Neuroscience of awe and its impact on time and self-boundaries
Neurobiological Foundations of Awe
Awe is a complex, self-transcendent emotion triggered by perceptually vast stimuli that challenge and ultimately transcend an individual's current frames of reference. Originating from encounters with expansive natural panoramas, profound artistic expressions, conceptual epiphanies, or the collective effervescence of spiritual practices, awe demands cognitive accommodation. This requirement for accommodation - the restructuring of mental schemas to integrate novel, overwhelming information - distinguishes awe from related affective states such as joy, surprise, or amusement 123.
Historically relegated to the realms of philosophy, theology, and qualitative psychology, awe has recently become a focal point of affective neuroscience. Neuroimaging techniques, primarily functional magnetic resonance imaging (fMRI) and voxel-based morphometry (VBM), have begun to map the neural architecture underlying this profound psychological state. The resulting data indicates that awe is not merely an elevated form of positive affect but a distinct neurobiological state characterized by widespread functional reorganization across large-scale brain networks, significant alterations in autonomic nervous system arousal, and systemic physiological shifts 124.
The defining phenomenological outcome of awe is the "small self" - a subjective diminution of the ego and a corresponding increase in perceived connectedness to the broader environment, humanity, or a higher conceptual order 1256. This psychological shift is mirrored by specific neural signatures, most notably the suppression of cortical regions responsible for self-referential processing, combined with the activation of networks that direct attention outward. Furthermore, recent scientific inquiries have bifurcated awe into two distinct psychological and neurobiological variants: positive awe, which aligns with aesthetic reward and prosociality, and threat-based awe, which is imbued with fear, uncertainty, and sympathetic autonomic arousal 789.
Large-Scale Brain Network Dynamics
Complex affective and cognitive states do not map neatly onto isolated brain regions; rather, they emerge from the dynamic interplay of large-scale neural networks. The experience of awe exemplifies this phenomenon, heavily modulating the activity within and between the Default Mode Network, the Salience Network, and the Central Executive Network 101112.
Default Mode Network Suppression
The Default Mode Network (DMN), comprising interconnected regions including the medial prefrontal cortex (mPFC), posterior cingulate cortex (PCC), precuneus, and inferior parietal lobule, is primarily active during rest, mind-wandering, and self-referential thought 1510. Activity in the DMN correlates with introspection, autobiographical memory retrieval, egoic narrative, and rumination.
During the experience of awe, fMRI studies demonstrate a marked downregulation of DMN activity 1451314. In controlled laboratory settings, researchers utilizing fMRI to scan participants viewing awe-inspiring natural phenomena (e.g., sweeping vistas from nature documentaries) contrasted against those viewing amusing animals or neutral landscapes found a distinct divergence in neural activation. When participants passively watched amusing or neutral videos, activity in the DMN increased. However, passive viewing of awe-inducing videos suppressed DMN activation significantly 114.
This neural dampening provides a physiological basis for the subjective "small self" and the cessation of egoic rumination reported during transcendent experiences 14614. The temporary dissolution of self-boundaries in awe shares striking neural similarities with other self-transcendent states, notably deep meditative absorption and the acute effects of classical psychedelics. Both rigorous meditation and psychedelic interventions are characterized by profound DMN deactivation and subsequent ego dissolution, positioning awe as an accessible, naturally occurring analog to these transformative states 1101615.
Salience and Central Executive Networks
The profound DMN suppression observed in awe is actively orchestrated by the Salience Network (SN). Anchored by the anterior insula and the dorsal anterior cingulate cortex (dACC), the SN contains specialized von Economo neurons and is responsible for detecting emotionally and homeostatically relevant stimuli in the environment, subsequently allocating neural resources 101116.
When an individual encounters a perceptually vast stimulus, the SN identifies this profound deviation from mundane expectations and acts as a dynamic cognitive switch. It downregulates the DMN, suspending the internal narrative, and simultaneously upregulates the Central Executive Network (CEN), also referred to as the fronto-parietal network 1101217.
