Neuroscience and evolution of the human need to belong
The human drive to form and maintain stable, positively valenced interpersonal relationships operates as a primary, evolutionarily hardwired biological imperative 123. Formalized comprehensively in the "need to belong" hypothesis, the desire for interpersonal attachment is recognized to be as fundamental to human survival and motivation as physiological drives for sustenance and shelter 234. Various psychological frameworks, including Maslow's Hierarchy of Needs and Self-Determination Theory (SDT), position relatedness and belonging as prerequisites for optimal psychological functioning, autonomy, and competence 1. When this need is satisfied, neurobiological systems maintain homeostasis, facilitating cognitive development, emotional regulation, and physical health 12. Conversely, when this drive is thwarted through social exclusion, interpersonal rejection, or chronic isolation, it precipitates a cascade of severe neurobiological, endocrine, and immunological disruptions 15.
The profound sensitivity of the human central nervous system to social acceptance and rejection represents millions of years of evolutionary adaptation. Early hominids relied entirely on group cohesion for foraging, predator defense, and rearing highly altricial infants 166. Consequently, the brain evolved sophisticated neural architectures to monitor social status, detect threats of exclusion, and reward prosocial affiliation 678. Understanding the neurobiology of the need to belong requires a synthesis of evolutionary anthropology, cognitive neuroscience, cross-cultural psychology, and psychoneuroimmunology.
Evolutionary Anthropology of Social Cohesion
Human sociality differs profoundly from that of other species, primarily characterized by unprecedented levels of cooperation, complex verbal communication, and the capacity for cumulative culture 2. The psychological mechanisms underlying the need to belong were forged in ancestral environments, where group living offered definitive survival and reproductive advantages over solitary existence 16.
Survival Benefits and the Costs of Ostracism
Throughout human evolutionary history, inclusion in a social group provided protection from predators, facilitated the sharing of resources during periods of scarcity, and enabled cooperative care for infants 166. Evolutionary models suggest that because the fitness costs of being isolated were overwhelmingly severe, natural selection favored individuals possessing a hypersensitive cognitive alarm system capable of detecting subtle cues of social disapproval or marginalization 6.
This evolutionary reliance on group inclusion explains persistent psychological phenomena observed in modern humans. Because group inclusion was universally beneficial and exclusion universally lethal in the ancestral environment, the human brain lacks a specialized cognitive mechanism to process "beneficial exclusion" or "costly inclusion" 6. Experimental paradigms utilizing virtual ball-tossing games demonstrate this evolutionary mismatch: individuals continue to experience acute psychological distress and declines in self-reported mood when excluded, even in experimental settings where being excluded yields a direct financial payout, or where inclusion costs money 6. The innate aversion to ostracism operates independently of higher-order rationalization, functioning much like a reflex designed to preserve social integration at all costs.
Ostracism as a Regulatory Mechanism
While ostracism is devastating to the excluded individual, it serves a vital regulatory function for the survival of the group. Evolutionary game theory models, particularly those simulating public goods scenarios, indicate that cooperation within a group cannot be sustained if defectors, rule-breakers, and free-riders are allowed to exploit collective resources without consequence 910. Ostracism functions as a costless punishment mechanism that removes non-cooperative members, thereby protecting the group's integrity, enforcing social norms, and deterring prospective norm-breakers 910.
Furthermore, the threat of ostracism, mediated by reputational tracking and gossip, acts as a powerful deterrent against selfish behavior. Experimental studies demonstrate that when individuals know their behavior can be communicated to future interaction partners via gossip, cooperation levels rise significantly 11. In environments where reputational concerns are salient, individuals actively engage in "competitive altruism," deliberately contributing to the public good to secure their social standing, avert the threat of exile, and demonstrate their relational value to the collective 11.
