Neuromolecular mechanisms and modulation of critical periods
Introduction
The human brain's capacity to restructure itself in response to environmental stimuli - a phenomenon known as neuroplasticity - is arguably its most profound evolutionary adaptation. Historically, developmental neurobiology has operated under the assumption that this plasticity is strictly time-bound, confined to brief developmental windows that occur almost exclusively in early life. These windows represent phases of exuberant neural circuit remodeling, allowing the organism to acquire essential traits such as binocular vision, language, and social attachment. For decades, the closure of these windows was viewed as a permanent, irreversible maturation process, a dogma heavily influenced by the foundational work of David Hubel and Torsten Wiesel in the 1960s. Their landmark experiments on the visual cortex demonstrated that sensory deprivation during a specific early window permanently altered cortical architecture, leading to lifelong deficits that could not be corrected in adulthood.
However, recent breakthroughs across molecular neuroscience, neuropharmacology, and digital therapeutics have systematically dismantled this rigid framework. Current research reveals that the cessation of heightened developmental plasticity is not a passive decay of learning capacity, but rather an active, molecularly regulated process involving the deployment of structural "brakes" within the central nervous system. By identifying these brakes - ranging from perineuronal nets to myelin-associated growth inhibitors - scientists have unlocked the theoretical and practical potential to safely reopen neuroplastic windows in adulthood. This paradigm shift holds unprecedented clinical implications for neurodevelopmental disorders such as autism spectrum disorder and schizophrenia, as well as for the rehabilitation of acquired brain injuries and strokes. This report provides an exhaustive analysis of the molecular mechanisms dictating plasticity, reevaluates classical linguistic hypotheses in diverse global populations, and synthesizes 2023 - 2025 breakthroughs in pharmacological and non-pharmacological interventions capable of rejuvenating adult neuroplasticity.
1. Conceptual Framework: Critical Periods versus Sensitive Periods
To evaluate the trajectory of lifespan neuroplasticity, it is essential to delineate the precise distinction between "critical periods" and "sensitive periods." While these terms are frequently conflated in early literature, contemporary neuroscience treats them as functionally distinct phenomena with different degrees of temporal rigidity and residual adult plasticity. Understanding this dichotomy is paramount for designing therapeutic interventions, as the mechanisms required to reopen a strictly closed circuit differ from those needed to simply enhance a resistant one.
The Critical Period: Strict Temporal Constraints
A critical period is defined as a bounded developmental epoch during which the nervous system is absolutely dependent on specific environmental stimuli to develop a particular functional circuit 123. If the requisite experience is withheld during this strictly defined window, the neural architecture fails to organize correctly, resulting in permanent, irreversible deficits 4. The classic, experimentally verified paradigm for a strict critical period is the development of binocular vision and ocular dominance columns in the primary visual cortex (V1) 456. As established by the aforementioned work of Wiesel and Hubel, if one eye is deprived of patterned light during a specific postnatal window, the cortical territory normally allocated to that eye is permanently co-opted by the open eye, leading to amblyopia 57. In a strict critical period, learning or structural recovery outside the window is fundamentally impossible under normal physiological conditions because the molecular mechanisms actively prohibit further rewiring 18.
This rigid constraint applies not only to sensory processing but also to non-verbal modalities. Neuroimaging studies utilizing functional magnetic resonance imaging (fMRI) have demonstrated that individuals who acquire American Sign Language (ASL) from birth exhibit distinct patterns of right-hemisphere brain activation compared to those who learn ASL after puberty 910. Despite achieving high fluency, late learners of ASL exhibit persistent subtle grammatical errors, particularly with verb signs of motion, and significantly less right angular gyrus activation 10. This confirms that the brain's requirement for early exposure to organize foundational circuitry extends across diverse cognitive and sensory domains, enforcing a strict "use it or lose it" neurological policy 910.
The Sensitive Period: Optimal Windows and Residual Adult Neuroplasticity
Conversely, a sensitive period represents a phase of heightened susceptibility to environmental influences, but lacks the absolute rigidity and definitive closure of a critical period 911. During a sensitive period, experience exerts a maximal, highly efficient effect on brain circuitry, allowing for rapid, almost effortless acquisition of skills 912. However, if the stimulus is absent during this peak phase, the individual may still acquire the skill later in life, albeit requiring substantially more conscious effort, explicit instruction, and time, typically resulting in a lower ultimate level of proficiency 111313.
