Neuroscience of face processing and prosopagnosia
Cortical Architecture of Face Perception
The recognition of individual faces represents one of the most computationally demanding tasks executed by the primate visual system. Given the evolutionary imperative to distinguish among conspecifics, interpret complex emotional states, and evaluate social hierarchies, the human brain has evolved a highly specialized and localized neural architecture dedicated to face perception. The foundational understanding of this localized modularity emerged from the functional region of interest (fROI) approach, which successfully pinpointed the exact functional neuroanatomy of face processing within the human neocortex 12. This discovery, later paralleled by the identification of analogous face patches in macaque monkeys via single-unit recordings, demonstrated that face recognition is supported by dedicated, modular circuitry rather than generalized visual processing systems 13.
Face processing is executed through a distributed network of functionally distinct but highly interconnected cortical regions. This network is structurally divided into a "core" system responsible for the immediate visual analysis of facial features, and an "extended" system that links these perceptual representations to semantic knowledge, emotional resonance, and biographical memory 456.
The Core Face Network
The core face processing network is located primarily within the ventral occipitotemporal cortex (VOTC). Activity within this network relies on feed-forward effective connectivity projecting from the primary and secondary visual cortices. While bilateral activation is routinely observed during face viewing tasks, the network exhibits a fundamental right-hemisphere dominance, with face-selective responses and corresponding deficits localized more strongly to the right hemisphere 478. The core network comprises three principal regions that process facial stimuli sequentially.
The Occipital Face Area (OFA), situated in the lateral inferior occipital gyrus, acts as the earliest cortical stage of face processing within the core network. The OFA is responsible for the part-based, structural analysis of faces, encoding individual facial features - such as the eyes, nose, and mouth - prior to holistic integration 4910. Functional neuroimaging indicates that the OFA provides critical structural information necessary for subsequent perceptual processing in downstream regions. Lesions localized to the OFA disrupt this feed-forward cascade, precipitating profound deficits in basic face perception 11.
Information regarding invariant facial features flows from the OFA to the Fusiform Face Area (FFA), located in the lateral middle fusiform gyrus (with primary peak activation coordinates frequently observed around MNI: 42, -50, -19) 59. The FFA integrates discrete structural parts into a holistic, configural representation, a computational step essential for establishing unique identity. The FFA exhibits high face selectivity compared to non-face visual object categories and responds automatically upon viewing a face 19. Within the FFA, researchers have identified distinct neural patterns corresponding to individual identities, suggesting that this region maintains a complex, multidimensional representational "face space" 12.
While the OFA and FFA compute the invariant aspects of facial identity, the posterior Superior Temporal Sulcus (pSTS) is functionally specialized for processing dynamic, changeable facial features. The pSTS monitors transient movements in eye gaze, mouth articulation, and shifting facial expressions 469. Consequently, the pSTS shows significantly stronger blood-oxygen-level-dependent (BOLD) responses to dynamic, moving faces compared to static imagery 13.
| Core Region | Anatomical Location | Primary Computational Function |
|---|---|---|
| Occipital Face Area (OFA) | Lateral inferior occipital gyrus | Structural encoding; part-based analysis of distinct facial features; provides feed-forward input. |
| Fusiform Face Area (FFA) | Lateral middle fusiform gyrus | Holistic integration of features; processing of invariant properties to establish and maintain identity. |
| Posterior Superior Temporal Sulcus (pSTS) | Posterior superior temporal sulcus | Dynamic tracking; analysis of changeable aspects including eye gaze, expressions, and articulation. |
The Extended Face Network
The core network projects to the extended face network, which comprises the ventral anterior temporal lobe (vATL), the anterior temporal cortex (aTC), the inferior frontal gyrus, the amygdala, and the precuneus 451414. The FFA projects directly to the anterior middle temporal gyrus (aMTG) and anterior inferior temporal gyrus (aITG), which manage the retrieval of biographical and semantic information associated with known identities 414. Damage to the anterior temporal lobe preserves basic visual face perception but severs the connection to identity, preventing the recognition of highly familiar individuals.