Neural Mechanisms of Social Rejection
The profound behavioral aversion to social exclusion is orchestrated by highly specialized neural networks. Functional magnetic resonance imaging (fMRI) has extensively mapped the brain's response to social rejection, though the precise interpretation of these regional activations has undergone significant theoretical evolution over the past two decades.
The Social Pain Hypothesis
The earliest prevailing framework for understanding the neurobiology of exclusion is the "social pain hypothesis." This theory posits that the evolutionary imperative to maintain social bonds co-opted existing neural circuitries originally dedicated to processing physical nociception (pain) 121314. In a seminal 2003 study utilizing the Cyberball paradigm - a virtual ball-tossing game where participants are unexpectedly excluded by computerized avatars - researchers observed robust activation in the dorsal anterior cingulate cortex (dACC) and the anterior insula (AI) 4141516.
Because the dACC and AI are heavily implicated in the affective distress component of physical pain, researchers hypothesized that social rejection genuinely "hurts" at a neurological level 41417. Under this model, the shared neural circuitry acts as an alarm system, ensuring that social isolation captures attention and motivates the individual to repair broken social bonds, mirroring how physical pain motivates the avoidance of tissue damage 41317.
The Anterior Cingulate Cortex Debate
Despite the prominence of the social pain hypothesis, the specific role of the dACC during social exclusion generated intense scientific debate. Critiques argued that the dACC's response during the Cyberball task might not reflect the emotional distress of rejection, but rather the cognitive processing of an "expectancy violation" 41819. Because human subjects implicitly expect to be included in a fair game, sudden exclusion violates a cognitive script, thereby triggering the dACC's well-established function in conflict monitoring and discrepancy detection 418.
Subsequent studies attempted to uncouple expectancy violation from social rejection by utilizing modified paradigms. Studies incorporating "over-inclusion" (where participants receive an unexpectedly high amount of ball tosses) or explicit social evaluation tasks yielded mixed results 41920. Some data suggested that the dACC responds primarily to cognitive surprise, while the ventral anterior cingulate cortex (vACC) and subgenual ACC (sgACC) are more closely aligned with the subjective affective distress of rejection 18192021.
Default Mode Network and Meta-Analytic Findings
Recent large-scale analyses have challenged the centrality of the dACC in social exclusion entirely. A comprehensive 2021 meta-analysis utilizing activation likelihood estimation (ALE) across 53 Cyberball fMRI studies (encompassing 1,817 participants) found that the dACC was not reliably activated across the literature 222324. When using a probabilistic atlas to define the region, fewer than 15% of the analyzed studies reported peak coordinates in the dACC 222324.
Instead, consistent recruitment was observed in the ventral anterior cingulate, posterior cingulate cortex (PCC), inferior frontal gyrus (IFG), and posterior insula 222324. Meta-analytic connectivity mapping indicated that the neural response to social exclusion aligns closely with the topography of the Default Mode Network (DMN), particularly regions associated with mentalizing, autobiographical memory, and theory of mind (e.g., medial prefrontal cortex, temporoparietal junction, and precuneus) 162224. This suggests that social exclusion primarily prompts extensive socio-cognitive processing as individuals attempt to infer the motives of their excluders and reassess their social standing.
Relational Value Computations
Further clarifying these neural mechanisms, a 2024 neurocomputational study dissociated two distinct learning pathways involved in social affiliation: tracking "rewarding outcomes" versus tracking "relational value" 2526.