Second language acquisition serves as the premier example of a sensitive period 131415. While infants absorb phonetic and syntactic rules effortlessly from mere exposure, adults can still learn a second language and, in select cases with extensive immersion, achieve near-native grammatical competence and vocabulary 1113. The presence of this residual adult neuroplasticity indicates that the molecular brakes applied at the end of a sensitive period do not entirely abolish synaptic remodeling. Instead, they elevate the threshold for plasticity, demanding stronger, more salient, or highly repetitive stimuli to enact structural changes in the neural network 1113. This residual capacity is exactly what therapeutic interventions aim to amplify, seeking to lower the activation energy required to forge new neural pathways in the mature brain.
2. Molecular Mechanisms Dictating Plasticity Onset and Closure
The transition from a highly plastic developmental state to a stabilized, mature neural network is governed by an intricate interplay of cell-autonomous and non-cell-autonomous molecular signals. The temporal onset and eventual closure of plasticity windows are actively regulated by specific neurochemical pathways, which operate sequentially to first trigger network malleability and subsequently lock it into place.
Excitatory/Inhibitory (E/I) Balance and GABAergic Maturation
The onset of critical period plasticity is universally preceded by a necessary shift in the excitatory/inhibitory (E/I) balance within the cortex. Specifically, the maturation of fast-spiking, parvalbumin-expressing (PV+) GABAergic interneurons is the primary trigger for critical period opening 51617. Prior to PV cell maturation, developing cortical networks are overwhelmingly excitatory and lack the precise spatiotemporal resolution required to encode complex sensory or cognitive inputs 718.
As local circuit inhibition gradually increases during early development, it reaches a specific threshold that effectively opens the plasticity window 16. Experimental manipulations in animal models have robustly confirmed this mechanism: premature pharmacological activation of GABA-A receptors using benzodiazepines (such as diazepam) artificially accelerates the onset of critical periods 1620. Conversely, the genetic deletion of Gad65 - the enzyme responsible for synthesizing the inhibitory neurotransmitter GABA - prevents the normal maturation of these inhibitory circuits, thereby permanently delaying the opening of the visual critical period 1620. Thus, the precise calibration of the E/I balance acts as the initial gateway for experience-dependent neurodevelopment.
Growth Factors: The BDNF-TrkB Axis
The maturation of these PV+ GABAergic networks does not occur in a vacuum; it is heavily reliant on the presence of specific growth factors, most prominently Brain-Derived Neurotrophic Factor (BDNF) 1719. BDNF acts as a crucial intercellular signaling molecule, effectively translating experience-driven pyramidal neuron activity into the development and refinement of inhibitory interneurons. It is secreted and subsequently binds to its canonical TrkB receptor on PV cells, initiating an intracellular cascade of early/immediate genes that promotes their functional and morphological maturation 171920.