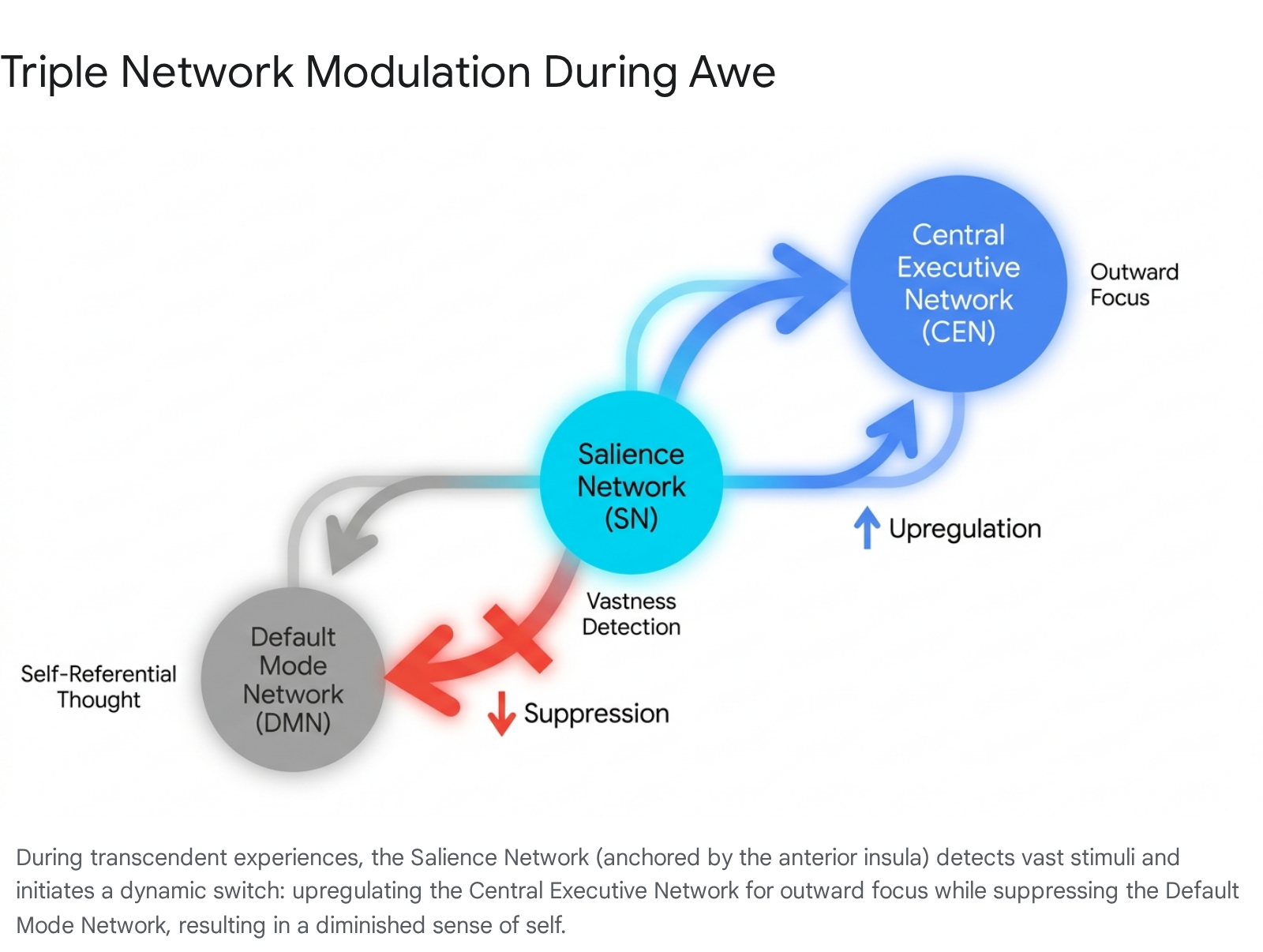
The CEN supports externally directed attention, working memory, and goal-directed cognitive processing. Elevated activity in the fronto-parietal network during awe underlines the captivating, immersive, and attention-grabbing nature of the emotion 114. When participants in fMRI studies were asked to count perspective changes while watching awe-inducing videos, activity in the fronto-parietal network increased significantly more than during exposure to other video types, demonstrating how awe pulls the individual entirely into the present sensory reality 114.