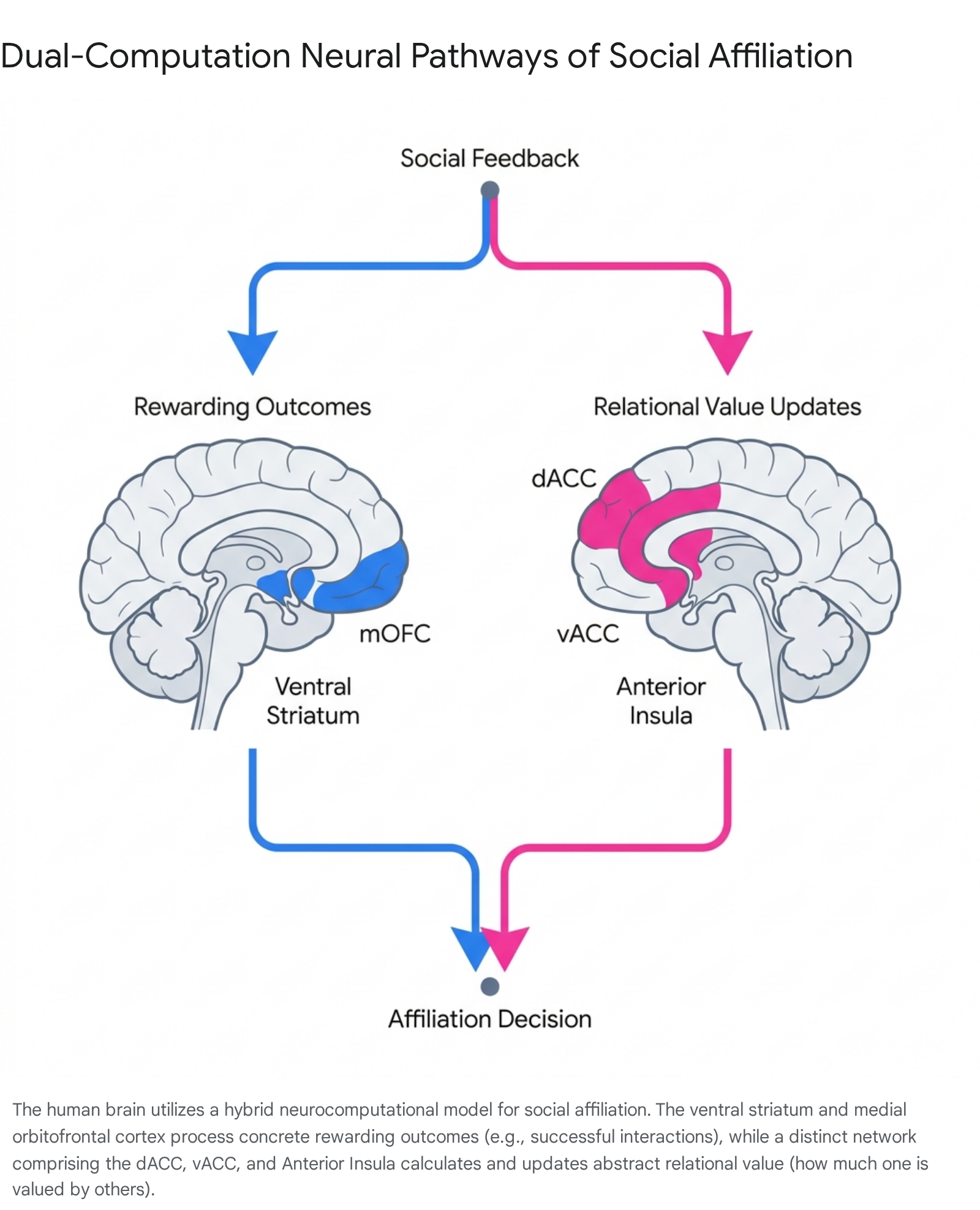
Utilizing computational modeling alongside neuroimaging, the researchers discovered that regions historically associated with the "social rejection network" (dACC, vACC, AI, and ventrolateral prefrontal cortex) do not exclusively encode negative "pain" signals.
Rather, these regions compute unsigned updates to an individual's relational value 25. When participants learned how much another person valued them (e.g., their abstract social rank), these brain areas activated in response to updates in that internal model, regardless of whether the update was positive or negative 25.
This neurocomputational model indicates that the brain builds an abstract internal map of social hierarchies and relational worth. While subjective distress occurs when this computed relational value drops precipitously, the primary function of the network is predictive social learning and value updating rather than mere nociception 25.
