Neuroscience of handwriting and keyboarding for learning
Introduction
The widespread transition from handwriting to keyboard typing as the primary mode of text generation represents a significant behavioral shift with profound neurobiological implications. While the QWERTY keyboard has achieved institutional dominance due to its instrumental efficiency in bridging the gap between human thought and machine-readable text, the physical act of writing by hand remains uniquely coupled with neural encoding, memory consolidation, and sensorimotor integration 1. Assessing the distinctions between these modalities requires examining evidence from functional neuroimaging, high-density electroencephalography (EEG), developmental psychology, and extensive educational meta-analyses.
Current research indicates that handwriting is an integrated cognitive-motor skill that recruits a distributed network of cortical and subcortical regions. This process necessitates the continuous integration of lower-level perceptual-motor processes with higher-level executive and mnemonic controls 23. Conversely, typing functions largely as a procedural task characterized by discrete, repetitive bimanual actions, which minimizes production friction but fundamentally alters the way information is processed and retained 14. The mechanisms governing these differences range from the immediate spatiotemporal synchronization of brain waves during the writing act to the long-term structural plasticity of cortical gray matter over the lifespan 56.
Neural Substrates of Handwriting and Typewriting
Cortical Networks in Motor Execution
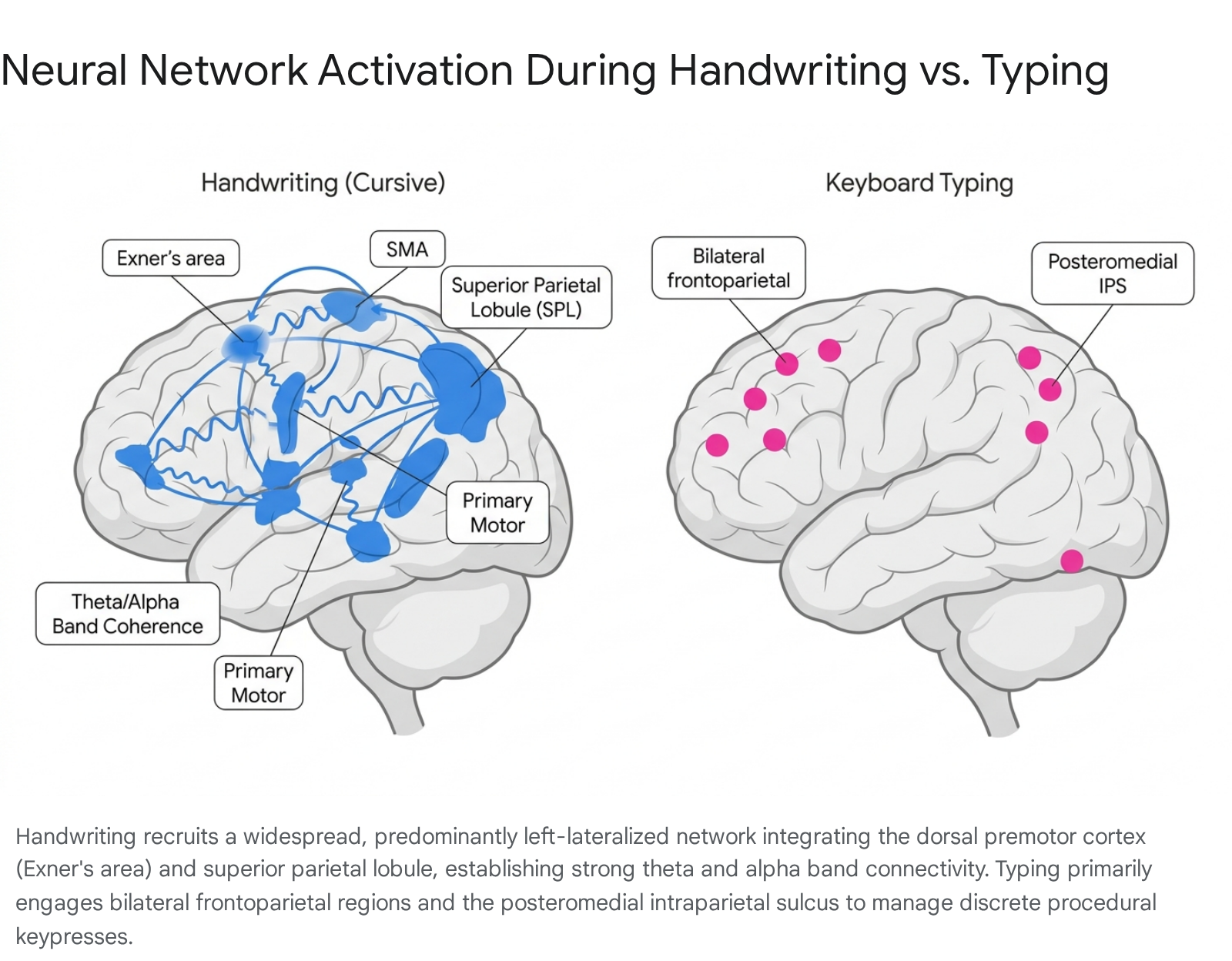
Handwriting is a complex visuospatial and motoric task that demands the coordination of multiple brain regions. Forming letters requires the continuous integration of visual feedback, tactile (haptic) feedback, and precise motor planning 267. Functional magnetic resonance imaging (fMRI) studies consistently identify a left-lateralized cortical network activated during handwriting, regardless of the writer's handedness, owing to the left hemisphere's dominance in language processing 48.
Key nodes in this network include the left dorsal premotor cortex - historically referred to as Exner's area - which is specifically implicated in translating stored orthographic representations into serial motor commands 6910. This region works in concert with the left superior parietal lobule (SPL) and the intraparietal sulcus (IPS), which support visuospatial integration and the formulation of letter-specific motor programs 68. Additionally, handwriting recruits Broca's area (inferior frontal gyrus, pars triangularis) for linguistic processing and phoneme-to-grapheme conversion, alongside the primary motor cortex and supplementary motor area (SMA) 4711. The cerebellum is heavily involved in adjusting movement precision, coordination, and timing during the continuous strokes required for handwriting 412.