Schema Liberation and the Middle Temporal Gyrus
The second core component of awe - the need for cognitive accommodation - has been localized to specific temporal lobe dynamics. The left middle temporal gyrus (MTG) plays a critical role in matching existing cognitive schemas and predictive models to incoming sensory events 718.
Functional MRI studies contrasting awe against control emotions (such as amusement or fear) reveal that both positive and threat-based awe experiences consistently deactivate the left MTG 71819. This deactivation is interpreted as a precise neural correlate for the "schema liberation" process. Because awe-inducing stimuli are too vast or complex to be assimilated into pre-existing mental models, the brain must temporarily suspend its standard predictive coding mechanisms. By deactivating the MTG, the brain effectively drops its current conceptual frameworks, allowing the individual to process the profound novelty of the experience without forcing it into inadequate, pre-existing cognitive categories 718.
Structural and Functional Divergence in Awe Variants
While early conceptualizations framed awe uniformly as a positive, self-transcendent state, contemporary affective science recognizes a bifurcation based on the nature of the eliciting stimulus. "Positive awe" arises from aesthetic beauty, spiritual epiphanies, or human virtue. Conversely, "threat-based awe" (or negative awe) is triggered by vast, overwhelming forces that carry potential danger, such as natural disasters, predatory magnitude, or coercive charisma 789. These two variants share the core appraisals of vastness and the need for accommodation but diverge sharply in their neuroanatomical correlates, functional connectivity, and physiological arousal profiles.
Gray Matter Volume Correlates
Voxel-based morphometry (VBM) analyses reveal that individual predispositions toward experiencing positive versus threat-based awe are associated with distinct structural differences in regional gray matter volume (rGMV) 3820.
Individuals prone to positive awe exhibit greater rGMV in the precuneus, a region implicated in sophisticated self-awareness, reflective processing, and subjective life satisfaction 82021. Conversely, positive awe propensity is negatively correlated with rGMV in the left fusiform gyrus and right calcarine cortex, regions typically associated with basic visual and facial processing 382021.
In contrast, an individual's propensity for threat-based awe is negatively correlated with rGMV in the bilateral insula and the left superior temporal gyrus (STG) 3820. The insula is a primary hub for interoception, automatic threat detection, and the processing of fear and anxiety 8. This structural correlation suggests that the threshold for experiencing threat-awe is deeply tied to the neurocircuitry that maps homeostatic states and risk awareness.
Divergent Functional Connectivity Pathways
Though both forms of awe deactivate the left MTG to facilitate schema liberation, they route this liberated cognitive state through entirely different functional networks, resulting in vastly different phenomenological experiences 719.