Transgenic models involving the specific overexpression of BDNF in the visual cortex result in an accelerated, premature onset and subsequent early closure of the ocular dominance critical period, fundamentally shifting the developmental timeline 1920. Interestingly, the transition to a fully mature state requires achieving a finely tuned synaptic balance. A disruption in BDNF expression alters this trajectory, maintaining a state of synaptic imbalance that leaves developing neurons excessively vulnerable to external stressors and incapable of consolidating learned information 7.
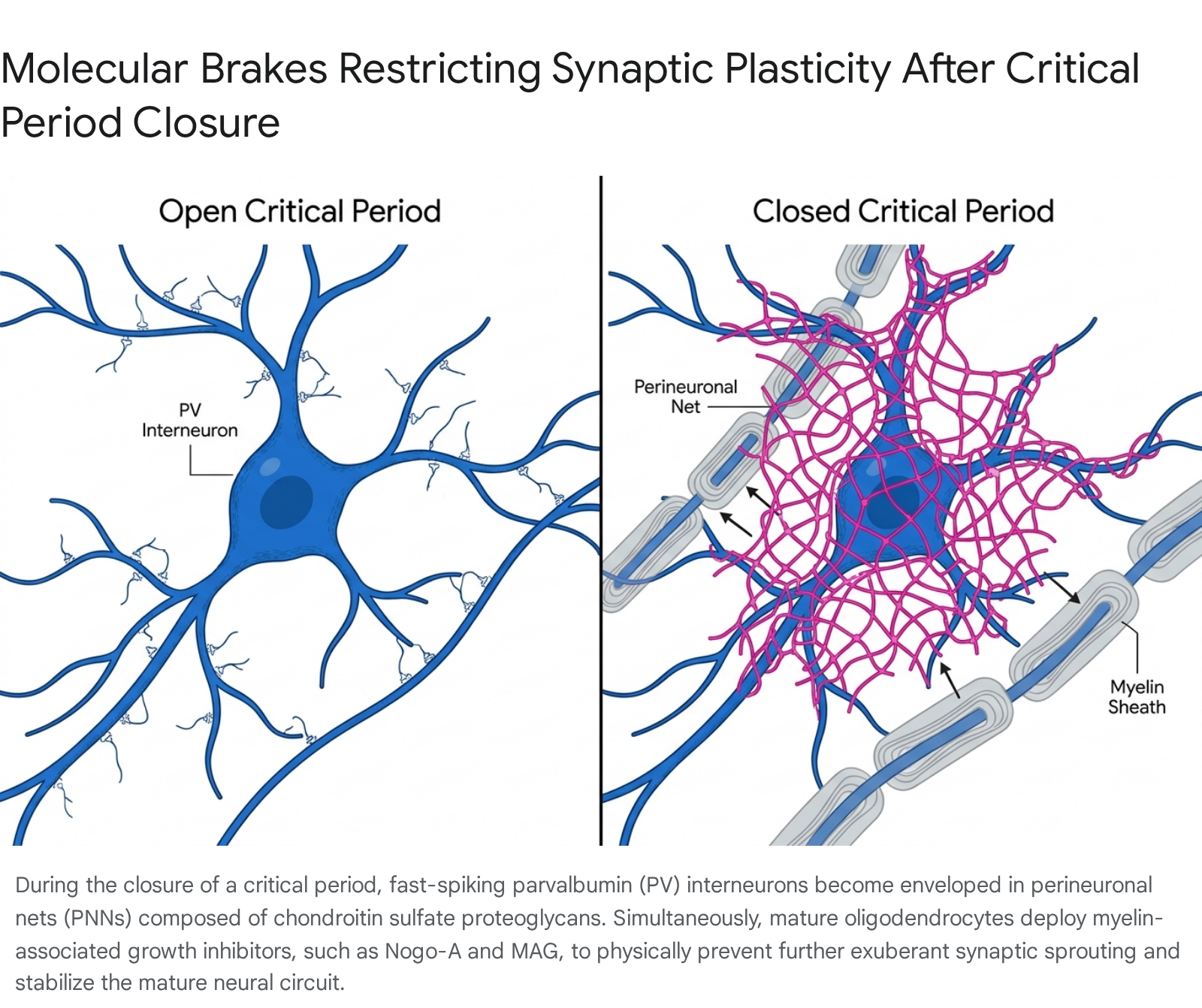
Extracellular Brakes: Perineuronal Nets (PNNs)
As the critical period begins to close, the brain transitions from a state of dynamic plasticity to one of structural stability. This stabilization is achieved through the active construction of the extracellular matrix (ECM), specifically the formation of perineuronal nets (PNNs) 816. PNNs are specialized, cartilage-like mesh structures composed primarily of chondroitin sulfate proteoglycans (CSPGs), tenascin-R, and other extracellular matrix components 1720. These nets physically encase the soma and proximal dendrites of mature PV+ interneurons 1618.
Functionally, PNNs act as a molecular "brake" on the system. They provide neuroprotection against oxidative damage induced by the high metabolic demands of fast-spiking activity, but more importantly, they physically prevent the formation of new synaptic connections and restrict the lateral mobility of surface receptors 161718. The accumulation of PNNs is strongly correlated with the termination of sensory and cognitive plasticity. In experimental models, the enzymatic degradation of CSPGs via the direct application of chondroitinase-ABC successfully dissolves PNNs. This intervention effectively reopens visual plasticity in adult subjects and has been shown to reinstate the capacity for fear memory extinction in the adult basolateral amygdala, proving that the physical removal of the ECM restores juvenile-like malleability 816.
Glial Contributions and Myelin-Associated Growth Inhibitors
Parallel to the development of perineuronal nets, the proliferation and maturation of macroglia - specifically astrocytes and oligodendrocytes - further restrict adult neuroplasticity 58. The timing of astrocyte maturation, mediated by factors such as connexin 30 (Cx30), is closely linked to the maturation of inhibitory circuits; genetic knockdowns of astrocytic Cx30 have been shown to delay PV neuron maturation and thereby keep ocular dominance plasticity open in adult mice 5.
Simultaneously, oligodendrocytes produce myelin to insulate axons, enhancing the speed and efficiency of action potential propagation across the central nervous system. However, mature myelin contains potent growth-inhibitory proteins, including Nogo-A, Myelin-Associated Glycoprotein (MAG), and Oligodendrocyte Myelin Glycoprotein (OMgp) 5. These molecules bind to the specific Nogo-66 receptor on nearby neurons, actively initiating signaling cascades that collapse growth cones, thereby inhibiting axonal sprouting and large-scale synaptic reorganization 58. The dramatic, developmental increase in MAG and Nogo-A abundance tightly mirrors the closure of visual critical periods 58. Blocking oligodendrocyte maturation or pharmacologically neutralizing these myelin-associated inhibitors has been shown to extend ocular dominance plasticity well beyond its typical developmental closure, highlighting myelination not merely as an insulator, but as a dominant structural inhibitor of lifelong learning 5.