By contrast, typewriting relies heavily on bilateral frontoparietal regions to manage spatial processing and attention control, specifically for mapping keys to fingers 46. A distinct region - the left posteromedial intraparietal cortex - shows greater activation during typing than handwriting, reflecting the specific spatial demands of bimanual keyboard interaction 9. Typing engages procedural memory and repetitive finger tapping, demanding significantly less multisensory integration than the continuous manual tracing of letterforms 67. While typing still activates language centers for spelling and lexical retrieval, the execution phase bypasses the intricate sensory-motor integration loops required for shaping physical letters 47.
The following table details the primary neural recruitment profiles for different writing execution modalities identified in fMRI literature.
| Writing Modality | Primary Neural Correlates | Functional Role |
|---|---|---|
| Handwriting (General) | Left dorsal premotor cortex (Exner's area), SPL, SMA, Cerebellum | Translation of graphemes to serial motor commands; complex visuospatial coordination and timing 4810. |
| Keyboard Typing | Posteromedial IPS, bilateral frontoparietal network, SMA | Bimanual spatial mapping; discrete procedural keypresses and attention shifts 49. |
| Stylus on Touchscreen | Primary motor cortex, supplementary motor area, visuospatial networks | High-vigor motor execution; reinforces embodied sensorimotor coupling similar to paper handwriting but with distinct kinematic friction 1314. |
Brain Connectivity and Oscillatory Dynamics
Beyond the localization of brain activity, the temporal synchronization of neural networks differs substantially between handwriting and typing. High-density EEG studies measuring oscillatory brain activity provide evidence that the physical act of handwriting primes the brain for learning by synchronizing specific frequency bands critical for memory formation.
Research analyzing university students using 256-sensor EEG nets found that writing with a digital pen induces widespread and elaborate connectivity patterns in the theta (3.5 - 7.5 Hz) and alpha (8 - 12.5 Hz) bands 515. This increased coherence was localized primarily to the parietal and central brain regions (specifically the parietal midline, central left, and central right regions) 5. These frequency bands and connectivity networks are robustly associated with sensorimotor integration, long-term potentiation in the hippocampus, and the encoding of new information 516. The complex kinesthetic feedback required to shape individual letters appears to drive this widespread functional connectivity 515.