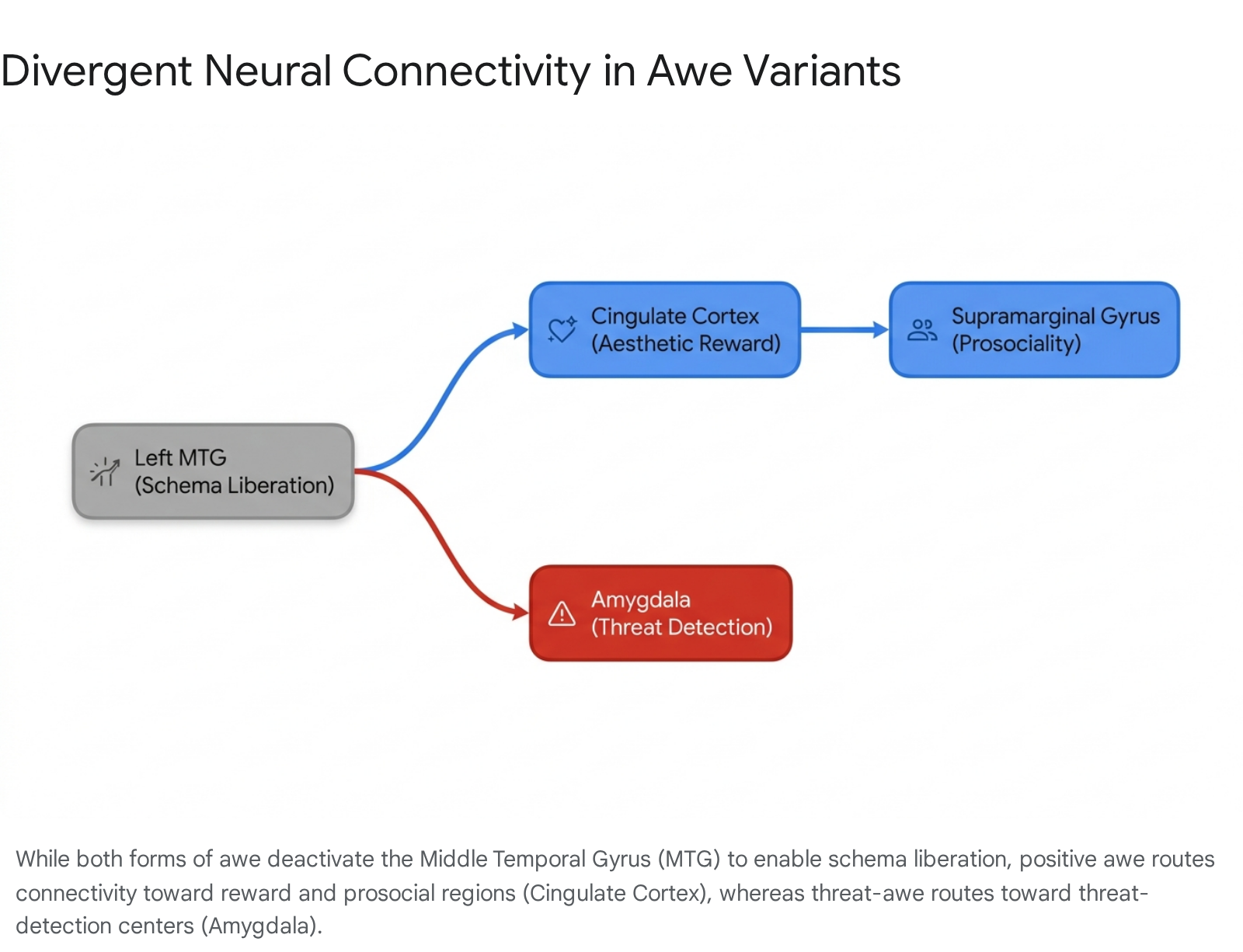
During positive awe, there is heightened functional connectivity between the MTG and both the anterior/posterior cingulate cortex (ACC/PCC) and the supramarginal gyrus (SMG). The cingulate structures are deeply integrated into the brain's aesthetic reward and valuation systems, explaining why positive awe feels euphoric, inspiring, and intrinsically rewarding 7. The SMG's involvement facilitates self-other representation, supporting the prosociality, empathy, and feelings of universal connection typically documented after positive awe experiences 722.
Conversely, threat-based awe demonstrates increased functional connectivity between the MTG and the amygdala, the brain's primary locus for detecting and processing threat stimuli. Furthermore, threat-awe increases connectivity between the amygdala and the SMG 719. This neural routing explains the phenomenological experience of threat-awe: a profound sense of vastness coupled with acute vulnerability, powerlessness, and fear, often culminating in a desire to seek safety within small, tight-knit communities rather than feeling a universal connection to humanity at large 48921.