Experimental Paradigms in Social Neuroscience
The robust insights regarding the neural correlates of acceptance and rejection rely heavily on standardized experimental paradigms. Methodological differences in how exclusion is simulated heavily influence the neural networks recruited during fMRI studies.
The Cyberball Paradigm
Developed by Williams and colleagues, Cyberball remains the most ubiquitous behavioral paradigm used to study ostracism in fMRI environments 222327. In this virtual ball-tossing game, participants believe they are interacting with human peers, though the avatars are pre-programmed. Following a period of fair inclusion, the participant is systematically excluded from the game 2227.
Cyberball's utility lies in its simplicity and reliability in inducing immediate, measurable psychological distress, threatening fundamental needs such as belonging, self-esteem, meaningful existence, and control 428. However, the paradigm is highly sanitized, lacking the rich, multimodal sensory components of real-world interpersonal rejection. Furthermore, its inherent conflation of social rejection with expectancy violation necessitates the use of supplementary task designs 41819.
Social Evaluation and Immersive Paradigms
To address the limitations of Cyberball, researchers utilize alternative methodologies that isolate specific social variables:
- Social Evaluation Tasks: In paradigms such as the Social Judgment and Chatroom tasks, participants submit personal profiles or photographs and later receive direct positive or negative feedback from ostensible peers 1929. These tasks allow researchers to disentangle expected versus unexpected feedback, isolating the pure neural response to social evaluation from cognitive surprise 1929.
- Overinclusion Tasks: A variant of the ball-tossing game where the participant receives a surprisingly large number of throws 427. This isolates the variable of expectancy violation without the negative valence of rejection, revealing activation in the dACC and temporary ameliorative effects on psychological functioning 427.
- Immersive Social Environments: Newer adaptations, such as the "Social Ball," attempt to increase ecological validity by mimicking modern digital environments 30. These tasks incorporate visual immersion, hand-waving avatars, and chat features, attempting to mirror contemporary online social dynamics where cyber-ostracism frequently occurs 3031.
Neural Mechanisms of Social Acceptance and Reward
If social rejection triggers the brain's alarm and value-updating systems, social acceptance heavily engages the brain's mesolimbic reward circuitry. The biological imperative to belong is reinforced by positive hedonic feedback elicited by affiliative interactions, cooperative behavior, and social approval 73233.
The Ventral Striatum and Reward Circuitry
The core of the brain's reward system - particularly the ventral striatum (VS), which encompasses the nucleus accumbens (NAcc), alongside the ventromedial prefrontal cortex (vmPFC) and orbitofrontal cortex (OFC) - is robustly activated by social acceptance 733343536. When individuals receive positive social feedback, experience reciprocal cooperation, or view socially approving faces, dopaminergic projections from the ventral tegmental area (VTA) to the NAcc generate signals of subjective pleasure and reinforcement 7323337.
The brain processes social rewards using the same basal ganglia and prefrontal architecture employed for primary survival rewards (e.g., food) and secondary monetary rewards 73438. However, while the dorsal and ventral striatum are engaged during the anticipation of both social and monetary rewards, the consumption phase of social rewards elicits distinct neural patterns, heavily recruiting the putamen, bilateral insula, and specific frontal areas not utilized for monetary consumption 3839.
Neurochemical Modulation by Oxytocin
The rewarding nature of social inclusion is fundamentally modulated by the neuropeptide oxytocin (OT). Synthesized primarily in the paraventricular nucleus (PVN) of the hypothalamus, oxytocin is critical for pair bonding, maternal care, and the establishment of social trust 404142. Oxytocin exerts its influence by interacting directly with the mesolimbic dopamine system; oxytocin receptors (OTRs) are highly expressed in the VTA and NAcc 4143.