3. Human Developmental Domains: A Comparative Timeline
The timing of plasticity onset, peak efficiency, and eventual closure varies drastically depending on the specific functional domain. This staggered timeline reflects an evolutionary imperative: lower-order sensory systems must mature rapidly to ensure immediate survival and accurate environmental processing, whereas higher-order cognitive and social networks exhibit protracted development, remaining malleable into late adolescence and early adulthood to allow for complex cultural assimilation 9212223.
Visual milestones, for instance, begin almost immediately after birth. Binocular functions such as fusion, stereopsis, and stereoacuity emerge abruptly around three to four months of age 2123. This rapid onset coincides with the development of layer 2/3 horizontal connections in the primary visual cortex, establishing the critical period for binocularity 2123. During the first two years of life, the brain is most susceptible to abnormal visual input; asymmetrical input during this phase can lead to irreversible amblyopia if not aggressively treated before age five, marking the steep closure of this window 21.
Conversely, language and social attachment operate on much longer trajectories. The foundational blocks of language, such as phoneme discrimination, begin in utero. Infants can differentiate between dialects of all global languages up until about four months of age 924. Following this peak, a rigorous process of synaptic pruning eliminates the perception of non-native sound contrasts by the first birthday, an elegant demonstration of the "use it or lose it" principle dictating early sensitive periods 9. Syntax and grammar acquisition, however, remain open far longer, correlating with the slower maturation of the prefrontal cortex 4. Similarly, the neurobiological windows governing social attachment and reward, driven heavily by oxytocin signaling, remain somewhat malleable through juvenile development and into late adolescence, preparing the individual for complex adult social hierarchies 325.
| Developmental Domain | Onset Age | Peak Plasticity Window | Closure / Functional Decline Phase | Neurobiological Markers & Notes |
|---|---|---|---|---|
| Binocular Vision & Stereopsis | ~3 - 4 months | 1 - 2 years | 5 - 7 years | Development of layer 2/3 horizontal connections in V1; high susceptibility to amblyopia if visual input is asymmetrical. Binocular summation matures rapidly. 212223 |
| Phoneme Discrimination | In utero / Birth | 6 - 8 months | 10 - 12 months | Infants distinguish all global phonemes initially; neural pruning eliminates non-native sound contrast perception by the first birthday ("use it or lose it"). 924 |
| Syntax Acquisition (Language) | ~12 - 24 months | 2 - 10 years | ~17.4 years (Steady decline thereafter) | Correlates with prefrontal cortex development and working memory expansion. Both "easy" and "difficult" syntax follow identical offset curves. 1126 |
| Social Attachment & Reward | Birth | Infancy / Juvenile Phase | Late Adolescence / Early Adulthood | Driven by oxytocin-mediated long-term depression (LTD) in the nucleus accumbens. Disruption relates to attachment and social deficits. 325 |
4. Language Acquisition: Reevaluating the Lenneberg Hypothesis
The study of language acquisition represents the most intensely debated frontier of critical period research. In 1967, Eric Lenneberg popularized the Critical Period Hypothesis for language in his seminal work Biological Foundations of Language, proposing that natural, native-like language acquisition ends abruptly at puberty due to the completion of brain lateralization and the biologically programmed loss of neuroplasticity 426. Decades of subsequent research have both supported and challenged the strict boundaries of Lenneberg's proposition, suggesting that language acquisition is governed by sensitive periods rather than a singular, unforgiving critical period 41424.
Large-Scale Insights: The ELSD Computational Model
A major historical hurdle in resolving the Lenneberg debate has been the reliance on underpowered studies and the conceptual difficulty of measuring ultimate attainment in learners who started at vastly different ages. Recent computational analyses of unprecedented scale have provided the first direct empirical estimates of how language-learning ability changes across the human lifespan. Utilizing a massive dataset of 669,498 native and non-native English speakers, researchers deployed an advanced computational model to disentangle current age, age at first exposure, and total years of immersive experience 26.
The findings fundamentally disrupt the classical view: the ability to acquire syntax to a native-like level does not abruptly terminate at puberty, as Lenneberg hypothesized. Instead, the capacity for optimal grammar-learning is preserved almost to the crux of adulthood, precisely ending at approximately 17.4 years of age, after which an inevitable, steady, and continuous decline occurs 26. Crucially, this age of offset applies uniformly to both "difficult" syntactic phenomena and "easy" phenomena that are typically mastered early in first-language acquisition 26. Furthermore, the data revealed that both native and non-native learners require approximately 30 years to reach asymptotic performance levels, fundamentally redefining the temporal scale of language acquisition 26.