The distinction between positive and threat-based awe is summarized in the table below, delineating the divergent neural, physiological, and behavioral outcomes.
| Metric | Positive Awe | Threat-Based Awe |
|---|---|---|
| Primary Elicitors | Natural beauty, human virtue, artistic majesty, epiphanies | Natural disasters, wrathful deities, predatory forces |
| Common MTG Response | Deactivation (Schema Liberation) | Deactivation (Schema Liberation) |
| Functional Connectivity | MTG ↔ Cingulate Cortex & Supramarginal Gyrus | MTG ↔ Amygdala & Supramarginal Gyrus |
| Autonomic Response | Increased Parasympathetic Arousal (Elevated Vagal Tone) | Increased Sympathetic Arousal (Fight/Flight Activation) |
| Gray Matter Correlates (VBM) | Higher GMV in Precuneus; Lower in Fusiform/Calcarine | Lower GMV in bilateral Insula and Superior Temporal Gyrus |
| Psychological Outcomes | Increased well-being, universal prosociality, connectedness | Lower momentary well-being, feelings of powerlessness, fear |
Alterations in Time Perception
One of the most persistent anecdotal features of transcendent experiences is the distortion of time. During profound moments of awe, individuals frequently report that time seems to slow down, expand, or stand entirely still 2324.
Subjective Expansion of Time
Behavioral research strongly supports the hypothesis that awe alters the subjective perception of time availability. In experiments contrasting awe with other emotions (such as happiness, neutral states, or amusement), participants primed with awe consistently reported feeling that they had more time available in the present moment 2325. This phenomenon is driven by awe's capacity to forcefully anchor attention in the immediate sensory environment. Because awe stimuli demand massive cognitive resources for accommodation, the brain temporarily abandons prospective (future-oriented) and retrospective (past-oriented) mental time travel, which are traditionally mediated by the DMN 12324.
The consequences of this subjective time expansion are highly prosocial and beneficial to well-being. Participants experiencing awe-induced time dilation exhibited less impatience, a greater willingness to volunteer their time to help others, a stronger preference for experiential over material goods, and greater overall life satisfaction 232425. By bringing individuals firmly into the present moment, awe acts as a buffer against the chronic "time famine" that characterizes modern psychological stress.
Neural Encoding of Time Duration
The neurological basis for subjective time perception is not localized to a single biological "clock" but is constructed hierarchically across multiple brain networks. Sensory input is processed by early visual cortices, read out by parietal regions, and finally categorized and assigned subjective duration in the frontal cortex and anterior insula 26.
The anterior insula, a critical node of the Salience Network, serves as the neural interface where physical objective reality is translated into subjective duration - what time "feels like" to the individual 101626. Because awe fundamentally alters the activity of the Salience Network and the anterior insula (which must adjudicate between the overwhelming external stimulus and internal homeostatic states), the process of subjective time construction is temporarily perturbed 1011121626. This disruption in the anterior insula's normal integrative function likely underpins the subjective feeling of time dilation during transcendent experiences.
Experimental Limitations in Implicit Time Perception
It is critical to note that while subjective questionnaires consistently capture the "time expansion" effect of awe, implicit and objective measures of time perception yield mixed results in laboratory settings. In temporal bisection tasks, where participants must implicitly categorize the duration of vibrotactile or visual stimuli while watching awe-inducing videos, researchers have not always found consistent effects of awe on implicit time perception 2427.
Some temporal bisection studies observe temporal lengthening, others note temporal shortening, and some find no significant divergence from positive control emotions 2427. These discrepancies suggest that laboratory-induced awe (e.g., viewing videos on a two-dimensional screen) may capture visual attention sufficiently to disrupt conscious time processing, but it may lack the ecological validity and total sensory immersion required to reliably trigger the deep neurobiological shifts that produce profound implicit time dilation 24. The failure to replicate time dilation uniformly in controlled bisection tasks underscores the difficulty of studying transcendent emotional states within the confines of laboratory environments.
Physiological Correlates and Autonomic Responses
The neural shifts that define awe trigger a cascade of peripheral physiological changes. These systemic alterations are responsible for the physical sensations that accompany transcendent experiences and contribute significantly to awe's well-documented physical and mental health benefits.
Vagal Tone and Systemic Inflammation
Positive awe exhibits a highly specific autonomic profile that differentiates it from other emotions. Unlike high-arousal positive emotions (like excitement, enthusiasm, or amusement) which activate the sympathetic nervous system, positive awe is associated with reduced sympathetic arousal and elevated parasympathetic activity, specifically increased vagal tone 419. The vagus nerve, which interfaces with the parasympathetic nervous system, acts to slow the heart rate, deepen respiration, and promote states of calm and social engagement. Threat-based awe, conversely, triggers classic sympathetic autonomic arousal, preparing the body for potential defensive action 4891928.
