Neurological effects of leadership and power
The phenomenon of leadership fundamentally alters the neurobiological landscape of the individual. When human beings transition into roles characterized by elevated power, status, and responsibility, their brains undergo measurable functional and structural adaptations. While leadership has traditionally been studied through the lenses of sociology, political science, and organizational psychology, modern neuroimaging, electroencephalography (EEG), and endocrinological assays have established that social hierarchies exert a profound, immediate, and continuous influence on neural plasticity, neurochemistry, and physiological stress responses. The contemporary evidence indicates that power operates as an active sculptor of the central nervous system, modifying the circuits responsible for empathy, risk evaluation, prosocial behavior, and threat detection 123.
Understanding the neuroscience of leadership requires moving beyond static trait theories of personality to examine how systemic authority alters information processing. This report synthesizes findings from cognitive neuroscience, social neuroendocrinology, and cultural neuroscience to delineate the precise mechanisms by which the acquisition and exercise of power alter human brain function.
Evolutionary Framework of Social Hierarchy
Human beings, as obligatorily interdependent social animals, inherently form hierarchical structures to solve the complex challenges of group coordination, conflict resolution, and resource allocation 245. These social structures are not arbitrary cultural artifacts but are deeply rooted in evolutionary biology, reflecting navigation constraints and the cognitive capacity required to manage complex dyadic relationships within groups 67.
Empirical evidence and mathematical modeling of hominin evolution suggest a complex historical trajectory regarding hierarchy. Theoretical models of social navigation constraints indicate that human social structures likely transitioned from hierarchical to egalitarian phases driven by encephalization over the last two million years, before reverting to heavily hierarchical structures due to rapid demographic expansions during the Neolithic period 67. The cross-cultural consistency of hierarchy formation in modern humans suggests that the brain possesses specialized, evolved mechanisms for deciphering, maintaining, and navigating social ranks instantaneously and automatically 27.
Dual Strategies of Rank Acquisition
Evolutionary anthropology and social neuroscience propose the Dual Strategies Theory, which posits that humans possess two distinct, evolved pathways for ascending social hierarchies: dominance and prestige 98119. These pathways differ substantially in their behavioral manifestations, evolutionary histories, and, critically, their underlying neurobiological and endocrinological profiles 51310.
Dominance represents an older, phylogenetically ancient mammalian strategy characterized by the acquisition of status through coercion, intimidation, and the threat of withholding resources 9119. In human populations, the dominance strategy is strongly correlated with hubristic pride, narcissistic tendencies, heightened assertiveness, and a measurable reduction in prosocial cooperative behaviors 4119. Biologically, dominance is governed by an underlying neurochemistry optimized for threat and conflict. It relies heavily on the amygdala for rapid threat detection and is hormonally characterized by elevated baseline testosterone combined with lowered serotonin availability 913. While dominance is effective for rapidly coordinating groups during existential crises, its coercive nature often creates instability, as subordinates may coordinate to suppress or overthrow the dominant individual through reverse dominance mechanisms 6119.
Prestige, by contrast, is uniquely human. It represents a social innovation wherein status is freely conferred by group members in exchange for the leader's expertise, competence, and wisdom 9119. The prestige pathway operates as a social contract based on voluntary emulation and social learning. Individuals utilizing prestige strategies exhibit authentic pride, heightened cooperativeness, and an absence of toxic organizational behaviors 411. The neural architecture of prestige involves the medial prefrontal cortex (mPFC) and the nucleus accumbens, which facilitate affiliation and reward processing 13. Furthermore, prestige relies heavily on the dorsal raphe nucleus (DRN) and is regulated by serotonin transporter availability, serving to maintain social calculation, empathy, and impulse inhibition over extended periods 913.
The divergence between these two systems underscores that leadership is not a monolithic biological state, but rather a flexible activation of distinct neural architectures based on the leader's environment, group dynamics, and psychological disposition.