Conversely, typewriting - which involves identical, simple motor movements (striking a key with a single finger) regardless of the specific letter being produced - does not elicit these widespread theta and alpha connectivity patterns 515. The minimal variation in proprioceptive feedback during typing leads to narrower neural activation and shallower cognitive engagement. This suggests that the inherent "production friction" of handwriting - the slower, deliberate movement required to form shapes - actively facilitates memory encoding by forcing the brain to dedicate more cortical resources to the task 11617.
Sensory Feedback and Visuospatial Anchoring
The tactile medium of writing also significantly influences cognitive encoding. Analog physical media, such as paper notebooks, provide haptic feedback and spatial permanence that digital screens currently struggle to replicate. Deficits in proprioception and tactile perception correlate heavily with handwriting difficulties and somatodyspraxia in children, underscoring the reliance on this sensory input for effective graphomotor movement 12.
Research exploring the differences between scheduling on physical paper datebooks versus digital tablets or smartphones demonstrates that paper provides unique, complex spatial and tactile information that enhances memory recall . In an fMRI study involving university students, writing on physical paper led to significantly greater activation in the hippocampus, precuneus, and language-associated areas compared to using digital tools 18. The hippocampus relies on spatial cues for navigation and memory retrieval; physical paper offers tangible permanence, irregular strokes, and fixed spatial locations (e.g., visualizing a note on the bottom-left corner of a specific page) . Digital environments, characterized by uniform scrolling, standardization, and a lack of fixed physical boundaries, fail to provide the brain with comparable spatial landmarks, resulting in faster but shallower cognitive processing 19.
Orthographic Depth and Script Variations
The neural substrates of handwriting vary detectably depending on the orthographic depth, morphological complexity, and physical constraints of the script being produced. The brain adapts its functional architecture to accommodate the specific processing demands of different writing systems.
Cursive Versus Block Print
Within alphabetic scripts, block-letter printing requires discrete motor planning, as the pen is lifted between each character. Cursive handwriting, however, is a continuous motor activity that necessitates smooth transitions between letters, requiring the writer to anticipate the subsequent character in an uninterrupted motion 46.
Neuroimaging demonstrates that this fluidity results in greater recruitment of the primary motor cortex and cerebellar regions compared to block printing 4620. An fMRI study comparing the perception of handwritten cursive letters versus standard printed letters revealed that viewing cursive letters induced stronger activation in the left primary motor cortex and the supplementary motor area 20. This suggests that the visual perception of cursive is highly embodied; the brain covertly simulates the action sequence required to produce the continuous script in order to recognize it 1020. At the whole-brain level, perceiving cursive also activates additional posterior regions, including the right superior frontal gyrus, middle occipital gyrus, parahippocampal gyri, and left fusiform gyrus 20.
Logographic Versus Syllabic Processing: Kanji and Kana
Studies examining Japanese orthography - which utilizes both the syllabic Kana script and the logographic Kanji script - illustrate how the brain processes differing levels of orthographic depth. Kana is a phonemic system with a direct grapheme-to-phoneme correspondence, similar to alphabetic languages. Kanji is a morphogrammatic (logographic) system where characters represent entire semantic concepts, functioning similarly to Chinese characters 2122.
Neuroimaging data reveal a clear dissociation in the neural pathways utilized for these scripts. Reading and writing Kana relies primarily on a dorsal processing stream, linking the occipital cortex to the inferior parietal area, and uniquely activates the left inferior parietal lobule (BA 40) during conflict detection tasks like the Stroop test 2223. This pathway specializes in phonological recoding - translating symbols to sounds before accessing meaning 24.