Furthermore, positive awe has been uniquely linked to the downregulation of systemic inflammation. Among a wide spectrum of positive emotions evaluated in longitudinal and daily diary studies, self-reports of awe are the most robust predictor of lower levels of Interleukin-6 (IL-6), a pro-inflammatory cytokine 429. Chronic elevation of IL-6 is associated with numerous autoimmune, cardiovascular, and depressive disorders. The neural dampening of the DMN and the elevation of vagal tone during awe appear to send regulatory signals that suppress this inflammatory biomarker. This suggests a direct mechanistic pathway between transcendent psychological experiences - such as nature immersion or deep spiritual contemplation - and physiological rejuvenation 416.
The Piloerection Discrepancy
A widely cited physiological correlate of awe in qualitative research and self-reports is the sensation of "chills" or "goosebumps" (piloerection). However, recent rigorous physiological investigations have revealed a surprising discrepancy between subjective human experience and objective measurement.
While individuals consistently self-report experiencing goosebumps during awe-inducing stimuli (often citing it as a primary physical indicator of the emotion), objective physiological monitoring of the arrector pili muscles - the skin structures responsible for piloerection - frequently shows no physical evidence of the response 3031. The arrector pili muscles and surrounding arterioles are heavily innervated by the sympathetic nervous system and utilize neurotransmitters and markers such as vasoactive intestinal peptide (VIP) 3032. Because positive awe generally downregulates sympathetic activity in favor of parasympathetic tone, literal piloerection is physiologically counterintuitive to the dominant awe state 3031.
Researchers suggest that the subjective feeling of "the chills" during awe may be a psychological phenomenon completely distinct from actual piloerection 3031. Because humans have evolved complex emotional responses, the neurological sensation of schema incongruence and profound novelty may be interpreted cognitively as "chills" without the accompanying sympathetic motor output required to contract the skin muscles 30. This divergence underscores the necessity of pairing self-report metrics with objective physiological data in affective neuroscience.
Cognitive Outcomes: Equanimity and Meaning
Recent empirical research has expanded the understanding of awe's psychological utility, demonstrating that it functions not just as a fleeting emotion, but as a catalyst for durable shifts in cognitive perspective and meaning-making.
Equanimity Through Temporal Distancing
A growing body of literature indicates that awe fosters emotional equanimity - a balanced, non-reactive state of mind toward all experiences, regardless of their positive or negative valence. Experimental inductions of awe, longitudinal diary studies, and even 5-day awe-walk interventions demonstrate that the experience of vastness reduces emotional reactivity to daily stressors 293533.
This cultivation of equanimity operates via the cognitive mechanism of temporal distancing. By instilling a sense of vastness, awe encourages individuals to adopt a broader temporal perspective, allowing them to view immediate stressors as transient and insignificant within the larger scope of time and existence 293533. Natural language processing (NLP) analyses of social media discourse confirm that individuals expressing awe-related language simultaneously exhibit higher levels of temporal distancing and equanimity, pointing to a robust effect in ecologically valid, real-world contexts 29.
Meaning in Life and the Boredom Pathway
Awe interacts with the human search for meaning through a complex dual pathway. Experimental evidence demonstrates that awe can simultaneously influence feelings of boredom and meaning in life through competing mechanisms depending on how the vastness is cognitively appraised 3334.
When awe results in a sense of "vastness vis-à-vis the self" - where the individual feels connected to something larger than themselves - it significantly increases the perceived meaning in life, which subsequently reduces feelings of boredom 34. However, if the awe experience results primarily in self-diminishment (feeling small, powerless, and insignificant in the face of the universe), it can have a neutral or even negative impact on meaning, potentially leaving boredom unchanged 34. This nuance highlights that the integration of the awe experience, rather than just the exposure to vast stimuli, dictates the long-term cognitive benefits.