| Neurobiological & Behavioral Trait | Dominance-Based Hierarchy | Prestige-Based Hierarchy |
|---|---|---|
| Primary Method of Acquisition | Coercion, intimidation, force, and fear 9119. | Freely conferred deference, skill demonstration, teaching 9119. |
| Evolutionary Origin | Ancient reptilian/mammalian competitive survival 9. | Uniquely human adaptation for cultural transmission and social learning 910. |
| Primary Neural Correlates | Amygdala (threat detection) 913. | Medial prefrontal cortex (mPFC), nucleus accumbens 13. |
| Endocrine Profile | High testosterone, low serotonin, low baseline cortisol 91310. | High serotonin transporter availability 913. |
| Associated Pride Type | Hubristic pride 119. | Authentic pride 119. |
| Subordinate Response | Fear, subordination, vigilance, low relational satisfaction 99. | Respect, admiration, voluntary emulation, high cooperation 499. |
The Empathy Deficit and Motor Resonance
As individuals accumulate social power, psychological and neuroscientific studies have documented a progressive deterioration in specific socio-cognitive capacities, most notably empathy and perspective-taking. This phenomenon, formalized as the "power paradox," outlines that the very qualities that often elevate individuals to leadership positions - empathy, fairness, generosity, and collaboration - are frequently the first cognitive capacities to erode once power is secured and exercised 3151112.
Functional Adaptation Versus Structural Damage
Public discourse has occasionally sensationalized these findings, operating under the dramatic metaphor that "power causes brain damage" 1315121813. While philosophical literature has long debated the corrupting influence of authority, empirical neuroscience specifies that this deficit is not the result of structural lesions, neurodegeneration, or literal cellular death. Rather, power induces a state of altered neuroplasticity - a functional shift in how the brain allocates metabolic and cognitive resources 11213.
Powerful individuals operate in environments saturated with asymmetric influence, heavy cognitive demands, and elevated responsibilities 2. This environment necessitates highly efficient filtering of stimuli. Consequently, the brain adaptively suppresses certain interpersonal monitoring mechanisms that are computationally expensive. Research indicates that maintaining deep, individualized empathetic connections with a vast number of subordinates is metabolically draining; thus, the brains of powerful individuals shift toward using heuristics and stereotypes to process social information rapidly 11512.
Suppression of the Mirror Neuron System
The physiological foundation of this empathy deficit has been localized to the human mirror neuron system (MNS). The MNS is a neural network that activates both when an individual performs an action and when they observe another individual performing the same action, serving as the neural basis for motor resonance, emotional contagion, and intent recognition 1415.
