Neuroscience and socioeconomic factors of habit formation
The mechanisms governing human behavioral change - specifically the transition from conscious, goal-directed action to automated, habitual routine - represent one of the most intensively studied and critically debated domains within cognitive neuroscience, health psychology, and behavioral economics. For decades, the paradigm of self-improvement and behavioral modification has been disproportionately influenced by highly marketable, yet scientifically truncated, pop-psychology frameworks. These commercial models frequently reduce the profound neurobiological and sociological complexities of habituation to simplistic, individualistic formulas. While such frameworks have effectively democratized behavioral theory for mass consumption, they dangerously strip the discipline of its necessary empirical rigor and contextual nuance 123.
Current peer-reviewed research, spanning longitudinal field studies and advanced neuroimaging from 2023 to 2026, highlights a critical epistemological inflection point. The scientific consensus, championed by foundational researchers such as Wendy Wood, David Neal, and Phillippa Lally, is actively diverging from derivative self-help literature 123. The contemporary empirical approach moves toward a multi-dimensional model that meticulously integrates neural circuit hand-offs, systemic and environmental friction, cross-cultural cognitive variability, and advanced digital intervention scaling 45910.
To comprehensively understand behavior change in the modern era, it is necessary to dismantle prevailing commercial myths - such as the arbitrary 21-day habit formation timeline, the linear compounding of incremental behavioral changes, and the metabolic resource model of ego depletion. In their place, a rigorous reliance on peer-reviewed data must be established. This exhaustive analysis provides a synthesis of contemporary updates to dual-system theory, details the precise neurological interplay between the prefrontal cortex and the basal ganglia, and establishes the profound impact of socioeconomic status (SES) and environmental friction on the individual's capacity to sustain goal-directed behavior 67138.
Neurological Foundations: The Transition from Goal-Directed to Habitual Control
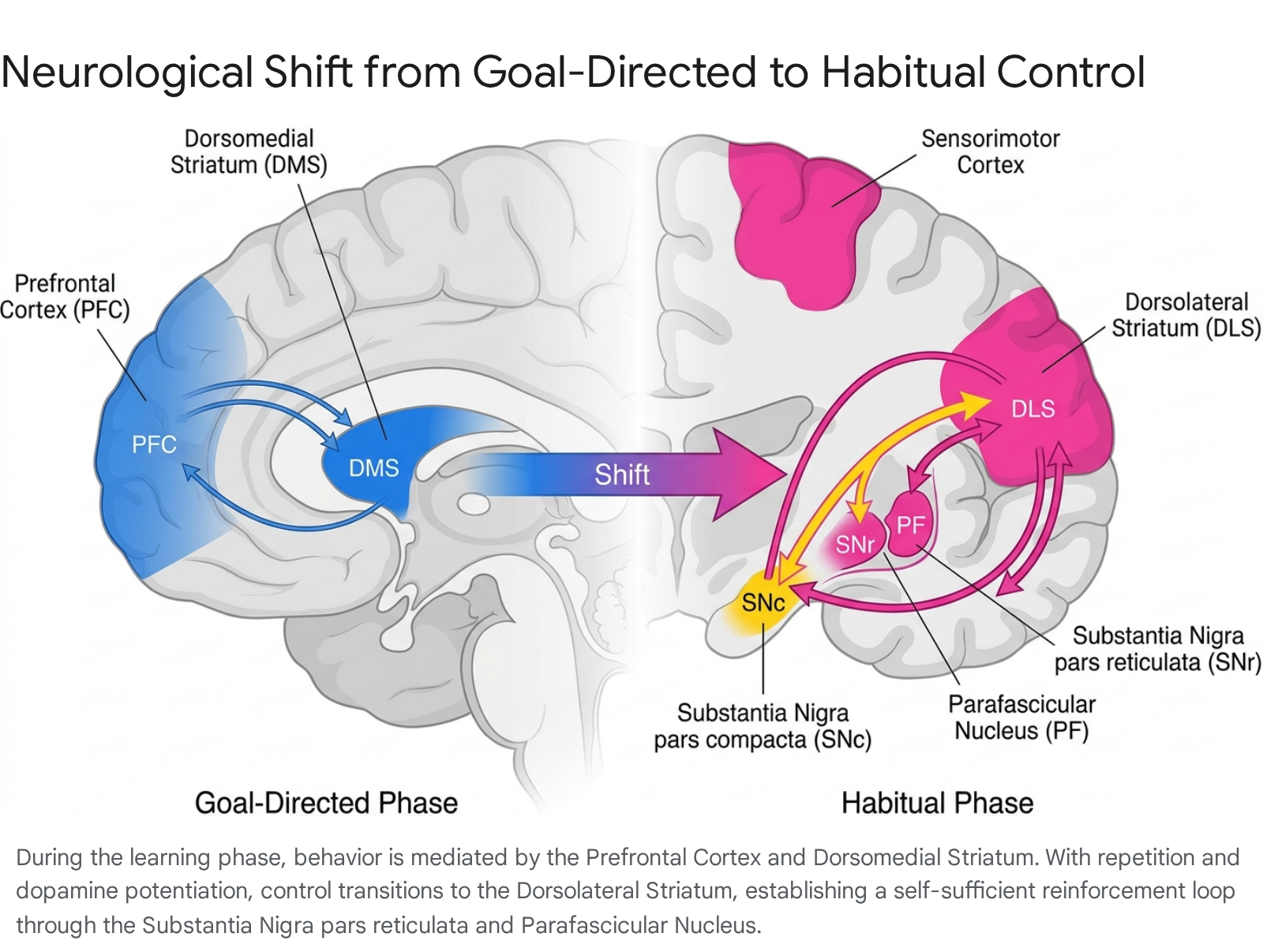