Oxytocin signaling functions to fine-tune the signal-to-noise ratio in social neural circuits, amplifying the salience of social cues over non-social stimuli 4144. Epigenetic regulation of the oxytocin receptor gene (OXTR) - such as varying levels of DNA methylation - can alter the density and sensitivity of these receptors in the brain. Individuals with highly methylated OXTR genes, or specific common genetic variants, demonstrate altered ventral striatal and amygdala reactivity to social stimuli, influencing their sensitivity to social reward and their vulnerability to social-affective dysregulation 4344. Furthermore, endogenous oxytocin release during human social interactions (such as physical touch) is dynamic and context-dependent, modulating subsequent neural responses in the medial prefrontal cortex and correlating with reductions in systemic cortisol 45.
Differentiating Social and Monetary Rewards
While social and monetary rewards share overlapping neural substrates, they drive distinct behavioral outcomes. Monetary rewards are typically associated with self-interest, competence, and individual utility, whereas social rewards are intrinsically linked to affiliation, group cohesion, and mutual well-being 46.
When participants engage in decisions that result in non-costly social benefits to others (prosocial choices), robust activation is observed in the vmPFC and dorsomedial prefrontal cortex (dmPFC) 47. Individuals with a baseline selfish orientation require greater activation in these prefrontal cognitive control regions to make prosocial choices, suggesting that overriding egoistic impulses requires explicit, deliberative neural effort 47. In contrast, highly prosocial individuals exhibit intuitive, rapid recruitment of the striatal reward system during cooperative acts, highlighting how individual differences in prosociality modulate the brain's fundamental reward architecture 3947.
Table 1 summarizes the primary brain networks associated with different facets of the human belongingness drive.
| Social Phenomenon | Primary Brain Regions Implicated | Core Cognitive / Affective Function |
|---|---|---|
| Social Exclusion / Rejection | Anterior Insula (AI), Ventral ACC, Inferior Frontal Gyrus (IFG) | Processing affective distress, negative arousal, and emotional regulation 414234849. |
| Relational Value Updating | Dorsal ACC, Ventral ACC, Anterior Insula, Ventrolateral PFC (vlPFC) | Tracking abstract social rank and computing updates to an internal model of social value 25. |
| Social Reward / Acceptance | Ventral Striatum (Nucleus Accumbens), Ventral Tegmental Area (VTA), Medial OFC | Dopaminergic reinforcement learning, processing positive social feedback and cooperative outcomes 7323335. |
| Social Mentalizing / DMN | Medial Prefrontal Cortex (mPFC), Temporoparietal Junction (TPJ), Precuneus | Theory of mind, inferring others' intentions, processing the social context of inclusion or exclusion 162435. |
Cross-Cultural Variations in Social Cognition
The universal biological drive to belong is heavily mediated by the cultural context in which an individual develops. Cultural neuroscience has established that divergent societal values - specifically individualism versus collectivism - profoundly shape the neural representations of the self and the socio-cognitive processing of rejection 50515253.
Individualism, Collectivism, and Neural Representation
Western cultures tend to be highly individualistic, promoting an "independent" self-construal where identity is defined by unique personal traits, autonomy, and distinct boundaries between the self and others 505455. Conversely, East Asian cultures are predominantly collectivistic, fostering an "interdependent" self-construal where identity is intrinsically tied to social roles, relationships, and group harmony 505556.
Quantitative meta-analyses of fMRI studies reveal that these cultural paradigms recruit distinct neural networks during social cognition. East Asian participants demonstrate stronger activity in the dorsomedial prefrontal cortex (dmPFC) and temporoparietal junction (TPJ) - regions essential for inferring the mental states of others, understanding social context, and emotion regulation 5257. Western participants, however, exhibit enhanced activity in the anterior cingulate, ventral medial prefrontal cortex, and bilateral insula, reflecting an emphasis on self-relevance encoding and direct emotional reactivity during social tasks 5257.