Conversely, Kanji processing heavily recruits the ventral visual stream, connecting the occipital lobe to the temporal cortex 22. The execution of Kanji writing elicits strong activation in the bilateral fusiform gyri, the left inferior frontal gyrus (BA 9), and right-hemisphere occipito-temporal areas (such as BA 37 and BA 39) dedicated to complex visuospatial processing 22232425. The brain processes logographs directly for meaning, relying heavily on visual word-form analysis and visuospatial attention networks rather than phonological conversion 212224.
| Japanese Script | Processing Stream | Primary Brain Activations | Functional Mechanism |
|---|---|---|---|
| Kana (Syllabic) | Dorsal Stream | Left inferior parietal lobule (BA 40), superior posterior temporal gyrus | Phonological recoding; mapping graphemes to phonemes 212223. |
| Kanji (Logographic) | Ventral Stream | Bilateral fusiform gyri, left IFG (BA 9), right occipito-temporal (BA 37, 39) | Visuospatial processing; direct mapping of morphograms to semantic concepts 232425. |
Developmental Impacts on Literacy and Cognition
Neural Pathway Formation in Early Childhood
The acquisition of literacy in early childhood is inextricably linked to the development of fine motor skills. In preliterate children, the act of handwriting facilitates the functional specialization of the brain's "reading circuit" in ways that typing and passive observation do not.
Neuroimaging studies comparing 5-year-old children who actively wrote, typed, or traced letters found that only the children who hand-printed the letters exhibited subsequent robust activation in the left fusiform gyrus during visual letter perception 2826. This region is critical for visual word recognition and spatial processing. This finding suggests that the perceptual networks required for reading are fundamentally driven by the motor execution of forming letters, rather than mere visual feedback 26272829. The process of self-generating a character by hand creates a highly robust internal representation of the letter, anchoring visual shapes to physical movements, which accelerates letter recognition and later reading fluency 28263031.
Subsequent studies assessing 7-year-old children learning cursive writing - either by writing the letters themselves or passively observing an adult write them - confirmed that active motor training led to increased recruitment of the sensori-motor network associated with letter perception, alongside activation in the insula and claustrum 272829. Passive observation yielded no such functional specialization, indicating that motor execution is a non-negotiable component of establishing these neural networks 27.
Interventions for Neurodiverse Learners
For neurodiverse children - including those with dysgraphia, specific learning disabilities, or delayed fine motor control - handwriting serves as a critical developmental and therapeutic tool. While the physical act of writing can be exceptionally challenging due to deficits in motor planning, executive function, and sensory processing, engaging in handwriting aids in visual-motor integration and emotional regulation 3233.
Recent advancements in haptic guidance systems demonstrate that supplementing handwriting practice with targeted tactile feedback can overcome cognitive and fine motor delays. Longitudinal experimental studies show that providing naturalistic haptic feedback along the trajectory of a handwriting task significantly improves motor functions in children with delayed pincer grasp control and intellectual difficulties 3438. These interventions leverage activity-dependent neural plasticity, wherein active engagement with individually tailored sensorimotor tasks drives adaptive cortical changes, improving not only the trained handwriting tasks but generalizing to untrained visual-motor tasks 123435.
Note-Taking Modalities and Academic Achievement
The debate over the optimal note-taking modality in higher education has generated substantial empirical scrutiny over the past decade. The controversy primarily centers on the trade-off between transcription volume (the advantage of typing) and encoding depth (the advantage of handwriting).
The Encoding Hypothesis and Verbatim Transcription
The modern debate was heavily influenced by the 2014 study by Mueller and Oppenheimer, which established the "encoding hypothesis." This hypothesis posited that laptop note-takers tend to act as stenographers, transcribing lectures verbatim because typing allows for rapid, low-friction text generation. Longhand note-takers, constrained by slower writing speeds, are forced to actively process, synthesize, and reframe the information in their own words 333637.
Their initial experiments reported that while typing yielded a significantly higher volume of notes and greater verbatim overlap, longhand writers significantly outperformed typists on conceptual application questions 333843. The original study reported an exceptionally large effect size for factual recall in the initial experiment (Cohen's d = 0.97), prompting widespread laptop bans in university classrooms and widespread media coverage 39.