Beyond WEIRD Populations: Linguistic and Geographic Diversity
A persistent limitation in developmental psychology and linguistics is the overreliance on Western, Educated, Industrialized, Rich, and Democratic (WEIRD) populations, predominantly analyzing the acquisition of Indo-European languages 27. Expanding research to linguistically diverse, non-WEIRD populations provides vital context on how varied socio-linguistic environments shape the parameters of the critical period.
In urban and rural African contexts, such as Ghana and Nigeria, early and sustained multilingualism is the norm rather than an academic exception. Infants in Accra, for example, routinely hear up to six distinct indigenous Ghanaian languages (predominantly from the Niger-Congo family, the largest linguistic family with 1554 languages) by just 12 months of age 27. African Indigenous educational systems have historically emphasized community-based, experiential language acquisition over formalized, segregated schooling 28. This challenges Western pedagogical frameworks by demonstrating that early, organic exposure to dense, multi-linguistic environments enhances overall communicative plasticity. In these contexts, children navigate multiple syntactical structures simultaneously, blurring the traditional boundaries between "first" and "second" language acquisition and suggesting that continuous exposure to linguistic diversity extends the sensitive period's functional utility 2728.
Tonal Language Acquisition and Perceptual Plasticity
Tonal languages, which dominate East Asia and Southeast Asia (such as Mandarin, Thai, and Hokkien), offer a unique lens through which to view perceptual neuroplasticity. Unlike non-tonal languages, where pitch is utilized solely to convey prosody or emotion, tonal languages use precise pitch variations (tonemes) to distinguish foundational lexical meaning 2930. Classical theories assumed that adults past the early critical period for phoneme discrimination would be biologically unable to accurately perceive and acquire novel tonemes 2931.
However, recent studies indicate significant, enduring residual plasticity for adult tone acquisition. American adults with absolutely no prior exposure to tonal languages demonstrated the ability to discriminate Mandarin tonemes at above-chance levels, achieving near-native perceptual proficiency after minimal computerized or classroom training 2931. Furthermore, research on L1 (first language) transfer reveals that learners originating from diverse tonal backgrounds process new tonal languages differently, leveraging their existing neural architecture. For example, Thai learners (who natively speak a tonal language) exhibit superior accuracy in Mandarin tone acquisition compared to Bangladeshi learners (whose native language is non-tonal) 3032. This emphasizes that the early establishment of a tonal neural network facilitates the assimilation of subsequent tonal parameters in adulthood, a concept closely aligned with the Speech Learning Model (SLM) 30.
Additionally, studies on individuals exposed to Hokkien exclusively during early caregiving, but who subsequently abandoned it and adopted English as their primary language, show that traces of Hokkien tonal phonology remain preserved in their implicit memory 33. When tested alongside peers with no such early exposure, the former group exhibited a distinct advantage in re-acquiring Hokkien tonemes. This demonstrates that early exposure lays a latent neuroplastic foundation that bypasses later critical period closures, waiting dormant until reactivated 33.
5. Pharmacological Reopening of Critical Periods: The Psychedelic Renaissance
One of the most consequential paradigm shifts in molecular neuroscience between 2023 and 2025 has been the discovery that specific pharmacological agents - specifically classical and atypical psychedelics - possess the profound, unprecedented ability to reopen critical periods in the adult brain 343536. Historically used in therapeutic contexts for post-traumatic stress disorder (PTSD), severe depression, and addiction, the unifying mechanism underlying the profound efficacy of these compounds remained elusive until recent mapping of the social reward circuitry in animal models.
MDMA, Oxytocin, and Social Reward Learning
Groundbreaking research led by neuroscientist Gül Dölen revealed that MDMA (an atypical psychedelic and empathogen) successfully reopens the critical period for social reward learning in adult mice 3253435. During normal juvenile development, animals possess a highly malleable window where they easily associate social interaction with reward, a necessary phase for establishing flocking, bonding, and mating behaviors. In adulthood, this capacity dramatically wanes as the critical period closes 2537.
However, Dölen's team demonstrated that a single dose of MDMA restores this juvenile neuroplastic state. It achieves this by stimulating a metaplastic upregulation of oxytocin-dependent long-term depression (LTD) within the nucleus accumbens, a core region of the brain's reward and reinforcement circuitry 33738. This MDMA-induced window of heightened social malleability relies heavily on the activation of oxytocin receptors 337. This molecular pathway directly explains why the drug generates powerful prosocial effects in humans and enhances the psychotherapist-patient bond during PTSD treatment: the drug fundamentally rewires the brain to value and integrate social support and emotional memory processing in a way typical only of early childhood 337.