Early psychological models theorized that the processing of identity (via the FFA) and expression (via the pSTS) operated in strictly parallel, independent streams. However, recent functional connectivity analyses demonstrate high interactivity. When observers view sequences of faces where the expression and gaze change but the underlying identity remains constant, functional connectivity between the pSTS and the FFA significantly increases 15. This functional coupling indicates that distinct neural pathways interact to process changeable features in a socially meaningful, identity-specific context 1315.
Subcortical Pathways and Rapid Peripheral Detection
While the cortical core network is responsible for the high-resolution discrimination required for individual identity recognition, recent electrophysiological evidence reveals that basic face detection occurs significantly earlier via an evolutionarily ancient subcortical pathway. The ability to rapidly detect a face - even in the low-acuity periphery of the visual field - is a critical survival mechanism for orienting attention prior to resource-intensive cortical processing.
A 2024 study mapped a rapid face-detection circuit routed through the midbrain superior colliculus (SC), a structure traditionally associated with eye movements and spatial orienting 161718. Visual signals bypass the slower cortical hierarchy, routing from the retina through the lateral geniculate nucleus (LGN) directly to the superior colliculus 1619. Visual neurons within the SC exhibit a powerful preference for face images, with this preference emerging within 40 to 50 milliseconds of stimulus onset 161819.
This early subcortical detection precedes activation in the cortical face patches of the VOTC, which typically occurs at latencies of 150 to 170 milliseconds 1819. At the population level, SC activity can distinguish faces from other visual objects with 80% to 92% accuracy within this ultra-short latency window, whereas discrimination of non-face objects requires approximately 65 to 100 milliseconds 161719. This subcortical shortcut functions to detect face-like stimuli in the periphery and rapidly trigger orienting saccades, bringing the face into the fovea where the high-resolution cortical networks can extract detailed identity and expression data 1820.
Experience-Dependent Specialization and the Other-Race Effect
The Other-Race Effect (ORE) is a consistently observed psychological phenomenon wherein individuals exhibit poorer recognition memory and decoding accuracy for faces belonging to racial or ethnic groups different from their own 1221. The ORE provides a primary model for understanding experience-dependent neural specialization and perceptual narrowing in the developing human brain.
Perceptual Narrowing in Infancy
During the first few months of life, the infant visual system is broadly tuned, allowing accurate discrimination between faces of any race. However, as infants approach 9 to 12 months of age, their face processing abilities undergo perceptual narrowing. Continuous exposure to predominantly own-race faces tunes the visual system to maximize sensitivity to the specific morphological variations present in their immediate environment, simultaneously reducing sensitivity to the structural variations characteristic of less frequently encountered other-race faces 2122.
The trajectory of the ORE is highly dependent on early visual and sociocultural experiences. Interventions utilizing multisensory, intersensory redundancy demonstrate the plasticity of this mechanism. When 12-month-old infants are familiarized with dynamic, audiovisual presentations of other-race faces, their ability to individuate those faces is preserved, indicating that enriched sensory input can mitigate the typical onset of the ORE 22.
Neural and Representational Signatures
In adults, the ORE is measurable through distinct electrophysiological and neural representational signatures. High-resolution EEG and fMRI decoding studies identify a reliable neural counterpart to the ORE: multivariate decoding accuracy is significantly reduced when processing other-race faces compared to same-race faces 12. Neural divergence begins early in the visual processing stream, with high degrees of cultural difference manifesting in primary visual cortex (V1) and OFA activity within the first 200 milliseconds of perception, reflected in differential P100, N170, and P200 event-related potential (ERP) amplitudes 2123.
