chronic, sterile inflammation-driven diseases (e.g., atherosclerosis, type 2 diabetes, neurodegeneration) and leaves the individual paradoxically vulnerable to viral challenges when they do occur 182743.
Mapping the Systems, Biomarkers, and Risks
To synthesize the complex pathophysiology detailed in the preceding sections, the following table maps the core biological systems implicated in social disconnection, their specifically observed biomarkers, the mechanism of dysregulation, and the associated disease trajectories.
| Biological System | Specifically Observed Biomarkers | Manifestation of Dysregulation | Associated Disease Risks |
|---|---|---|---|
| Hypothalamic-Pituitary-Adrenal (HPA) Axis | Salivary/Hair Cortisol, Glucocorticoid Receptor (GR) sensitivity | Flattened diurnal cortisol slope; elevated Cortisol Awakening Response (CAR); cellular glucocorticoid resistance via FKBP5 alteration. | Metabolic syndrome, cognitive decline, impaired stress recovery, chronic inflammatory states. |
| Sympathetic Nervous System (SNS) | Catecholamines (Epinephrine, Norepinephrine), Heart Rate Variability (RMSSD, HF-HRV) | Reduced resting HRV; blunted parasympathetic withdrawal under acute social stress; continuous hemodynamic shear stress. | Essential hypertension, coronary heart disease, endothelial dysfunction, ischemic stroke. |
| Immune System (Transcriptomic) | CTRA Gene Expression Profile (NF-κB, AP-1) | Upregulation of pro-inflammatory genes; profound downregulation of Type 1 interferon antiviral responses. | Increased vulnerability to viral infections, impaired antibody synthesis, accelerated biological aging. |
| Immune System (Systemic) | suPAR, IL-6, hsCRP, Fibrinogen | Persistent elevation of systemic chronic inflammation markers; suPAR acts as a stable index of long-term immune weathering. | Atherosclerosis, type 2 diabetes, cancer progression, all-cause mortality, frailty. |
| Circadian Rhythm / Sleep Architecture | Actigraphy metrics: Wake After Sleep Onset (WASO), Sleep Efficiency | High sleep fragmentation; decreased sleep efficiency; lack of nocturnal blood pressure dipping due to implicit hypervigilance. | Cognitive impairment, dementia progression, secondary allostatic load, impaired social cognition. |
Post-Pandemic Epidemiological Evidence (2023 - 2026)
The empirical validation of these physiological mechanisms is heavily supported by massive, globally diverse longitudinal studies published in the wake of the COVID-19 pandemic. The pandemic served as an unprecedented global natural experiment, severely restricting physical proximity and accelerating research into the cardiovascular and neurological endpoints of prolonged disconnection 4. Publications in The Lancet and JAMA over the last three years have provided granular insights into how universally toxic loneliness is across disparate cultures.
Cardiovascular and Cardiometabolic Outcomes
Recent multi-cohort epidemiological studies highlight the uniform toxicity of loneliness, dismantling the assumption that cultural norms might buffer its impact. A robust 2023 analysis compared longitudinal data from the Health and Retirement Study (HRS) in the United States (n=13,073) to the Korean Longitudinal Study of Aging (KLoSA) in South Korea (n=8,311), specifically tracking the incidence of fatal and non-fatal cardiovascular disease (CVD) 4448. Despite the stark cultural differences between the individualistic framework of the U.S. and the collectivist framework of South Korea, loneliness increased the odds of experiencing an incident CVD event by an almost identical margin. In the fully adjusted models (controlling for age, income, physical activity, and smoking), loneliness was associated with an 18% higher likelihood of developing CVD in the U.S. cohort and a 16% higher likelihood in the South Korean cohort 4448.
These trans-cultural findings are further supported by prospective data from the China Health and Retirement Longitudinal Study (CHARLS). Tracking over 10,668 middle-aged and older adults over an 8.9-year median follow-up, researchers found that lonely individuals exhibited a 24% increased risk of incident heart disease and a 26% increased risk of stroke compared to their non-lonely counterparts 45. Data from the Survey of Health, Ageing and Retirement in Europe (SHARE) and the English Longitudinal Study of Ageing (ELSA) corroborate these metrics, emphasizing that subjective loneliness predicts incident coronary heart disease independent of traditional clinical risk factors across Europe 3850.