Classical Psychedelics and Extracellular Matrix Remodeling
Initially, researchers assumed this reopening was a unique property of MDMA due to its explicit prosocial nature. However, subsequent studies systematically evaluated classical psychedelics (LSD, psilocybin), dissociatives (ketamine), and hallucinogenic stimulants (ibogaine) 2534. Stunningly, despite operating through entirely different primary neurotransmitter receptors (e.g., LSD and psilocybin act primarily via serotonin 5-HT2A receptors, unlike MDMA or ketamine), all these disparate substances act as "master keys" to reopen the social reward critical period 253436.
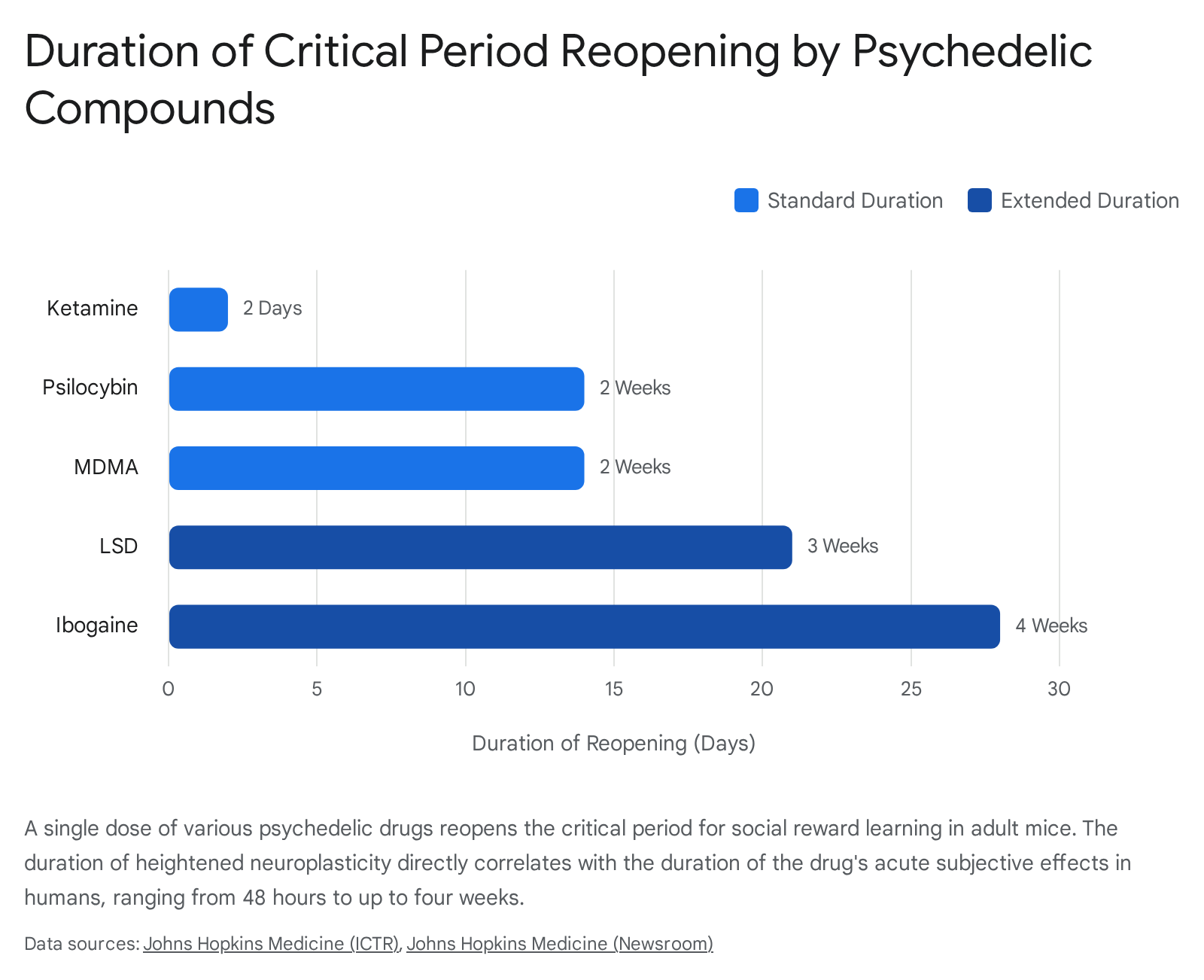
The defining variable among these drugs is the duration the critical period remains artificially open. The research uncovered a fascinating parallel: the length of the open state in mice is strictly proportional to the length of the drug's acute subjective effects (the "trip") in humans. Ketamine, a short-acting drug, maintains the open state for approximately 48 hours; psilocybin and MDMA for roughly two weeks; LSD for three weeks; and ibogaine, known for its multi-day acute effects, holds the window open for a full four weeks following a single administration 3436.