The traditional dual-system model of behavior posits a fundamental computational dichotomy between a goal-directed system and a habitual system. The goal-directed system is characterized as flexible, computationally demanding, and highly sensitive to immediate outcome valuation. Conversely, the habitual system is characterized as rigid, computationally efficient, and largely outcome-insensitive, relying on historical reinforcement rather than active decision-making. Recent neuroimaging, functional magnetic resonance imaging (fMRI), and optogenetic studies have substantially refined our understanding of the specific anatomical structures and synaptic alterations that facilitate the transition between these two modes of control 491011.
The Prefrontal-Striatal Hand-Off Mechanism and Subcortical Specialization
During the initial stages of learning a novel behavior, an organism relies heavily on the prefrontal cortex (PFC), specifically the ventrolateral and dorsolateral prefrontal cortices (vlPFC and dlPFC), working in tandem with the dorsomedial striatum (DMS), which corresponds to the caudate nucleus in humans 11121314. The motor cortex (M1 and M2) provides a vital "template" for the required movement, coordinating the precise timing of neuronal activation within the striatum 4. This early phase is highly susceptible to cognitive load and requires continuous outcome valuation and error correction.
Recent literature identifies the vlPFC as a critical neural substrate responsible for temporally multiplexing fast and slow learning processes. The vlPFC manages the inherent tension between the flexibility needed to adapt to new rewards and the stability required to retain functional habits 13. As learning progresses, a sophisticated division of labor emerges within the striatum along a rostro-caudal axis. The rostral caudate head (CDh) mediates adaptability through fast value learning and rapid forgetting, making it ideal for navigating volatile environments. In contrast, the caudate tail (CDt) mediates long-term stability by engaging in slow value learning and slow forgetting, thereby preserving object-reward associations over extended periods 1314.
As a behavior is repeatedly executed in a stable context and consistently followed by a reward, a neurobiological "hand-off" occurs. Control transfers from the computationally expensive cortico-basal ganglia loops to more localized, efficient subcortical circuits. The primary site of habit storage shifts definitively to the dorsolateral striatum (DLS), corresponding to the putamen in the human brain 491011. The DLS contains highly specific microdomains representing distinct body regions and motor sequences. Once a behavior is fully automated, the cortical areas originally required for its initiation and timing can be transiently inactivated without impairing the flawless execution of the motor program 4.