Neural Correlates of the Interdependent Self
The deep interconnection between the self and the social group in collectivist cultures is visibly mapped in the brain's functional anatomy. When Western individuals perform trait-judgment tasks evaluating themselves versus evaluating a close relative (e.g., their mother), functional imaging reveals distinct, separate patterns of activation in the medial prefrontal cortex (mPFC) 53555658. In stark contrast, when Chinese participants perform the exact same task, the mPFC exhibits overlapping neural activation for judgments of the self and the mother 53555658. This shared neural representation provides a literal biological substrate for the interdependent self-construal, demonstrating that cultural values dictate how closely the self is tethered to social affiliates at the neurological level.
Cultural Moderation of Rejection Responses
Cultural values also dictate the neurocognitive strategies utilized to cope with social exclusion. Research investigating African American and White participants utilizing the Cyberball task demonstrated that cultural priming alters frontal lobe recruitment during ostracism. Specifically, individualistic values moderate the experience of rejection through the recruitment of explicit, top-down cognitive control areas, notably the dorsolateral prefrontal cortex (dlPFC) 5059. In contrast, collectivistic values operate as a coping mechanism mediated through implicit, socioemotional regulation regions of the brain, such as the ventromedial prefrontal cortex (vmPFC) 50. Therefore, the neurobiology of belonging is not merely a static evolutionary inheritance but a plastic system continuously molded by the cultural environment.
Table 2 highlights the distinct neural responses between individualist and collectivist populations during social cognitive tasks.
| Cultural Orientation | Dominant Self-Construal | Distinct Neural Activation Patterns | Processing Characteristics |
|---|---|---|---|
| Western / Individualist | Independent Self | Enhanced ACC, bilateral insula, distinct mPFC for self vs. others 525657. | Focus on self-relevance encoding, distinct boundaries between self and affiliates 505255. |
| East Asian / Collectivist | Interdependent Self | Enhanced dmPFC, TPJ, overlapping mPFC for self and close others 52535657. | Focus on mentalizing, context inference, shared neural representation of self and family 525558. |
Systemic Biological Impacts of Chronic Loneliness
When the need to belong is chronically unmet, the psychological state of loneliness translates into severe physiological deterioration. Loneliness is not merely an emotional affliction; it is a profound biological threat. Epidemiological data equate the mortality risk of prolonged social isolation to smoking 15 cigarettes a day, driving pathogenesis across multiple organ systems 60616263.
Endocrine Dysregulation and the HPA Axis
The primary mechanism by which social isolation gets "under the skin" is through the chronic hyperactivation of the body's stress response systems. The nervous system interprets social disconnection as an imminent physical threat, activating the hypothalamic-pituitary-adrenal (HPA) axis and the sympathetic nervous system (SNS) 60616465.
In socially isolated individuals, this prolonged activation results in a severely dysregulated diurnal cortisol rhythm. While healthy individuals display a sharp cortisol peak in the morning that steadily declines throughout the day, lonely individuals frequently exhibit a flattened diurnal cortisol slope, characterized by persistently elevated cortisol levels that fail to decline appropriately during evening hours 56667. This chronic cortisol exposure desensitizes glucocorticoid receptors, impairing the body's ability to regulate subsequent stress and contributing directly to insulin resistance, hypertension, and disrupted sleep architecture (fragmented REM and N3 phases) 5616264.
Immune System Reconfiguration and Inflammation
The SNS overdrive associated with loneliness triggers a highly specific reconfiguration of the immune system known as the Conserved Transcriptional Response to Adversity (CTRA) 568. From an evolutionary standpoint, an isolated hominid faced a higher risk of physical wounding and bacterial infection (due to a lack of group protection), but a lower risk of socially transmitted viral diseases 5. Consequently, chronic loneliness prompts the immune system to upregulate genes responsible for pro-inflammatory responses while simultaneously downregulating genes involved in antiviral defense 568.