Under classical theoretical models, faces are encoded as vectors within a multidimensional "face space." Extensive experience optimizes the dimensions of this space to differentiate same-race faces effectively. Because the system lacks tuned dimensions for unfamiliar morphological traits, other-race faces are densely clustered in a compressed region of this representational space. Data-driven image reconstruction from EEG patterns reveals that the brain systematically distorts the visual representation of other-race faces, encoding them with less distinct identity information. Consequently, other-race faces are frequently perceived as more "typical" or average, as well as physically younger and more expressive than their true morphological structure dictates 1224.
The ORE is increasingly conceptualized as a form of perceptual expertise. Humans process own-race faces at a highly individuated, subordinate level - akin to how an expert views a specialized domain - while processing other-race faces at a categorical, basic level 2526. Immersive exposure to an other-race culture, which provides both perceptual richness and the social motivation to individuate, can shift this processing dynamic, improving other-race face recognition and modifying the underlying neural tuning biases 2526.
| Processing Metric | Same-Race Face Processing | Other-Race Face Processing |
|---|---|---|
| Cognitive Level | Subordinate level (Individuated identity). | Basic level (Categorical classification). |
| Representational Space | Widely distributed across optimized dimensions. | Compressed and densely clustered. |
| Neural Decoding | High multivariate decoding accuracy. | Significantly reduced decoding accuracy. |
| Visual Reconstruction | Accurate extraction of distinct features. | Biased toward average, younger, or more expressive representations. |
Artificial Models of Face Processing
Deep Convolutional Neural Networks (DCNNs) have emerged as powerful computational models for understanding the human visual system. Historically, behavioral phenomena such as the Face Inversion Effect (a severe drop in recognition accuracy when faces are presented upside down) and the Other-Race Effect were cited as evidence that face recognition relied on exclusive, biologically innate mechanisms 27.
Recent computational research demonstrates that these phenomena are not biologically exclusive, but rather mathematically optimal solutions that emerge naturally when a system is tasked with high-accuracy, subordinate-level discrimination. DCNNs optimized strictly for face identification spontaneously develop a characteristic representational space and exhibit human-like behavioral signatures, including the Face Inversion Effect. Crucially, CNNs trained solely on general object categorization, or trained simply to detect the presence of a face without individuating identity, do not develop these signatures 2728. Similarly, when CNNs are trained on demographically skewed datasets (e.g., exclusively Asian or exclusively White faces), they develop experience-dependent deficits that mirror the human Other-Race Effect 29.
Furthermore, training regimens suggest that functional modularity is an optimal outcome of multi-task learning. "Dual-task" CNNs trained simultaneously on face identification and object categorization spontaneously develop functionally segregated processing streams, echoing the modularity of the human ventral visual pathway. This dual optimization provides a computational explanation for visual phenomena such as face pareidolia (seeing faces in inanimate objects); only CNNs trained for both object categorization and face identification rely on human-like facial features (e.g., eyes and mouths) to classify pareidolia stimuli 3031.
Despite these structural convergences, deep learning models possess limitations in replicating human neural dynamics. While DCNN activations correlate strongly with human behavioral output during static image processing, they fail to adequately model the human brain's response to dynamic, moving faces. fMRI analyses reveal that artificial neural codes extracted from DCNNs correlate weakly with human brain activity when participants view naturalistic videos of faces in motion 32. The unique information encoded in the human brain relates to dynamic temporal integration, working memory, and social attention - complexities that current feed-forward, static-image-trained DCNN architectures cannot fully emulate 3233.
Pathology of Recognition: Prosopagnosia
Prosopagnosia constitutes a severe, selective deficit in the ability to recognize facial identity, occurring in the absence of generalized visual impairment, cognitive decline, or widespread memory dysfunction 34. The study of prosopagnosia isolates the specific neuroanatomical and electrophysiological substrates necessary for face perception. The condition is categorized into acquired and developmental etiologies.