The Reinforcement Loop, Thalamostriatal Synapses, and Dopaminergic Potentiation
The precise mechanism enabling this profound cortical independence is the strengthening of the DLS-SNr-PF-DLS neurocircuitry loop 4. During the learning phase, the motor cortex activates specific populations of striatal projection neurons (SPNs) within the DLS. These SPNs inherently inhibit the Substantia Nigra pars reticulata (SNr). The SNr functions as the primary output nucleus of the basal ganglia, acting as a "keyboard for movement control" that tonically inhibits downstream motor centers. Therefore, its inhibition by the DLS results in a critical disinhibition of the Parafascicular Nucleus (PF) of the thalamus 4.
The newly disinhibited PF then sends potent excitatory signals back to the original microdomains in the DLS. Crucially, successful performance and subsequent reward perception trigger a burst of dopamine from the Substantia Nigra pars compacta (SNc). This dopamine burst potentiates the active thalamostriatal synapses 4. While early skill acquisition relies heavily on corticostriatal synaptic plasticity, the flawless execution of mature habits is largely dependent on the strengthened signal transmission within these subcortical thalamostriatal synapses. This architectural shift from cortical reliance to thalamostriatal sufficiency explains why habits are executed so rapidly, efficiently, and effortlessly: they literally bypass the metabolically expensive evaluative cortices 41516.
Historically, neuroscientists rigidly separated the nigrostriatal dopamine system (originating in the SNc and projecting to the dorsal striatum, traditionally associated strictly with motor function) from the mesolimbic system (originating in the Ventral Tegmental Area, VTA, and projecting to the ventral striatum, associated with motivation and reward) 171819. However, current empirical literature renders this strict anatomical dichotomy obsolete. Both nominal systems actively participate in reward function, memory consolidation, and habit formation. Target neurons in both the primary terminal fields of the SNc (dorsal striatum) and the VTA (ventral striatum) respond similarly to rewards and reward-predictors 171920.
Furthermore, recent developmental neurobiology studies reveal that these discrete dopaminergic pathways are established through highly specific molecular signaling. Factors released from the target tissues regulate pathway formation: BMP6/BMP2 interacting with Smad1 regulates the nigrostriatal connections, while TGFβ2 interacting with Smad2 establishes the mesolimbic connections 18. The coordinated activation of both pathways demonstrates that habit formation relies on a highly synchronized dopaminergic cascade that permanently bridges the domains of abstract motivation and automated physical motor execution.
To consolidate this complex, multi-layered neuroanatomy, the distinct neurological structures and their evolving roles during the shift from goal-directed action to automated habit are summarized in the table below.
Table 1: Neurological Structures and Their Functions within the Habit Loop
| Neurological Structure | Primary Pathway/Network | Function in Goal-Directed Phase (Learning) | Function in Habitual Phase (Automaticity) |
|---|---|---|---|
| Prefrontal Cortex (vlPFC/dlPFC) | Corticostriatal | High activation; continuous outcome valuation, conscious planning, and management of cognitive load. | Diminished involvement; actively bypassed by subcortical circuits to conserve cognitive resources. |
| Motor Cortex (M1/M2) | Corticostriatal | Provides the neural "template" and precise timing for movement sequences. | Cortical input becomes unnecessary; structures can be inactivated without impairing execution. |
| Caudate Head (CDh) | Cortico-basal ganglia | Mediates adaptability via fast value learning and rapid memory forgetting. | Function diverges from habitual stability; remains sensitive to volatile reward prediction errors. |
| Caudate Tail (CDt) | Cortico-basal ganglia | Encodes long-term value memory (LTVM); slow learning and slow forgetting. | Preserves stable object-reward associations independently of immediate performance reinforcement. |
| Dorsolateral Striatum (DLS) / Putamen | Thalamostriatal | Minimal initial role; passively receives cortical signals to activate specific projection neurons. | Primary site of habit storage; drives automated motor programs via thalamic feedback loops. |
| Substantia Nigra pars reticulata (SNr) | Basal Ganglia Output | Tonically inhibits downstream motor and cognitive centers. | Inhibited by the DLS, resulting in the disinhibition of the thalamus (PF). |
| Parafascicular Nucleus (PF) of Thalamus | Thalamostriatal | Receives disinhibitory signals from the SNr. | Sends excitatory reinforcement back to the DLS, sustaining the automated, self-sufficient loop. |
| Substantia Nigra pars compacta (SNc) | Nigrostriatal Dopaminergic | Delivers immediate dopamine bursts upon successful reward prediction and execution. | Potentiates active thalamostriatal synapses, cementing the operational independence of the circuit. |
| Ventral Tegmental Area (VTA) | Mesolimbic Dopaminergic | Signals initial reward value and motivation; initiates goal-directed action. | Works in concert with SNc to bridge motivation with automated motor consolidation. |
Data synthesized from current neuroimaging, optogenetic, lesion, and developmental studies 4910111314171819.