At the molecular level, proteomic analyses published in Nature Human Behaviour in 2025 utilizing the UK Biobank (n=42,000) mapped the proteins connecting social disconnection to poor health. The team identified 175 specific proteins associated with objective social isolation and 26 proteins associated with subjective loneliness 4647. A critical finding was the elevation of Adrenomedullin (ADM) in lonely individuals. ADM is known to regulate stress and social hormones, including oxytocin. The researchers found a strong inverse association between elevated ADM levels and the volume of the insula, a key brain region for interoception and emotional regulation, providing direct molecular and anatomical evidence connecting social network deprivation to metabolic disruptions like type 2 diabetes and stroke 47.
Cognitive Decline and Neurological Deterioration
The impact of social disconnection on brain health is equally profound and rapidly progressive. Loneliness is increasingly recognized as a primary, modifiable risk factor for dementia. Data from the Framingham Heart Study revealed that baseline loneliness conferred a 54% increased 10-year risk for incident dementia 222748. Crucially, this risk is highly age-dependent and interacts significantly with genetics; the risk skyrocketed to a three-fold increase among younger-old adults (aged 60-79) who lacked the APOE-ε4 genetic variant, suggesting that loneliness can independently drive neurodegeneration even in populations without standard genetic predispositions 2227.
Furthermore, comprehensive meta-analyses involving over 600,000 individuals have confirmed that loneliness increases the risk for all-cause dementia by approximately 30%, Alzheimer's disease by 39%, and vascular dementia by an alarming 73% 2049. Neurologically, chronic loneliness accelerates gray matter loss and alters connectivity in critical circuits, particularly the default mode network and the ventral tegmental area-nucleus accumbens pathway, which governs reward processing 224950. The degradation of these pathways drives the anhedonia and cognitive withdrawal frequently seen in isolated older adults. Furthermore, the elevated systemic inflammation (indexed by markers like suPAR) acts synergistically with sleep fragmentation to accelerate the breakdown of the blood-brain barrier and the accumulation of neurotoxic amyloid and tau proteins 485651.
Mortality and Survival Rates
Julianne Holt-Lunstad's foundational meta-analyses established the dire mortality risks associated with poor social connection 56. Updated 2024-2025 longitudinal analyses in JAMA and Psychosomatic Medicine confirm and extend these figures. In the general population, objective social isolation increases the risk of all-cause mortality by approximately 32%, and cancer mortality by 24% 156. Subjective loneliness independently elevates overall mortality risk by 14% to 23% depending on chronicity and severity 1556.
Among highly vulnerable clinical populations, such as cancer survivors, loneliness exerts a clear dose-response relationship with early mortality 56. The unique burden of cancer survivorship - including the stigma of physical changes, prolonged medicalization, and fear of recurrence - fosters deep existential isolation that severely hampers physiological recovery, immune surveillance, and adherence to medical treatment protocols, directly contributing to excess mortality 52.
Methodological Limitations: Confounding, Reverse Causality, and Genetics
While the epidemiological evidence linking social disconnection to disease is vast, the field of social epidemiology has historically faced significant methodological challenges. Chief among these are the presence of confounding variables and the risk of reverse causality. Critics have long argued that poor health may cause social isolation (e.g., an individual immobilized by a stroke or severe arthritis loses their social network), rather than isolation causing the physical disease 1241.
Disentangling Loneliness from Depression
A major specific confounding factor in loneliness research is Major Depressive Disorder (MDD). Loneliness and depression are highly comorbid, share similar behavioral manifestations (e.g., social withdrawal, fatigue, anhedonia), and are both linked to elevated systemic inflammation (such as elevated IL-6 and CRP) 4859. Historically, it was difficult to determine if loneliness was merely a sub-symptom of depression or a distinct pathogenic entity.
Advanced longitudinal and genetic studies have definitively proven that while they overlap, they are distinct constructs 4853. While depression can precede and precipitate social isolation, loneliness is recognized globally as an independent, modifiable risk factor. For instance, in both the UK Biobank and Framingham studies, the correlation between loneliness and the onset of dementia and cardiovascular mortality persists robustly even after rigorously adjusting for baseline depressive symptomatology and anti-depressant medication use 1248.
Insights from Mendelian Randomization
To conclusively bypass the issues of reverse causality and environmental confounding, researchers have increasingly utilized Mendelian Randomization (MR). MR acts as a genetic natural experiment, utilizing inherited genetic variants (Single Nucleotide Polymorphisms, SNPs) as instrumental variables to assess causal relationships, as genetic predispositions are assigned randomly at conception and are immune to subsequent disease onset or lifestyle factors 54.