Acquired Prosopagnosia
Acquired Prosopagnosia (AP) occurs when an individual loses a previously intact face recognition system due to focal brain injury, such as ischemic stroke, hemorrhage, trauma, or neurodegenerative disease affecting the posterior cortex 3435. Lesion network mapping of AP patients demonstrates that causal lesions consistently map to a functionally connected brain network intrinsically tied to the right FFA and left frontal regions (including the anterior prefrontal cortex, anterior cingulate cortex, and middle frontal gyrus) 36.
AP exhibits substantial clinical heterogeneity and is broadly divided into two neurocognitive subtypes:
- Apperceptive Prosopagnosia: This variant involves a fundamental failure in face perception and the encoding of facial structure. Patients struggle to match simultaneous unfamiliar faces or discern basic structural differences. Anatomically, apperceptive AP is strongly associated with occipitotemporal lesions, often involving the OFA and FFA bilaterally or in the right hemisphere. The feed-forward construction of the face percept is interrupted 343837.
- Associative Prosopagnosia: In this variant, patients accurately perceive and match faces, indicating intact structural encoding of the face percept. However, they fail to associate this percept with stored biographical memory or identity. Associative AP is primarily linked to lesions in the anterior temporal lobe and parahippocampal regions. The core network functions normally, but the structural connection to the extended semantic network is severed 343837.
Individuals with AP frequently exhibit highly specific neurological comorbidities due to the proximity of related processing areas. Cerebral dyschromatopsia (color blindness due to cortical damage) frequently co-occurs with apperceptive AP due to adjacent damage in the lingual and fusiform gyri 1434. Topographical disorientation - an inability to navigate familiar environments - is reported in nearly 30% of AP cases, corresponding to collateral damage in the parahippocampal place area 1434.
| AP Subtype | Cognitive Locus of Deficit | Behavioral Marker | Typical Lesion Location |
|---|---|---|---|
| Apperceptive | Failure to accurately perceive or encode structural facial configurations. | Fails tests of basic face matching and structural discrimination. | Occipitotemporal cortex (OFA/FFA); often bilateral or right unilateral. |
| Associative | Intact perception; failure to link face percept to stored memory or identity. | Passes face matching tests but fails famous/familiar face recognition. | Anterior temporal cortex; parahippocampal regions. |
Developmental Prosopagnosia
Developmental Prosopagnosia (DP), or congenital prosopagnosia, refers to a lifelong deficit in face recognition that manifests in early childhood without any history of macroscopic brain lesions, neurological trauma, or low-level visual acuity limitations 3839. Epidemiological estimates suggest that DP affects between 2% and 2.5% of the general population 343840. DP is highly heritable and is hypothesized to arise from neurodevelopmental anomalies affecting neural migration or connectivity during early brain maturation 741. Certain genetic markers, including mutations in the forkhead box G1 gene, have been identified in specific developmental cases 35.
Unlike the overt focal lesions seen in AP, the structural correlates of DP are nuanced. High-resolution MRI analyses reveal reduced gray matter density and volume in the temporal lobes - specifically the pSTS, middle temporal gyrus (MTG), and fusiform gyrus - alongside decreased white matter fractional anisotropy and connectivity within the core face network near the right FFA 47.
Functionally, DP patients exhibit abnormal task-induced brain activity. Feed-forward effective connectivity from early visual cortices into the core face network is severely attenuated, indicating a breakdown in network integration rather than the complete destruction of a specific node 47. Electrophysiologically, many DPs demonstrate an abnormal N170 ERP, which frequently lacks the typical right-hemisphere lateralization and face-selectivity observed in neurotypical individuals 47. Behaviorally, due to compromised holistic processing capabilities, individuals with DP often rely on piecemeal, feature-based compensatory strategies, navigating social interactions by identifying isolated traits such as a specific hairline, mole, or gait 73944.