Empirical measurements of blood and tissue samples from lonely individuals confirm this systemic inflammatory state. Social isolation is consistently associated with elevated circulating levels of pro-inflammatory cytokines and acute-phase proteins, particularly Interleukin-6 (IL-6), Tumor Necrosis Factor-alpha (TNF-α), and high-sensitivity C-Reactive Protein (CRP) 5636465666769. This low-grade, persistent inflammation creates a biological environment highly conducive to the development of cardiovascular disease, neurodegeneration, and premature mortality 56165.
Epigenetic Modifications and Accelerated Aging
The physical impact of social isolation extends to the molecular level, altering gene expression without changing the underlying DNA sequence. Epigenome-wide association studies have identified specific DNA methylation patterns intrinsically linked to chronic loneliness 6070. For instance, social isolation frequently induces methylation of the BDNF (Brain-Derived Neurotrophic Factor) gene, suppressing its transcription. This suppression impairs neuroplasticity and synaptic growth, drastically increasing susceptibility to cognitive decline and mood disorders 6071. Similarly, methylation of glucocorticoid receptor genes (NR3C1) blunts the negative feedback loop of the HPA axis, locking the body into a state of continuous stress reactivity 60.
Furthermore, analyses of methylation data from adult twins have revealed that individuals experiencing high levels of loneliness score significantly higher on the DunedinPACE epigenetic clock - a recognized biomarker that measures the exact rate at which a human body is biologically aging 6870. This indicates that the lack of social connection accelerates cellular aging, causing the body's systems to wear down at a demonstrably faster rate than in socially integrated counterparts 6870.
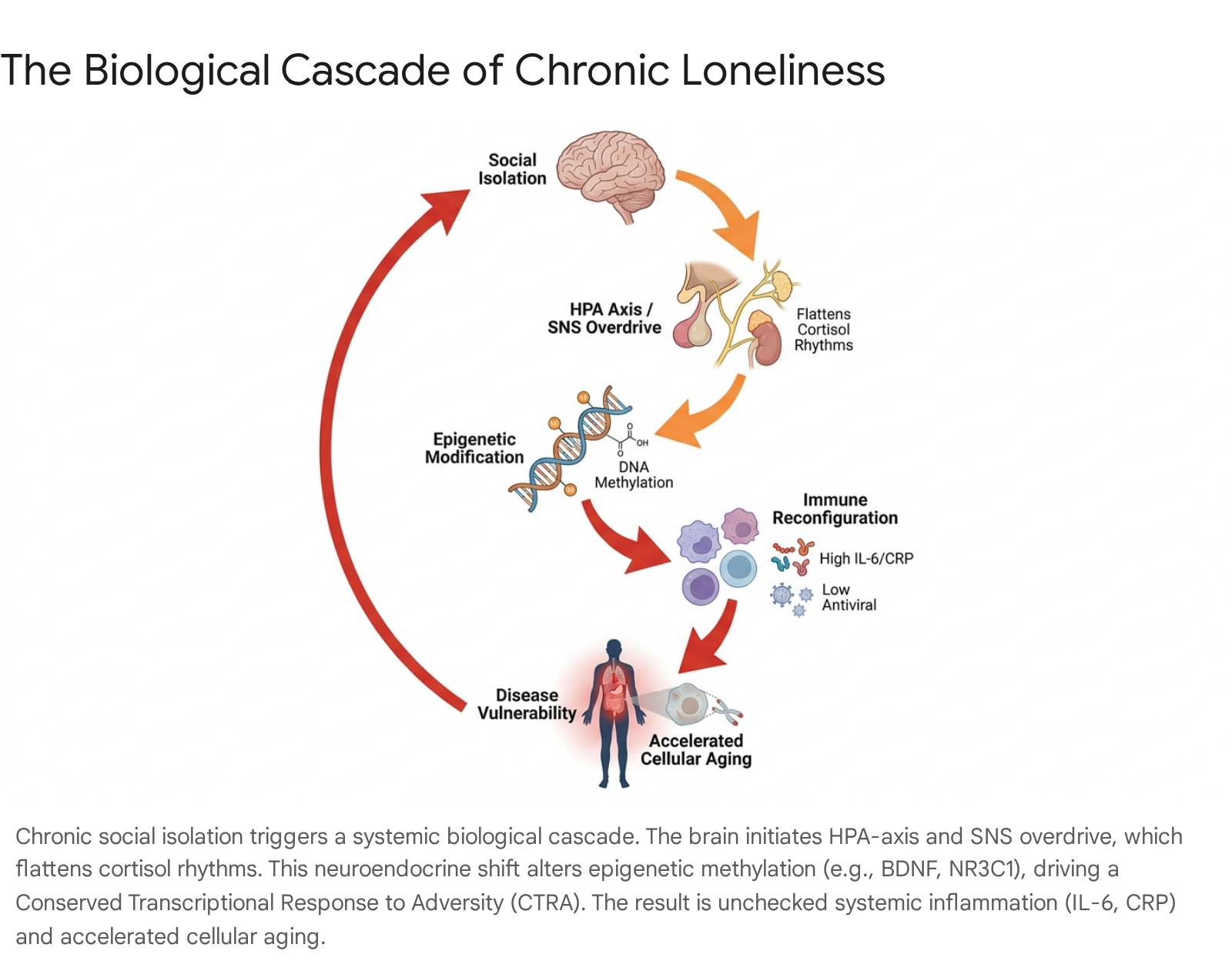
Table 3 categorizes the distinct systemic biomarkers utilized to quantify the physiological toll of chronic social isolation.
| Biomarker Category | Specific Markers | Physiological Impact of Chronic Loneliness |
|---|---|---|
| Neuroendocrine | Cortisol, Norepinephrine | Flattened diurnal cortisol slope; chronic HPA-axis and sympathetic nervous system hyperactivation 56466. |