The unifying molecular mechanism driving this extended plasticity involves profound transcriptomic shifts. Psychedelic administration alters the expression of over 65 protein-producing genes within the cells of the nucleus accumbens 3436. Notably, 20% of these differentially expressed genes are directly responsible for regulating the extracellular matrix 3436. By signaling the brain to dissolve and restructure the restrictive scaffolding of the perineuronal nets, psychedelics effectively remove the structural brakes of adulthood, restoring a state of youthful synaptic flexibility and metaplasticity that allows therapeutic integration to physically rewire the brain 2539.
Epigenetic Modulation: HDAC Inhibitors
Beyond psychedelics, epigenetic modulators have demonstrated profound efficacy in reversing critical period closures for distinct sensory domains, further proving that developmental locks can be chemically picked. Valproate (VPA), a common anticonvulsant and mood stabilizer, also functions as a histone-deacetylase (HDAC) inhibitor, effectively removing "epigenetic brakes" on gene transcription and increasing neural plasticity 404344.
In highly controlled, double-blind human trials, adult men administered VPA successfully acquired the ability to identify absolute pitch - an extremely rare auditory skill (present in 0.01% of the population) traditionally impossible to learn after the age of seven due to early critical period closure 404344. By lifting the epigenetic brakes, VPA facilitated critical-period learning in the adult human brain, allowing the perceptual system to shift weight from relative to absolute pitch information 4041. This indicates that the temporary release of epigenetic repression allows adult networks to form novel, fundamental sensory maps previously restricted to early childhood 4441.
6. Non-Pharmacological Interventions for Rejuvenating Adult Neuroplasticity
While pharmacological agents alter the global biochemical milieu of the brain, non-pharmacological interventions utilize targeted electrical, magnetic, and immersive behavioral stimuli to induce highly localized, activity-dependent neuroplasticity.
Neuromodulation: TMS and tDCS
Non-invasive brain stimulation techniques, such as Transcranial Magnetic Stimulation (TMS) and transcranial Direct Current Stimulation (tDCS), exert their effects by physically modulating cortical excitability. Anodal tDCS, for example, promotes the depolarization of neuronal resting membrane potentials, thereby lowering the threshold for neuronal firing and facilitating long-term potentiation (LTP) and synaptic strengthening 4243.
While standalone stimulation often yields modest or mixed clinical outcomes, contemporary protocols pair continuous anodal stimulation (A-tDCS) or intermittent theta-burst stimulation (iTBS) over specific regions - such as the dorsolateral prefrontal cortex - with concurrent, targeted cognitive or behavioral therapy 424344. This synergistic approach forces specific, activity-dependent circuit remodeling by ensuring that the exact neural networks activated by the behavioral task are simultaneously receiving electrical facilitation, effectively mimicking the heightened excitatory state of a developing brain 4243.
Immersive Environments, Video Games, and Adult Amblyopia
The advent of digital therapeutics (DTx) - including augmented reality (AR), virtual reality (VR), and specialized therapeutic video games - has revolutionized the approach to reopening sensitive periods. These modalities are particularly effective for adult amblyopia, cognitive decline, and motor rehabilitation because they provide high-fidelity, high-repetition, and highly engaging stimuli that trigger the brain's endogenous reward networks (e.g., dopamine release), a powerful catalyst for neuroplasticity 454647.
Historically, adult amblyopia ("lazy eye") was considered untreatable because the critical period for binocular vision closes by age seven 212223. Traditional patching of the strong eye in adulthood yields minimal results. However, modern targeted behavioral therapy utilizing immersive video games has circumvented this. By using specialized VR headsets, therapists present slightly different, complementary visual inputs to each eye while the patient engages in a highly stimulating game. To succeed in the game, the adult brain is forced to suppress its learned preference for the dominant eye and integrate the inputs binocularly. The intense attentional requirement and reward-feedback loop of the game environment effectively reawaken dormant binocular circuitry, demonstrating that perceptual learning paradigms embedded in immersive media can dismantle the rigid perceptual suppressions formed in childhood.
In 2025, the FDA granted groundbreaking 510(k) clearance for biofeedback-integrated software development kits aimed at treating various conditions via over-the-counter video games, validating the medical power of digital therapeutics 45. Clinical applications like "Recovery Rapids," a motion-sensor video game for stroke survivors, force the utilization of paretic limbs in immersive environments. Patients simulate kayaking by performing gross arm motions, reaching, and steering in a virtual river. This fun, interactive environment ensures high patient adherence and facilitates motor cortex remodeling with only one-fifth of the conventional in-person therapist hours 48. Furthermore, VR training coupled with A-tDCS is actively being deployed to reverse cognitive impairment in multiple sclerosis by simultaneously lowering the threshold for neuronal firing while driving the network with intensive, task-specific exergame stimuli 42.