Replication Outcomes and Methodological Nuance
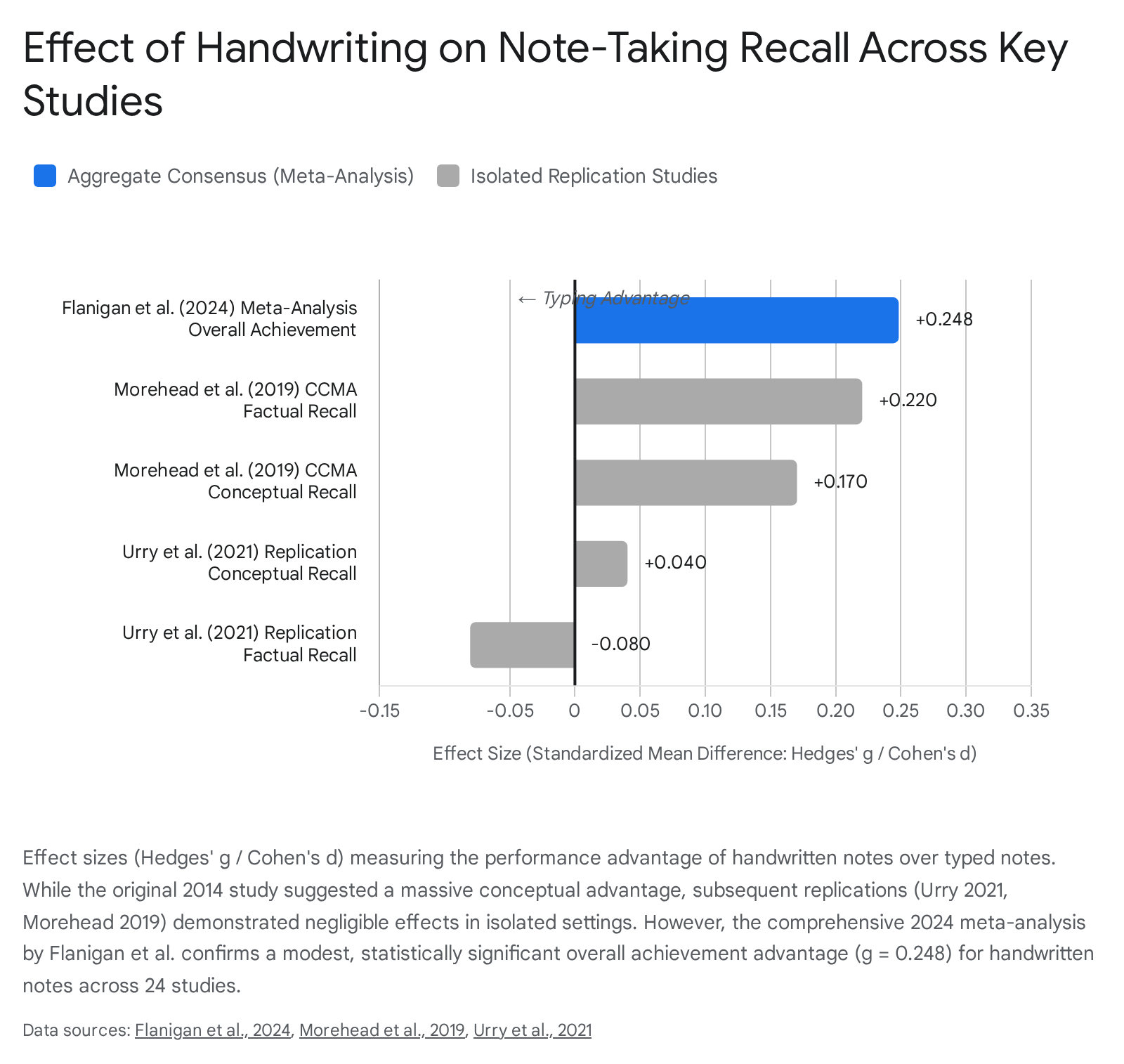
However, subsequent attempts to directly replicate these findings have yielded a highly nuanced picture, initiating a reassessment of the magnitude of the handwriting advantage. Rigorous direct replications by Urry et al. (2021) and Morehead et al. (2019) failed to reproduce the large conceptual advantages for handwriting when variables such as prior knowledge and test delay were controlled 1394041.
For instance, Urry et al. (2021), using the exact materials from the original study with 145 students, found negligible modality effects for immediate recall when students were not permitted to review their notes (Hedges' g = -0.08 for factual recall and g = 0.04 for conceptual recall) 14142. Similarly, Morehead et al. (2019) found that laptop users continued to produce significantly more words (Cohen's d = -0.58), but the advantage for longhand notes on immediate factual recall was small (d = 0.27) and not statistically significant for conceptual recall 40.