Cross-Cultural Variations in the Awe Experience
The majority of early neuroimaging and behavioral research on awe was conducted using Western participant cohorts, leading to the conceptualization of awe as an overwhelmingly positive, prosocial emotion 3335. However, cross-cultural research spanning diverse populations demonstrates that the neural and physiological signatures of awe are subject to significant cultural modulation.
Studies comparing participant cohorts in the United States and China reveal divergent profiles of the awe experience. In rigorous two-week daily diary studies and standardized laboratory inductions, Chinese participants consistently reported significantly higher levels of fear, anxiety, and uncertainty during awe experiences compared to their American counterparts 353335. Phenomenologically, Western traditions tend to experience awe as oceanic, merging, and horizontal ("we are all connected in a circle"), whereas in Chinese cohorts, awe is often experienced as hierarchical and vertical (reverence for a great teacher, an ancient authority, or a vast, untouchable force) 39.
These psychological differences manifest physiologically. When exposed to identical awe-inducing stimuli, cultural cohorts exhibited divergent patterns of autonomic activity, particularly in heart rate reactivity, though measures of skin conductance and respiratory sinus arrhythmia remained statistically comparable 3335. When travelers from 42 countries were assessed at Yosemite National Park (a globally recognized awe-elicitor) and asked to draw their "small self" on graph paper, cultural variations in the size and conceptualization of the self were evident, underscoring the interplay between culture and spatial-emotional perception 2640.
This data suggests that the predominantly positive, reward-driven neural signature of awe mapped in Western laboratories may not be a universal human baseline. Outside of modern Western contexts, awe frequently operates as a deeply mixed emotion, heavily retaining the fear, threat-appraisal, and hierarchical positioning that likely characterized the evolutionary origins of the emotion 333539.
The comparative differences between Western and Eastern expressions of awe are outlined below.
| Cultural Context | Phenomenological Structure | Dominant Affective Tone | Physiological Markers | Self-Diminishment Style |
|---|---|---|---|---|
| Western (e.g., U.S.) | Oceanic, merging, horizontal connection | Highly positive, euphoric, inspiring | Lowered heart rate, parasympathetic dominance | Integration into a broader egalitarian whole |
| Eastern (e.g., China) | Hierarchical, vertical deference | Mixed valence, reverent, tinged with fear | Divergent heart rate reactivity profiles | Subordination to a greater authority or force |
Comparative Neurobiology: Awe, Flow, and Aesthetics
Awe is frequently conflated with other high-absorption states, such as "flow" and aesthetic appreciation. While these states share phenotypic similarities - such as intense focus, a departure from mundane self-referential thought, and time distortion - they operate via distinct neurobiological mechanisms.
Distinctions Between Awe and Flow
"Flow" is a state of optimal performance and deep absorption in a specific, highly practiced task (e.g., a musician improvising, an athlete competing) 141. Neurobiologically, flow is characterized by "transient hypofrontality" - a temporary downregulation of the prefrontal cortex, which suspends conscious self-monitoring, temporal processing, and critical thought to allow for rapid, effortless motor and cognitive execution 41.
Awe, conversely, is not a state of action or performance, but a state of perception and accommodation. While awe also involves reduced self-referential processing (via DMN suppression), it does not downregulate the executive networks. Instead, awe actively upregulates the Central Executive Network (the fronto-parietal network) to maximize external attention and cognitive processing of the vast stimulus 11441. Flow is a cognitive state optimizing internal action; awe is a perceptual state optimizing external absorption.
Overlap With Aesthetic Appreciation
Aesthetic appreciation - the cognitive and emotional evaluation of art, music, or beauty - recruits a triad of networks: the DMN (for affective evaluation and memory retrieval), the Salience Network (for monitoring emotional resonance), and the Central Executive Network (for associative thinking and mental set shifting) 173637.