Updates to Dual-System Theory: The Rate-Correlation Framework
While the anatomical pathways of habit formation are becoming definitively clear, the computational logic underlying the shift from goal-directed to habitual behavior has undergone a significant theoretical update. The conventional model assumed a straightforward, zero-sum competition where habits simply overwrite goal-directed actions through sheer repetition. However, the Rate-Correlation Theory, formalized and subsequently updated by Perez and Dickinson (2020, 2024), proposes a far more nuanced, concurrent operation of both systems 62128.
Under the rate-correlation framework, the total probability of an organism responding to an environmental cue is the sum of the concurrent strengths of both the goal-directed and habitual systems. 1. The Goal-Directed System is sensitive to the macroscopic correlation between the overall rate of a specific behavior and the overall rate of reinforcement over a prolonged period. It utilizes a sophisticated local memory of time samples, constantly adding new data and randomly forgetting older data to compute this correlation mathematically 621. 2. The Habitual System, conversely, aligns directly with model-free reinforcement learning principles. It is microscopic and heavily reliant on immediate reward prediction errors. Habit strength is driven purely by the strict contiguity between a specific response and its direct reinforcement 622.
This updated model elegantly explains behavioral phenomena that classical theories fail to address. For example, it elucidates why ratio reinforcement schedules (where every nth response is rewarded) maintain goal-directed control far more effectively than interval schedules (where the first response after a set time is rewarded). In ratio schedules, increasing the response rate directly and linearly increases the reinforcement rate, keeping the rate-correlation high and the goal-directed system robustly engaged. In interval schedules, once all available reinforcers are collected in the given time frame, further responding yields zero extra reward. This drops the rate correlation, allowing the contiguity-driven, model-free habit system to rapidly dominate 623.
Crucially, the rate-correlation theory demonstrates that human goal-directed behavior is highly resistant to interventions specifically designed to break the action-outcome contingency, such as extinction training (removing the reward entirely) or non-contingent training (delivering rewards randomly, independent of action). During extinction, the habit system experiences profound negative reward prediction errors, rapidly reducing its contribution to responding. The observable cessation of behavior is not the unlearning of a goal; rather, it is the habit system masking the goal-directed system. The underlying goal-directed knowledge regarding the value of the outcome remains entirely intact and can rapidly resurface 62124.
This mechanism comprehensively explains the high relapse rates observed in clinical addiction and behavioral interventions: the automated pathway is suppressed through negative reinforcement, but the abstract goal-directed knowledge of the substance's or action's value is never erased 1921. Furthermore, human data indicates that affective stress and anxiety radically alter this balance. Individuals exhibiting high-stress states compromise the working memory required for complex rate-correlation computations, forcing the brain to default almost immediately to model-free contiguity learning. Consequently, stressed individuals develop rigid habits exponentially faster, even with minimal training 624.
The Empirical Critique of Pop-Science Self-Improvement
The profound complexity of neuroanatomy, model-free reinforcement learning, and rate-correlation theory stands in stark, unavoidable contrast to the narratives currently propagated by the multi-million-dollar self-improvement industry. Frameworks popularized by authors such as James Clear (Atomic Habits) and BJ Fogg (Tiny Habits) have commodified behavioral science. They distill complex psychological mechanisms into highly marketable, highly individualistic aphorisms that resonate with consumers but lack scientific validity 1323334.