| Inflammatory / Immune | IL-6, CRP, TNF-α | Upregulation of systemic pro-inflammatory cytokines; compromised antiviral immunity and diminished vaccine response 563646569. |
| Epigenetic | DNA Methylation (BDNF, NR3C1), DunedinPACE | Altered gene expression suppressing synaptic plasticity; accelerated epigenetic clock indicating faster biological aging 60687071. |
Transgenerational Epigenetic Inheritance
The biological scars of social isolation are not strictly confined to the individual experiencing them. Emerging research on transgenerational epigenetic inheritance suggests that severe social stress can imprint upon the germline 60627273. When an organism experiences chronic isolation, the resulting hypermethylation of stress-response genes can be transmitted to subsequent generations 6273. Consequently, the offspring of severely socially isolated individuals may be born with a pre-configured, hypersensitive HPA axis and a heightened vulnerability to anxiety and inflammatory disorders, regardless of their own immediate social environment 606273. While environmental interventions (such as social enrichment) can mitigate these transmitted sensitivities, the data highlights that social isolation exerts multi-generational impacts 6062.
Conclusion
The human need to belong is a profound evolutionary legacy, deeply inscribed into the species' neurobiology, endocrinology, and genetic expression. The brain's architecture processes social rejection not as a trivial emotional inconvenience, but as an acute threat to survival, mobilizing neurocomputational networks to update relational value models and deploying distress signals to force reintegration. Simultaneously, the mesolimbic reward system strongly reinforces social acceptance, utilizing dopaminergic and oxytocinergic pathways to bind communities together.
When society fails to meet this fundamental biological requirement, the resulting chronic loneliness triggers a devastating systemic cascade. The flattening of cortisol rhythms, the upregulation of systemic inflammation via the CTRA, and the acceleration of epigenetic aging all underscore a singular scientific reality: humans are an obligatorily gregarious species. Connection is not merely a cultural luxury; it is a physiological necessity required to sustain the biological integrity of the human organism.