| Feature | Acquired Prosopagnosia (AP) | Developmental Prosopagnosia (DP) |
|---|---|---|
| Etiology | Sudden onset following focal brain damage (stroke, trauma, tumor, degeneration). | Lifelong condition; genetic or early neurodevelopmental origins; no macroscopic lesion. |
| Neural Substrates | Macroscopic focal lesions obliterating specific nodes (e.g., FFA, OFA, ATL). | Reduced gray matter volume and compromised white matter integrity. |
| Network Pathology | Localized structural destruction. | Disrupted feed-forward functional connectivity between otherwise intact anatomical regions. |
Prosopagnosia and Autism Spectrum Disorder
Face recognition deficits are frequently observed among individuals with Autism Spectrum Disorder (ASD). Recent rigorous assessments have revealed an extraordinarily high rate of co-occurrence: over 36% of autistic adults without intellectual disability meet the strict clinical diagnostic criteria for prosopagnosia, compared to roughly 2% in the neurotypical population 45424743.
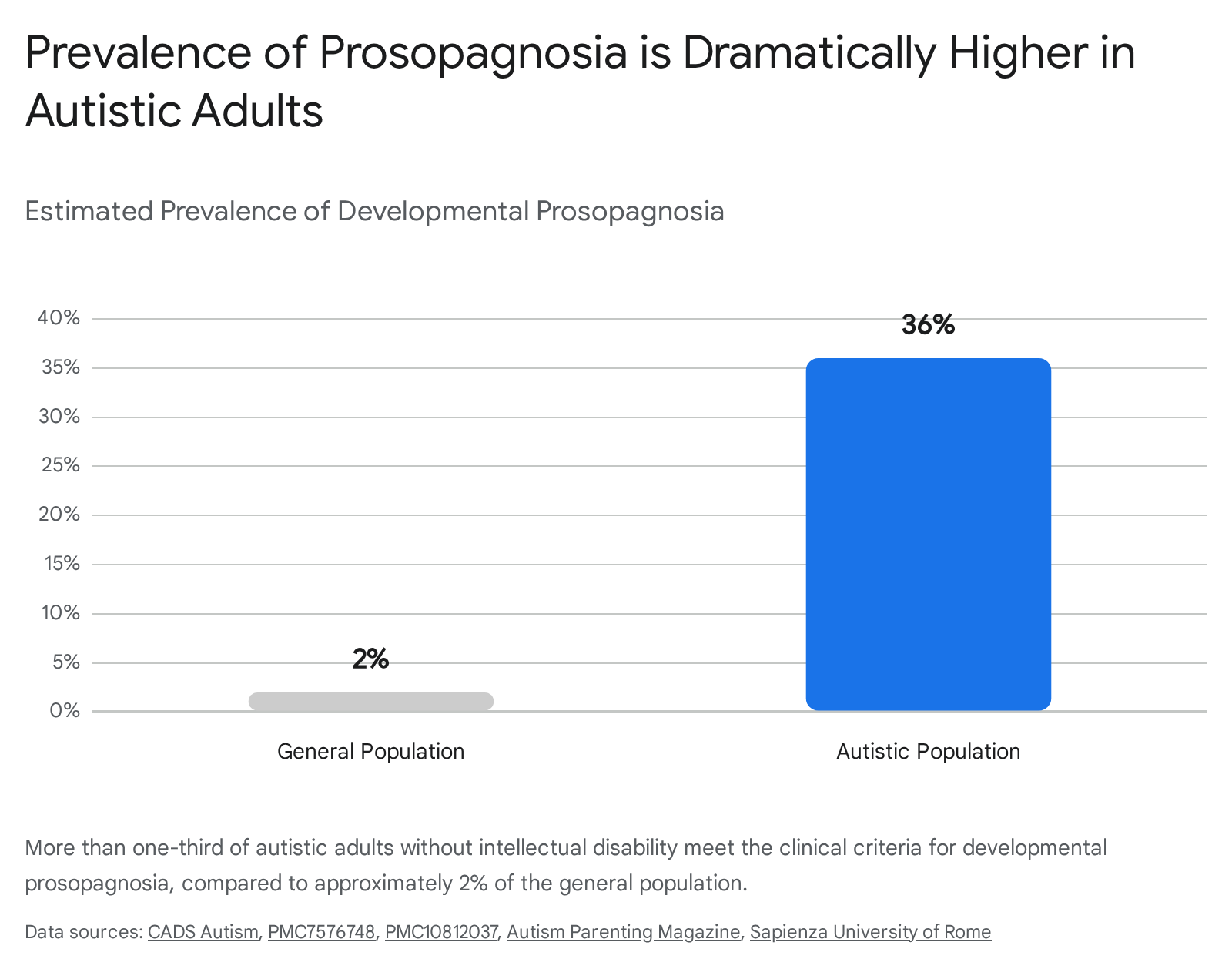
Historically, it was hypothesized that poor face processing in ASD was a secondary, behavioral consequence of reduced social motivation - specifically, an avoidance of eye contact that prevented the acquisition of perceptual expertise with faces. However, emerging cognitive neuroscience evidence points toward foundational, structurally distinct neurobiological deficits. Neuroimaging studies show that autistic individuals exhibit reduced functional connectivity between the FFA and frontal cortices, presenting a network underconnectivity profile strikingly similar to that seen in classic DP 47. Furthermore, hypothesized deficits in the rapid subcortical face-detection pathway (the superior colliculus) may prevent autistic infants from involuntarily orienting toward faces during critical developmental windows, fundamentally altering the trajectory of downstream cortical specialization 171820.
Crucially, researchers have identified a clear dissociation between pure perceptual face memory and broader socio-emotional cognitive skills. Autistic individuals with prosopagnosia do not differ from autistic individuals without prosopagnosia in overall autism symptom severity (as measured by the ADOS or AQ), general intelligence, empathy measures, or levels of alexithymia 404243. Face identity recognition is instead linked specifically to mental state recognition from the eye region (measured via the RMET) 4243. This dissociation indicates that the face identity recognition deficit in ASD represents an independent, potentially genetic endophenotype, rather than merely a downstream side effect of autism's broader social communication deficits 4243.