The use of specific statistical effect size metrics is critical in interpreting this literature. Cohen's d represents the standardized mean difference between two groups, but it exhibits an upward bias in small sample sizes (typically below 20-50 per group). Meta-analysts prefer Hedges' g, which applies a correction factor to remove this small-sample bias, preventing small studies from inflating pooled estimates 48435044. When early, small-sample studies are corrected using Hedges' g, the initially massive effect sizes surrounding the longhand advantage attenuate significantly 3944.
Meta-Analytic Synthesis of Educational Outcomes
Despite the attenuation of the original extreme effect sizes in highly controlled laboratory settings, broad meta-analyses confirm a consistent, reliable achievement advantage for handwriting over typing in ecologically valid classroom settings.
A 2024 meta-analysis by Flanigan et al. encompassing 24 studies and over 3,000 college students concluded that taking and reviewing handwritten notes leads to higher overall course achievement (Hedges' g = 0.248, $p < 0.001$) 454647. Crucially, the meta-analysis confirmed the volume disparity: typists generate a vastly larger volume of notes (Hedges' g = 0.919, $p < 0.001$) 4547.
The primary moderating variable appears to be the opportunity for review. The advantage of handwritten notes is stabilized and amplified when students are allowed to study their notes prior to the final assessment 384046. When immediate tests are administered without a review period, the high volume of typed notes occasionally offsets the shallower encoding 140. However, for long-term retention, the synthesis of these findings suggests that while typing is an unmatched instrument for rapid data capture, handwriting forces an initial layer of cognitive synthesis that results in superior memory consolidation.
The table below summarizes the aggregate effect sizes from prominent meta-analyses and replication syntheses in the domain of note-taking.
| Meta-Analysis / Synthesis | Sample Scope | Primary Metric Assessed | Reported Effect Size | Clinical Interpretation |
|---|---|---|---|---|
| Flanigan et al. (2024) | 24 studies (N=3,005) | Overall Academic Achievement | Hedges' g = 0.248 | Modest but highly significant advantage for handwritten notes 4547. |
| Flanigan et al. (2024) | 24 studies (N=3,005) | Note-Taking Volume (Word Count) | Hedges' g = -0.919 | Massive volume advantage for keyboard typing 4547. |
| Urry et al. (2021) | Direct Replication | Conceptual Recall (Immediate) | Hedges' g = 0.04 | Negligible difference without note review 141. |
| Morehead et al. (2019) | CCMA of Replications | Factual Recall | Cohen's d = 0.22 | Small, non-significant encoding benefit for longhand 40. |
| Allen et al. (2020) | 14 studies (N=3,075) | Measured Educational Outcomes | r = -0.142 | Electronic device use reduced measured outcomes 48. |
(Note: Negative volume/outcome effect sizes traditionally denote an advantage for the laptop/typing condition depending on the coding schema of the specific meta-analysis).
Structural Neuroplasticity and Cognitive Aging
The neurological benefits of handwriting are not limited to childhood development and academic performance; they extend across the lifespan, influencing structural brain plasticity and acting as a biomarker for cognitive health in aging populations.
Volumetric Changes in Gray and White Matter
Prolonged engagement in cognitively and motorically demanding tasks, such as handwriting, contributes to the maintenance of brain structure and cognitive reserve 4950. Advanced neuroimaging techniques, particularly Voxel-Based Morphometry (VBM) and Diffusion Tensor Imaging (DTI), track these macro and microstructural changes. Normal aging is associated with a progressive decline in fine motor precision, sensory feedback, and cognitive slowing 51. VBM analyses comparing young and older cohorts reveal widespread age-related reductions in gray matter (GM) volume in the frontal, insular, and cingulate cortices, alongside white matter (WM) degradation prominently in the thalamic radiations 52. Field strength dependencies (e.g., 1.5 T vs 3 T scanners) also show significant age-related differences in cerebellar and visual-associated GM volumes 53.