In transcranial magnetic stimulation (TMS) studies examining motor cortical output, researchers demonstrated that participants primed with feelings of high power exhibited a significant reduction in motor resonance when observing others' actions 151216. The suppression of this mirroring process in powerful individuals is not a conscious, deliberate choice but an involuntary physiological state. Even when explicitly instructed to attempt perspective-taking and actively empathize, subjects under the influence of experimentally induced power struggled to re-engage these mirroring circuits, suggesting a temporary but profound neurochemical blockade 15.
Advanced functional magnetic resonance imaging (fMRI) meta-analyses demonstrate that the MNS is anatomically bifurcated into social and non-social pathways, further explaining how power selective impairs specific types of empathy 17.
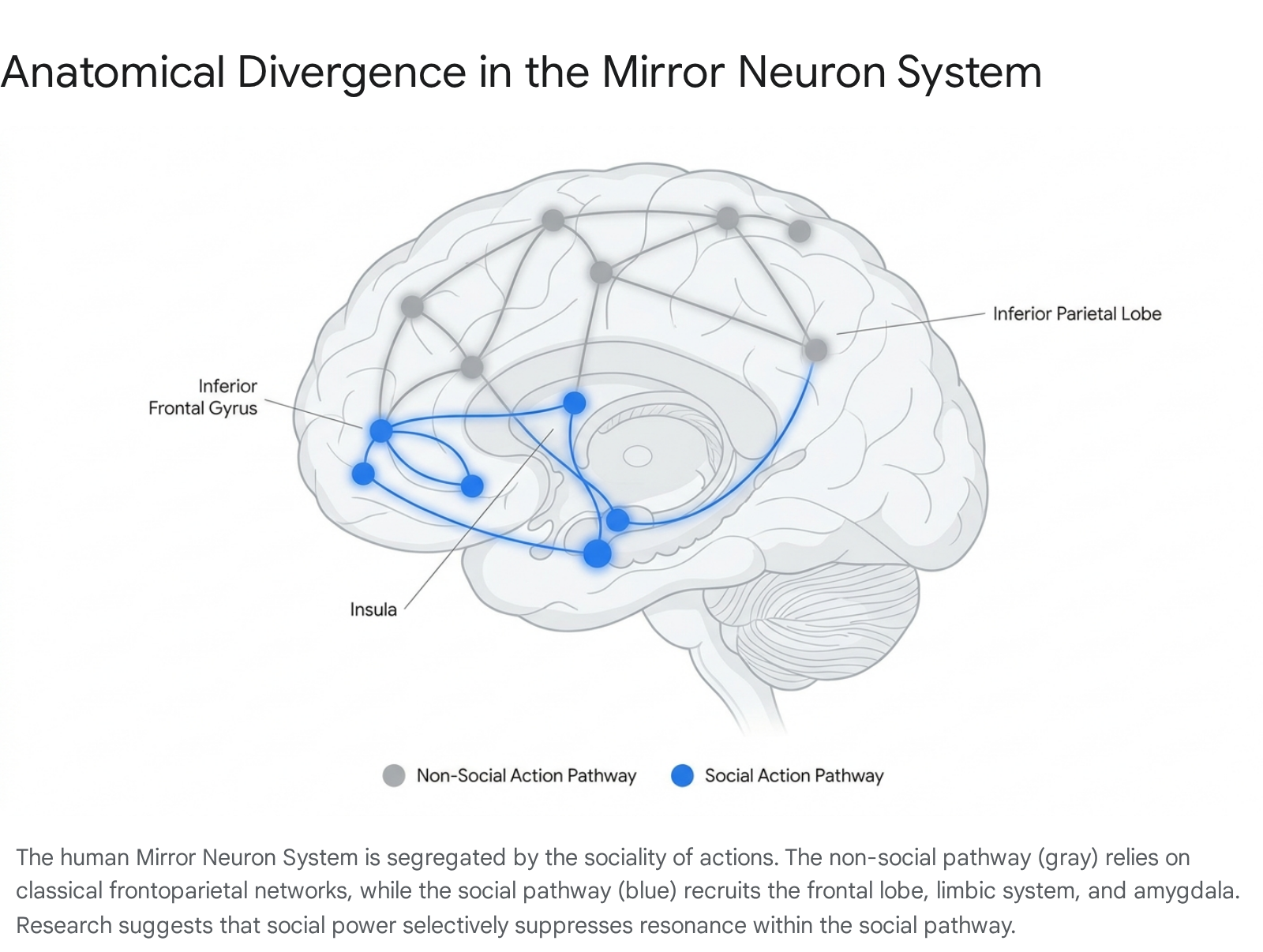
Non-social actions (e.g., grasping an inanimate object) recruit a classical frontoparietal network, specifically the bilateral inferior parietal lobe (IPL), superior parietal lobule (SPL), postcentral gyrus, and precuneus 17. Conversely, actions carrying social significance (e.g., communicative gestures, facial expressions of emotion) bypass the heavy reliance on the parietal lobe and instead activate the frontal lobe, limbic system, bilateral amygdala, right insula, and bilateral inferior frontal gyrus (IFG) 17.
Power specifically dampens the sensitivity of this social MN pathway. Because the brains of powerful individuals are less likely to automatically simulate the emotional states of subordinates via the IFG and insula, leaders become increasingly susceptible to treating subordinates instrumentally rather than empathetically 11318.
Endocrine Modulators of Executive Function
The intersection of leadership and neuroendocrinology reveals that the pursuit, acquisition, and maintenance of power are heavily mediated by fluctuations in systemic hormones and neurotransmitters. Leaders must continuously navigate uncertainty, calculate risk, and resolve interpersonal conflict, all of which are managed by specific endocrinological profiles.
Dopaminergic Regulation of Risk and Reward
Leaders, particularly entrepreneurs and corporate executives, are fundamentally tasked with decision-making under conditions of volatility and uncertainty. The neurobiology of entrepreneurial risk-taking and strategic shifting is heavily mediated by the dopaminergic system, specifically the mesolimbic pathway, which connects the ventral tegmental area (VTA) to the nucleus accumbens (NAcc) 2519.
Dopamine is frequently misunderstood strictly as a "pleasure" chemical; in the context of executive leadership, it functions primarily as a neuromodulator of anticipation and reward-prediction error 2519. When environmental rules switch from stable to volatile - a common scenario for organizational leaders - dopamine levels surge in the striatum. This release facilitates rapid adjustment to changing scenarios, allowing the leader to unlearn obsolete rules and relearn necessary action-outcome associations 1927.