Awe overlaps significantly with aesthetic appreciation but represents a more extreme neurobiological state. While aesthetic appreciation involves a balanced, synergistic interaction between these networks, awe fundamentally interrupts standard network coupling 131737. The profound vastness of an awe stimulus forces a distinct deactivation of the MTG for schema liberation, a feature not strictly required for general aesthetic enjoyment 738. Furthermore, positive awe triggers immense connectivity with the anterior/posterior cingulate cortex, hijacking the aesthetic reward process to produce transcendent euphoria rather than simple appreciation 7.
Convergence With Psychedelic States
The closest neurobiological analogue to profound awe is not flow or aesthetic appreciation, but the altered state of consciousness induced by classical psychedelics. Both awe and psychedelic states fundamentally disrupt high-level cognitive priors (predictive coding models) and violently suppress the Default Mode Network 11615.
Under the REBUS (RElaxed Beliefs Under psychedelics) model, psychedelics relax the brain's predictive coding hierarchy, allowing bottom-up sensory information to overwhelm top-down expectations, leading to ego dissolution and hyper-connectivity between normally segregated brain regions 16. Awe mimics this process endogenously via MTG deactivation and schema liberation. This shared disruption of entrenched neural hierarchies explains why both naturally occurring awe and psychedelic-assisted therapies are highly effective at breaking loops of depressive rumination, fostering cognitive flexibility, and instilling a durable sense of universal connectedness 1615.
The following table summarizes the neural network dynamics across these distinct but related cognitive states.
| Cognitive State | Default Mode Network (DMN) | Central Executive Network (CEN) | Key Neural Signature | Primary Function |
|---|---|---|---|---|
| Awe | Highly Suppressed | Upregulated (Fronto-Parietal) | MTG Deactivation (Schema Liberation) | External perception and cognitive accommodation |
| Flow | Suppressed | Downregulated (Transient Hypofrontality) | Prefrontal cortex suppression | Optimal performance and action execution |
| Aesthetic Appreciation | Active (Evaluation/Memory) | Active (Associative Thinking) | Balanced Tri-Network Synergy | Cognitive valuation of beauty/art |
| Psychedelic State | Severely Suppressed | Disorganized/Hyper-connected | Global reduction of predictive coding | Ego dissolution and synaptic neuroplasticity |
Methodological Considerations in Awe Research
As the field of affective neuroscience advances, several methodological challenges regarding the measurement and interpretation of awe have surfaced, demanding rigorous analytical frameworks.
Resting-State fMRI and Systemic Noise
Much of the literature regarding the baseline functional connectivity of individuals prone to awe relies on resting-state fMRI (rs-fMRI) metrics, such as the fractional amplitude of low-frequency fluctuations (fALFF) and regional homogeneity (ReHo) 3940. However, the BOLD signal utilized in fMRI is not purely a representation of neural activity; it is intrinsically linked to the vascular response.
Systemic physiological modulators, specifically cerebral blood flow (CBF) and cerebrovascular reactivity (CVR), easily influence these resting-state metrics 4041. Because awe alters autonomic function - modulating heart rate, vagal tone, and sympathetic arousal - it induces systemic cardiovascular changes that can artificially bias BOLD-based functional connectivity measurements 4041. Researchers must employ rigorous general linear models to account for the effect of physiological modulators like venous blood oxygenation and CVR when comparing resting-state networks (DMN, CEN, SN) in awe states versus control states. Failure to disentangle vascular response from neural activity may lead to overestimating the specific neural connectivity generated by awe.
Ecological Validity of Stimuli
Finally, the measurement of awe faces inherent challenges regarding ecological validity. The vast majority of neuroimaging studies rely on laboratory-induced awe, typically exposing participants to two-dimensional videos of nature or architecture while they lie immobilized in an MRI scanner 1714. While these methods successfully elicit self-reported awe and measurable neural shifts, they strip the experience of the multisensory immersion, physical risk, and spontaneous unpredictability that characterize real-world transcendent experiences 2433. The discrepancy in piloerection responses and implicit time perception between lab settings and anecdotal reality highlights the limitations of current experimental paradigms. Future research aims to bridge this gap through mobile neuroimaging technologies (such as wearable EEG) and immersive virtual reality environments, allowing for the capture of awe's neurological signatures in ecologically robust settings.