Recent bidirectional, two-sample MR studies utilizing genome-wide association study (GWAS) data from over 450,000 individuals of European descent have provided groundbreaking causal evidence. These analyses demonstrated a significant, unidirectional causal effect of genetically predicted loneliness on the risk of developing essential hypertension (OR = 1.07), atherosclerotic heart disease (OR = 1.04), angina, and ischemic heart disease (IHD) 5455. Network MR studies revealed that the causal pathway from loneliness to IHD is significantly mediated by downstream metabolic alterations, including increased body fat percentage, waist-hip ratio, total cholesterol, and elevated LDL cholesterol 55. Crucially, reverse causality models in these specific cardiovascular studies showed that ischemic heart disease does not cause the genetic predisposition for loneliness, confirming that loneliness serves as the upstream antecedent to the cardiovascular pathology 55.
Furthermore, bidirectional MR analyses evaluating Severe Mental Illness (SMI) confirm a causal, risk-increasing effect of loneliness on the development of Major Depressive Disorder and Schizophrenia. This relationship appears to be partially mediated by systemic inflammatory signaling 5963. Unlike the cardiovascular findings, the reverse causality models in psychiatric evaluations showed that while depression causes loneliness, loneliness independently causes depression, indicating a vicious, bidirectionally causal cycle of psychological and biological degradation 5963.
Global Public Health Directives and Clinical Implications
The profound biological embedding of loneliness necessitates an immediate paradigm shift in how global health systems address social needs. The 2024 - 2025 flagship reports from the WHO Commission on Social Connection emphasize that "social health" must be recognized as the missing third pillar of human wellness, standing co-equally with physical and mental health 5657. The economic burden is staggering, with loneliness costing societies billions annually in increased healthcare utilization, emergency department visits, lost economic productivity, and the long-term care required for accelerated dementia 24.
Recognizing loneliness and isolation as modifiable clinical risk factors requires the integration of psychosocial screening directly into routine primary care 6. Because objective isolation and subjective loneliness operate via fundamentally different psychological and biological mechanisms, clinical interventions must be precision-targeted.
For objective social isolation, structural interventions such as "social prescribing" have shown immense promise. Social prescribing enables healthcare professionals to refer patients to non-medical community services - such as arts programs, cultural activities, and local volunteer networks - to rebuild social capital 58. This approach has been particularly effective in rebuilding functional independence and reducing mortality risks in the WHO Western Pacific regions, where rapidly aging populations face acute structural isolation 58.
However, for subjective, chronic loneliness, simply placing an individual in a room with others is biologically insufficient and may even exacerbate stress. Because chronic loneliness involves entrenched "Social Safety" threat schemas, hostile attribution biases, and autonomic hypervigilance, interventions must focus on psychological and cognitive restructuring. Meta-regressions of randomized controlled trials demonstrate that therapies focusing on distress tolerance and Cognitive Behavioral Therapy (CBT) aimed at correcting maladaptive social cognition yield the highest effect sizes in reducing subjective loneliness 6225960. By addressing the underlying cognitive biases that sustain the HPA axis and sympathetic threat responses, these targeted therapies can effectively deactivate the physiological alarm system.
Conclusion
The distinction between objective social isolation and subjective loneliness represents a critical frontier in psychosomatic medicine and public health. While structural isolation deprives the human body of essential behavioral supports and emergency physical buffers - driving up terminal mortality rates - subjective loneliness orchestrates a devastating, top-down biological assault. By triggering an evolutionary mismatch, chronic loneliness traps the brain in a state of perceived social threat, sustaining severe HPA axis dysregulation, persistent sympathetic nervous system overactivity, and profound circadian fragmentation. These forces converge to induce the Conserved Transcriptional Response to Adversity (CTRA) and elevate stable chronic inflammatory markers like suPAR, quietly and persistently accelerating the pathogenesis of cardiovascular disease, dementia, and premature mortality.
Armed with definitive causal evidence from Mendelian randomization and massive post-pandemic longitudinal cohorts spanning diverse global cultures, the medical community can no longer view social connection as a peripheral quality-of-life metric. It is a fundamental, measurable biological imperative. Mitigating the global epidemic of social disconnection requires a unified approach, integrating structural community investments with targeted psychotherapeutic interventions to dismantle the biological embedding of loneliness, ultimately charting a path to healthier, more resilient societies.