However, neuroimaging and pharmacological interventions demonstrate that the relationship between dopamine and optimal decision-making follows an inverted U-shaped curve. A moderate release of dopamine allows leaders to exist in a zone of optimal sensitivity, wherein they can process negative feedback, adapt, and learn from mistakes without being overwhelmed 27. Conversely, excessive dopaminergic activity hyper-sensitizes the brain to the prospect of potential gains 19. This generates an "optimism bias," resulting in choice inflexibility, impulsive risk-taking, and a failure to adequately weigh the probabilities of massive losses.
Genetics also influence this leadership trait. Variations such as the DRD4 7-repeat allele have been associated with higher financial risk tolerance and lower aversive responses to uncertainty, effectively lowering the brain's natural braking system against risky ventures 19. In high-stakes environments, the prefrontal cortex (the rational brain calculating probabilities) is frequently engaged in a tug-of-war with the dopamine-rich emotional centers pushing for instant gains and thrill 25.
The Dual-Hormone Hypothesis
Testosterone is traditionally implicated in status-seeking behavior, social dominance, and the motivation to maintain rank. However, the influence of testosterone is highly context-dependent and heavily regulated by glucocorticoids, particularly cortisol. The Dual-Hormone Hypothesis posits that testosterone only successfully predicts dominant social behavior when basal cortisol levels are simultaneously low 1320.
Cortisol, the primary hormone released during the body's physiological stress response, acts as an inhibitory gatekeeper. When an individual experiences high psychological stress (high cortisol), the behavioral expression of testosterone is muted, leading to a suppression of status-seeking behaviors and risk aversion. Conversely, the combination of high testosterone and low cortisol creates an endocrine profile strongly associated with stable, effective leadership, proactive dominance, and emotional regulation, distinguishing it from reactive, fear-based aggression 1320.
The Stress-Buffering Hypothesis of Authority
Conventional wisdom dictates that as leaders ascend to more powerful positions, the burden of managing subordinates, stakeholders, and organizational crises results in chronic psychological stress. Consequently, leadership is frequently assumed to cause physiological wear-and-tear and an elevated neuro-burden 2122. However, extensive physiological analyses of real-world executives, military officers, and government officials completely invert this assumption.
Psychological Control and Cortisol Attenuation