7. Clinical Implications: Neurodevelopmental and Acquired Brain Disorders
The realization that critical periods can be extended, delayed, or actively reopened provides a radically new conceptual framework for understanding the pathogenesis of severe mental illness and for treating a spectrum of acquired neurological conditions.
Neurodevelopmental Pathologies: Autism and Schizophrenia
Aberrations in the precise timing of critical period onset and closure are increasingly recognized as core pathophysiological mechanisms in severe psychiatric and neurodevelopmental disorders 549. The maturation of PV+ GABAergic interneurons - the cellular clock for critical periods - is frequently disrupted in these populations 549.
In Autism Spectrum Disorder (ASD), substantial evidence points to a precocious (premature) closure of sensory and social critical periods 516. This accelerated maturation limits the developing organism's time to accurately map complex environmental and social stimuli, resulting in the hyper-sensitivities, rigid behavioral patterns, and impaired social reward learning characteristic of the disorder 5735.
Conversely, Schizophrenia is characterized by the delayed or failed maturation of these inhibitory networks 549. In the absence of properly timed PV cell maturation and the subsequent failure to form protective perineuronal nets, the critical period remains aberrantly open. This leaves the cortical circuits in a prolonged state of synaptic instability and severe E/I imbalance 5749. This lack of structural stabilization perfectly correlates with the typical onset of disorganized thought processes, cognitive deficits, and auditory hallucinations in late adolescence, exactly when the prefrontal cortex should be finalizing its structural pruning.
Adult Brain Injury and Stroke Recovery
In the realm of adult neurorehabilitation, clinical doctrine has long held to the rigid concept of a "Golden Period" - a narrow 3 to 6-month window immediately following a stroke characterized by peak, injury-induced neuroplasticity, during which 60-80% of functional recovery occurs 5055. Historically, the perceived hard closure of this window justified the general cessation of intensive physical therapy for chronic patients, under the assumption that the brain was entirely "broken" and incapable of further repair 4450.
However, high-resolution analyses of post-stroke recovery trajectories have thoroughly challenged this dogma. Recent models analyzing individual patient data reveal an extended, smooth gradient of enhanced sensitivity to treatment that continues far beyond 12 months post-stroke, allowing improvements in body function even at late chronic stages 50. This lingering adult plasticity is currently being exploited through novel neurotechnologies.
The FDA-approved Vivistim Paired VNS System represents a major breakthrough, utilizing an implantable pulse generator to deliver vagus nerve stimulation precisely timed with active rehabilitative movements 51. By artificially triggering the release of neuromodulators during specific motor actions, paired VNS hacks the brain's plasticity pathways. In pivotal trials, chronic stroke patients (up to 20 years post-injury) receiving VNS achieved an average Upper Extremity Fugl-Meyer Assessment score increase of 5 points, generating two to three times more hand and arm function than standard therapy alone 4451. Advanced treatments, ranging from extended endovascular thrombectomy windows (up to 24 hours post-onset) to experimental intracerebral mesenchymal stem cell (MSC) transplantations (which locally secrete high levels of BDNF during critical post-stroke periods), further underscore that the adult brain retains a latent, robust capacity for profound structural repair if provided the correct molecular and behavioral keys 5152.
Conclusion
The traditional neuroscience paradigm, which characterized the adult brain as structurally rigid and developmentally finalized, has been permanently eclipsed by an understanding of dynamic, lifelong neuroplasticity. Critical and sensitive periods are not strictly dictated by chronological age alone, but are meticulously governed by a reversible biochemical architecture, including excitatory-inhibitory balance, growth factor signaling, and the restrictive physical scaffolding of the extracellular matrix and myelin inhibitors.
By deciphering the specific mechanisms that brake neuroplasticity - such as perineuronal nets and myelin-associated glycoproteins - the scientific community has initiated an era of therapeutic plasticity reopening. Psychedelic compounds have emerged as profound agents capable of dissolving these structural constraints, reopening learning windows strictly proportional to their acute subjective effects, and facilitating the rewiring of deep-seated trauma and social reward deficits. Simultaneously, targeted epigenetic modulators, non-invasive brain stimulation, and immersive digital therapeutics are proving highly effective at inducing localized, activity-dependent network repair in adults.
From extending the stroke recovery window decades past the initial injury, to utilizing video games to force binocular integration in adult amblyopia, to reevaluating the temporal boundaries of linguistic mastery across diverse global populations, the evidence indicates a profound shift. The human brain's capacity to learn, adapt, and heal is fundamentally preserved throughout the lifespan. Future translational efforts must focus on optimizing the synergistic application of pharmacological catalysts, neurostimulation, and rigorous, targeted behavioral conditioning to fully harness the immense therapeutic potential of the artificially opened critical period state.