The scientific community - particularly authoritative empirical researchers in the domain such as Wendy Wood, David Neal, and Phillippa Lally - increasingly critiques these derivative texts for their severe oversimplification, lack of empirical rigor, and total failure to account for systemic, contextual, and environmental barriers 131235. The work of Wood and Neal explicitly emphasizes that habits are an associative learning process heavily reliant on stable context cues, operating independently of personal goals once formed. In contrast, pop-science methodologies attempt to force habituation through sheer willpower and minor environmental tweaks, fundamentally misunderstanding the neurological permanence of the habit-goal divergence 12.
The Oversimplification of the Habit Loop and the Compounding Myth
The core of Atomic Habits relies heavily on a mechanistic "cue, craving, response, reward" model. While intuitively appealing to the layperson, this framework fails to capture the fluctuating emotional, psychological, and allostatic complexities required to sustain behavioral change in uncontrolled environments 125. Critics specifically target the much-touted "compound effect" - the foundational claim that getting 1% better every day leads to exponential, limitless growth. This concept lacks broad empirical support in human biology. Biological systems are governed by homeostasis, physical constraints, and diminishing returns. Infinite linear compounding is a mathematical fiction when applied to human neurophysiology and behavioral conditioning 126.
Furthermore, Fogg's Tiny Habits framework explicitly eschews rigorous, randomized controlled trials. Fogg himself has publicly stated that traditional psychology experiments are "limited" and that he intentionally set them aside when developing his methodology, relying instead on subjective self-reporting and anecdotal success stories 35. This intentional departure from the scientific method means these frameworks dangerously generalize findings from highly specific, low-stakes scenarios (e.g., flossing a single tooth or drinking a glass of water) to complex behavioral pathologies like severe addiction, binge eating disorders, or clinical depression. In these clinical contexts, simplistic "tiny" strategies are woefully inadequate and can induce feelings of profound personal failure when they inevitably stall 113538.
Debunking the "21 Days to Form a Habit" Misconception
Perhaps the most pervasive and damaging misconception amplified by pop-science is the myth that a habit is formed in exactly 21 days. This misinterpretation originates from Dr. Maxwell Maltz's 1960 observations on patients' self-image adjustments following plastic surgery, which had nothing to do with neuroplastic habituation 327. Current peer-reviewed literature definitively and repeatedly refutes this timeline.
A landmark 2024 - 2025 systematic review and meta-analysis by Singh et al., analyzing data from over 2,600 participants across 20 distinct controlled studies, found that the true median time required to reach behavioral automaticity ranges from 59 to 66 days 3538. Crucially, the range of individual variability is massive - spanning from 4 days to an astonishing 335 days. This variance depends heavily on the complexity of the behavior, the stability of the environmental context, and the individual's baseline neuroplastic capacity 3527.
The empirical data from Lally et al. further demonstrates that missing a single day of practice does not permanently derail the habit formation process, directly contradicting the strict "streak-based" tracking encouraged by commercial apps and books 1. Simple behaviors tied to existing morning routines form faster, while complex behaviors demanding high executive function - such as overhauling a diet or beginning a rigorous exercise regimen - take significantly longer (often over 150 days) and face much higher rates of abandonment 53827.
The Fall of Ego Depletion and the Shifting Model of Willpower
Pop-science literature has also heavily relied on the concept of "ego depletion" - the theory popularized in the late 1990s and early 2000s by Roy Baumeister, which asserted that willpower is a finite metabolic resource that reliably depletes with use, akin to a muscle running out of cellular glucose 728. For over a decade, this formed the basis of widespread advice suggesting that individuals should front-load important decisions in the morning or consume sugary snacks to replenish self-control reserves.