Maintaining the complex sensorimotor integration required by handwriting can promote adaptive neural changes. MRI data reveals that the brain is capable of rapid structural plasticity. For instance, executing repetitive fine-motor tasks (such as a finger-tapping task) can induce measurable fluctuations in estimated GM volume within the motor cortex, superior occipital lateral cortex, thalamus, and insula in a matter of minutes 54. Regularly utilizing handwriting, rather than exclusively relying on digital keyboards, provides continuous, complex stimulation to these fine-motor and visuospatial pathways. This active engagement aligns with theories of neuroplasticity suggesting that sustained activity in functional networks can counteract age-related cortical thinning and cognitive decline 49505162. Furthermore, in stroke recovery populations, DTI metrics show that higher Fractional Anisotropy (FA) and lower Mean Diffusivity (MD) in regions like the globus pallidus and caudate nucleus correlate significantly with better motor function recovery, highlighting the importance of preserving white matter integrity through continued motor engagement 6.
Handwriting Kinematics as Biomarkers
Because handwriting relies heavily on the precise, uninterrupted communication between the cortex, basal ganglia, and cerebellum, it serves as a highly sensitive, non-invasive biomarker for neurological integrity. Neurodegenerative conditions - such as Parkinson's disease, Alzheimer's disease, and Multiple Sclerosis - often manifest in subtle handwriting kinematics long before overt clinical symptoms appear 25155.
In Parkinson's disease, the progressive disruption of the extrapyramidal tracts and basal ganglia alters the scaling and timing of fine motor movements. This frequently presents as micrographia (abnormally small handwriting), altered pen pressure, and disrupted stroke fluidity 255. High-resolution digital kinematic analyses of handwriting are now being utilized for early diagnosis, the longitudinal monitoring of disease progression, and the development of personalized rehabilitation interventions, bridging the gap between behavioral neuroscience, aging research, and digital medicine 5155.
The Digital Native Context
The generational shift in writing modalities has introduced the concept of "Digital Natives" - individuals born into pervasive digital environments - versus "Digital Immigrants." This divide has practical implications for cognitive load, digital stress, and the evolution of motor habits.
Kinematics and Cognitive Load in Smartphone Typing
While traditional keyboard typing requires bimanual coordination, modern digital interaction is heavily skewed toward touchscreen smartphone typing, primarily utilizing the thumbs. Kinematic analyses of touchscreen typing reveal that digital natives exhibit a higher-vigor motor execution profile characterized by faster general movements and later peak deceleration compared to older cohorts 14. Bimanual advantages and bigram frequency effects known to influence traditional QWERTY typing generalize to mobile typing when using two thumbs, indicating shared underlying motor processes despite the interface differences 56.
However, the rapid nature of this digital input has cognitive consequences. Systematic reviews indicate that while digital natives possess higher digital confidence and fluency, they are highly susceptible to social media fatigue, social comparison anxiety, and cognitive overload stemming from the continuous, high-speed influx of fragmented digital information 1357. The rapid scrolling and typing behaviors characteristic of smartphone use foster embodied fluency in digital actions but are associated with shallower reading comprehension and more fragile memory retention compared to traditional print reading and handwritten engagement 19. This suggests a cognitive trade-off: digital natives achieve highly optimized, low-friction motor execution at the expense of the deep encoding and sustained attention fostered by the friction of traditional handwriting 141950.
Conclusion
The convergence of functional neuroimaging, high-density EEG, and rigorous educational meta-analyses demonstrates a clear bifurcation in the utility of handwriting versus typewriting. Typing operates primarily as an instrumental tool for efficient output, relying on procedural memory and bilateral frontoparietal networks to minimize cognitive load and maximize transcription volume. It excels in environments where the rapid capture of verbatim text is the primary objective.
In contrast, writing by hand is a highly embodied, cognitively demanding process that requires continuous sensorimotor integration. By forcing the brain to execute precise serial motor commands, process haptic feedback, and navigate physical spatial constraints, handwriting activates a widespread network encompassing the dorsal premotor cortex, superior parietal lobule, and memory-critical regions. This activation results in synchronized oscillatory connectivity in the theta and alpha bands, creating a neurobiological environment optimized for encoding, synthesizing, and retaining complex information. While keyboards will inevitably maintain their dominance in digital production, handwriting remains an essential biological mechanism for early literacy development, deep conceptual learning, and the preservation of structural neuroplasticity and cognitive resilience across the human lifespan.