Superior Face Recognition Ability
Occupying the opposite end of the distribution from prosopagnosia are "super-recognizers" (SRs) - individuals possessing extraordinary facial identity processing capabilities. SRs perform at or above the 98th percentile on standardized objective tests designed to assess perceptual discrimination and long-term face memory, such as the Cambridge Face Memory Test (CFMT+), the Oxford Face Matching Test, the Facial Identity Card Sorting Test (FICST), and the Yearbook Test 444551.
The neurocognitive mechanisms driving this superior performance remain a subject of intense investigation. High-density EEG studies utilizing multivariate decoding approaches indicate that the brains of SRs process faces differently across varying temporal windows. Distinct representational differences between typical individuals and SRs emerge during early visual processing stages (around 150 milliseconds, corresponding to structural encoding) and persist into late processing stages (around 600 milliseconds, corresponding to semantic and mnemonic access) 45.
The identification and study of super-recognizers hold significant applied value. Law enforcement and forensic agencies actively recruit individuals with these specific cognitive profiles to perform complex tasks such as perpetrator identification, CCTV monitoring, and cross-border identity verification 4446. Studies utilizing forensically authentic material, such as the Berlin Test for Super-Recognizer Identification, confirm that lab-identified SRs demonstrate exceptional proficiency in operationally relevant 1-to-many face matching tasks. This confirms that their lab-based perceptual superiority translates directly to real-world forensic expertise, including emerging challenges like deepfake detection 4446. Furthermore, research is currently expanding into multi-modal domains to determine if "voice super-recognizers" exist, utilizing tools like the Zürich Voice Super-Recognizer Test (ZVSRT) to assess superior voice identity processing 46.