However, the field of psychology has recently undergone a severe, highly publicized replication crisis regarding ego depletion. Massive, multi-laboratory pre-registered replication attempts have consistently failed to find the ego depletion effect 414229. Researchers such as Michael Inzlicht have led the charge to dismantle the metabolic resource model entirely. The current scientific consensus has shifted away from the idea of a depleting reservoir of biological willpower. Instead, self-control fatigue is now understood as a process-oriented motivational and attentional shift 4129.
Exerting cognitive effort at Time 1 does not drain a physical resource; rather, it triggers a psychological recalibration where the brain actively devalues further effort at Time 2 and increases its attentional bias toward immediate gratification or rest 4229. This is not a failure of fuel, but a highly functional, evolutionary mechanism designed to continually assess opportunity costs and prevent the monopolization of cognitive bandwidth by any single, non-vital task.
Table 2: Contrasting Popular Self-Improvement Frameworks Against Current Scientific Consensus
| Domain | Popular Self-Improvement Narratives (Atomic Habits, Tiny Habits) | Current Scientific & Empirical Consensus (Wood, Neal, Lally, Inzlicht) |
|---|---|---|
| Habit Formation Timeline | Claims habits can be formed in exactly 21, 30, or 60 days via strict adherence and unbroken streaks. | Median is 59-66 days; massive individual variance (4 to 335 days). Missing a single day does not permanently derail neuroplastic adaptation. 13538 |
| Mechanism of Failure | Attributed to poor individual system design, lack of intrinsic motivation, or failure to make cues "obvious." | Driven by systemic friction, socioeconomic constraints (time poverty), and stress-induced shifts from vlPFC to contiguity-based DLS learning. 6138 |
| Willpower & Effort | Willpower is a depletable, metabolic resource (Ego Depletion theory); self-control must be aggressively conserved. | Ego depletion suffers a severe replication crisis. Fatigue is a functional motivational/attentional shift weighing opportunity costs, not a metabolic deficit. 414229 |
| Scalability & Scope | Suggests "1% improvements" compound linearly and can overcome severe behavioral or psychological pathologies. | Human biology is homeostatic; compounding faces diminishing returns. Simplistic behavioral loops fail against complex clinical addictions or systemic trauma. 126 |
| Data Foundation | Heavily relies on anecdotal narratives, survivor bias, and non-experimental self-reported observational claims. | Relies on systematic meta-analyses, fMRI/optogenetic tracking, multi-laboratory pre-registered studies, and large-scale field mega-studies. 924353031 |
The Paradigm of Systemic Friction: Environmental and Socioeconomic Determinants
The most glaring epistemological deficiency in individualistic self-improvement models is their total blindness to systemic friction. By localizing the responsibility for behavior change entirely within the individual's cognitive architecture, pop-science dangerously ignores the overwhelming power of the socioeconomic environment 313. The "Intention-Behavior Gap" - the measurable, observable void between stated pro-social/pro-health goals and actual lived actions - is not merely a psychological failing. It is a critical diagnostic data point revealing the failure of broader socio-ecological systems 913.
Socioeconomic Status (SES) and the Paralysis of Time Poverty
Behavioral economics and sociological research highlight that optimal habit formation is an inherently privileged endeavor. Individuals of high socioeconomic status (SES) exhibit superior regulatory functions and more efficient neurocognitive processing patterns, largely because their environments impose significantly less daily allostatic load 32. Conversely, lower-SES individuals frequently suffer from "time poverty" - the chronic, systemic deficit of discretionary time caused by extended workdays, multiple low-wage jobs, prolonged commutes, and unremunerated domestic labor 8334834.
Time poverty acts as a severe, insurmountable cognitive barrier. Formulating, executing, and monitoring goal-directed behaviors requires the metabolically expensive engagement of the prefrontal cortex 913. When an individual is chronically exhausted, stressed, or pressed for time, the brain cannot sustain System 2 deliberative thought. It inevitably defaults to familiar, low-effort System 1 pathways (habitual control), which are often the precise unsustainable or unhealthy behaviors the individual wishes to change 96.
A rigorous 2026 latent profile analysis utilizing data from over 9,800 participants in the China Family Panel Studies identified four distinct SES-time usage profiles. The "Double Poverty" subgroup (characterized by both low income and long working hours) consistently exhibited the worst health and well-being outcomes across all measured domains 48. Notably, the study revealed that temporal scarcity is often more detrimental to psychological well-being than absolute financial poverty, provided the individual has cleared baseline economic survival thresholds 48.
Structural Barriers and the Failure of Interventions
When individuals face rigid structural barriers, no amount of cognitive "habit stacking," identity reframing, or "cue optimization" can overcome the physical limits of the environment 3835. For example, a qualitative study analyzing medication adherence among hypertensive patients revealed that while cognitive forgetfulness was a minor factor, the primary barriers to habit formation were purely structural: cost-prohibitive medications, sudden insurance formulary changes, and intensive employment conditions that prevented the establishment of stable daily routines 3637.
Similarly, the precipitous dropout rates observed in youth sports during early adolescence (ages 13-15) are frequently misattributed to a psychological lack of motivation or burnout. In reality, sociological models demonstrate that these dropouts are driven by systemic friction: the clash between highly professionalized sports infrastructure, rapidly rising financial costs, and the necessary withdrawal of parental logistical support 35. Framing this as an individual failure of willpower absolves the systemic design flaws of the environment.
Furthermore, SES shapes profound, unconscious physiological responses during everyday social interactions. Unobtrusive laboratory studies demonstrate that low-SES individuals exhibit significantly stronger physiological linkage (attunement) to interaction partners than high-SES individuals 38. This suggests that individuals living in high-friction, low-resource environments adopt a more vigilant, "other-focused" cognitive stance for social survival. This constant external vigilance inherently siphons critical cognitive bandwidth away from self-directed internal habit optimization 2638. To demand perfect behavioral compounding from an individual whose cognitive architecture is continuously assaulted by systemic friction is to misapply behavioral science entirely.
Cross-Cultural Nuances in Behavioral Automaticity
The assertion that habit formation operates via universal, culturally agnostic mechanisms ignores the profound impact of cultural context on cognitive development. Executive function (EF) and the ability to delay gratification - critical cognitive components required for overriding prepotent, undesired habitual responses in favor of goal-directed actions - are deeply intertwined with, and scaffolded by, sociocultural practices 3940.
Recent cross-cultural empirical studies challenge the deficit-based narratives that are often inappropriately applied to minoritized or non-Western populations. For instance, classic delay-of-gratification tests (such as the famed Stanford marshmallow experiment) historically framed inhibitory control purely as a measure of innate, individual willpower. However, recent comparative studies between Dutch, urban Chinese, and Japanese toddlers, prominently discussed by Yanaoka et al. (2024), demonstrate that performance on these tasks is largely a reflection of culturally ingrained habits rather than isolated executive function 39.
In Japan, waiting patiently before eating is a highly ritualized, daily sociocultural practice. Therefore, Japanese children perform exceptionally well on food-based delay tasks not because of superior inherent willpower, but because their cultural environment has already automated the specific inhibitory response into a low-effort habit 39. Conversely, when the experimental reward is shifted from food to opening a wrapped gift - a practice that is delayed in the United States but not strictly regulated in other cultures - American children exhibit vastly superior inhibition 39.
This research underscores that the expression of executive function is not a universal constant but is profoundly shaped by cultural expectations and environmental routines. Therefore, behavioral interventions, productivity hacks, and self-help methodologies developed exclusively in hyper-individualistic Western contexts often fail when transplanted to collectivistic environments. They ignore the fundamentally different environmental cues, community structures, and social norms that trigger automaticity in diverse populations 39404142.
The 2023-2026 Frontier: Digital Interventions, Mega-Studies, and Neuroplasticity
As the severe limitations of purely cognitive, individualistic self-help become apparent, the scientific community has pivoted dramatically toward massive, data-driven systemic interventions and the direct, empirical tracking of neuroplastic changes in response to behavioral conditioning.
Neuroplasticity and Digital Behavior Change Interventions (DBCIs)
Digital Behavior Change Interventions (DBCIs) - spanning sophisticated mobile applications, virtual reality environments, and wearable biometric technology - are increasingly utilized to foster long-term health habits. Systematic reviews of current DBCIs reveal that the most effective behavioral change techniques (BCTs) rely heavily on automatic prompting, real-time self-monitoring, and granular goal-setting 4344.
More importantly, fMRI and EEG studies evaluating these digital interventions, particularly in populations with developmental disabilities (e.g., ADHD, Autism Spectrum Disorder), demonstrate that sustained engagement with these platforms induces actual structural and functional neuroplasticity 4546. Functional imaging reveals a crucial inverted-U relationship between cognitive load and neuroplasticity during learning 47. Digital environments that dynamically adjust task difficulty to maintain a moderate, optimal cognitive load maximize prefrontal-parietal network activation, thereby significantly accelerating the strengthening of the necessary thalamostriatal synapses required for habituation 47. Furthermore, interventions such as digital cognitive behavioral therapy (CBT) and neurofeedback have been shown to measurably alter brain wave patterns and decrease abnormal functional connectivity in default mode networks, proving that structured digital environments can literally rewire the neural circuitry underlying maladaptive habits 4548.
The Rise of the Mega-Study: Testing Interventions at Scale
To overcome the ongoing replication crisis and the limited statistical power of traditional, small-cohort laboratory experiments, behavioral economists have pioneered the "mega-study" methodology. Rather than testing a single hypothesis on a small group of undergraduate students, mega-studies simultaneously test dozens of distinct, theoretically diverse behavioral interventions on massive, real-world populations 306465.
A watershed mega-study led by Katherine Milkman et al. (2021) involved nearly 700,000 Walmart pharmacy patients to evaluate the efficacy of text-message nudges designed to increase influenza vaccinations. The study rigorously tested 22 different interventions rooted in various behavioral science principles 316465. The results dismantled many theoretical assumptions. The most effective message did not rely on complex habit-stacking, identity-shifting, or extended educational persuasion; it simply informed patients that a vaccine was "reserved for you" or "waiting for you." This specific wording successfully tapped into the psychological endowment effect and implied a default social norm, boosting vaccination rates by up to 11% compared to standard reminder messages 10303165.
However, the mega-study methodology also reveals the humbling, strict limits of behavioral nudges when applied to complex systemic issues. A massive field study involving 140,000 teachers and nearly 3 million students attempted to use behavioral nudges to increase student math proficiency on the Zearn Math platform. Despite testing 15 distinct, expert-designed nudges over several weeks, the average impact on student progress was surprisingly marginal - a mere 1.9% overall increase 49.
These findings solidify the current scientific consensus: while subtle behavioral nudges and carefully designed cues can effectively drive discrete, low-friction, one-off actions (such as getting a scheduled vaccine), they are vastly insufficient for altering complex, longitudinal behaviors (such as mastering mathematics, overcoming obesity, or escaping systemic poverty) without concurrent, massive structural overhauls to the environment 4950.
Conclusion
The scientific discourse surrounding habit formation, behavioral automaticity, and self-regulation has matured far beyond the simplistic, highly marketable heuristics of the commercial self-improvement industry. Human behavior is not merely a product of individual motivation or willpower, nor can complex pathologies be effortlessly hacked through an arbitrary 21-day routine or a mathematically flawed 1% daily compounding rule. It is, instead, the output of a highly complex neurobiological machine - characterized by the precise, dopamine-mediated shifting of control from the prefrontal cortex to the dorsolateral striatum - operating within the strict computational constraints of the rate-correlation theory.
Furthermore, this neurological machinery does not exist in an isolated vacuum; it is constantly subjected to the crushing realities of systemic friction, socioeconomic time poverty, and culturally specific scaffolding. As mega-studies and advanced functional neuroimaging data continually demonstrate, behavioral interventions will consistently fail if they treat the human brain as an isolated variable independent of its environment. True, sustainable behavioral change requires not only a precise understanding of the thalamostriatal synapses that encode our habits, but a concerted, systemic societal effort to alleviate the structural environments that make healthy habituation a metabolic and economic luxury.