Research measuring salivary cortisol and self-reported anxiety clearly demonstrates that leaders experience significantly lower levels of physiological stress compared to non-leaders 21222324. In robust empirical assessments, high-ranking leaders exhibit baseline cortisol levels up to 0.54 standard deviations lower than subordinates, a finding that persists after controlling for age, income, and education 22. Furthermore, this phenomenon displays a dose-response relationship: within leadership cohorts, those possessing greater authority and managing larger numbers of direct reports exhibit progressively lower cortisol levels and lower self-reported anxiety 2123.
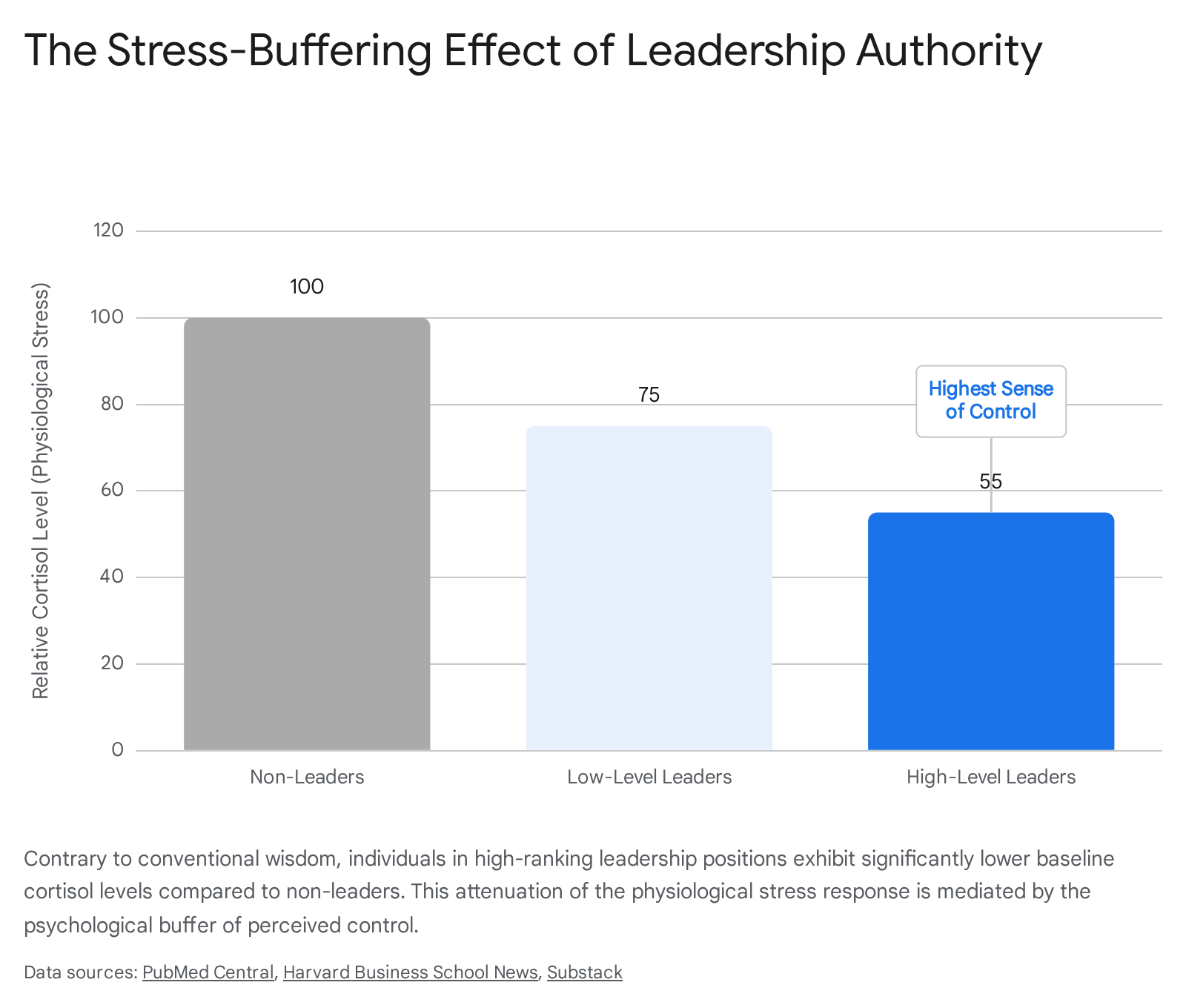
This counterintuitive reduction in the neuro-burden of responsibility is explained by the Stress-Buffering Hypothesis. The primary psychological mechanism mediating this attenuation of cortisol is the leader's heightened "sense of control" 212224. In human neurobiology, a stressor (e.g., an impending deadline, a financial shock) only triggers a severe hypothalamic-pituitary-adrenal (HPA) axis response if the organism perceives the threat as uncontrollable. Leaders possess greater autonomy over their schedules, the capacity to delegate threatening tasks, and the authority to shape their environments 224. This autonomy acts as a powerful psychological shield, preventing external demands from initiating a cascading neuroendocrine stress response.
The Impact of Hierarchical Instability
The physiological tranquility afforded by power is conditionally dependent on the stability of the social hierarchy. The stress-buffering effect rapidly dissipates, and can even reverse, when a leader's status is threatened or the organizational structure becomes unstable 2225.
Experimental paradigms subjecting leaders to social-evaluative stressors under unstable conditions (e.g., the threat of demotion or a fluctuating hierarchy) reveal significant spikes in both cortisol and testosterone reactivity 2225. In these volatile environments, the heightened testosterone - which normally facilitates decisive action when cortisol is low - transforms into a liability. It redirects cognitive resources toward hyper-vigilance, status defense, and aggressive posturing at the expense of strategic performance and competence 25. Thus, power acts as a neurobiological tranquilizer only when it is securely institutionalized; when contested, the loss of perceived control renders the leadership position a severe physiological stressor.
Neural Pathways of Prosocial Leadership
While power possesses the potential to suppress empathy, humans are uniquely capable of highly prosocial leadership - directing resources, effort, and strategic intent toward the betterment of the group at a direct cost to the self 262728. Advanced neuroimaging exposes that the brain does not utilize a single, generalized "reward" circuit for all positive outcomes; rather, selfish reward acquisition and prosocial resource distribution are governed by divergent neural systems.
Differentiating Selfish and Prosocial Networks
Conventional fMRI studies tracking reward processing (such as acquiring personal financial gain) reliably demonstrate activation in the core brain reward system. This encompasses the lentiform nucleus, caudate nucleus, thalamus, and parahippocampal gyrus 2629.
In stark contrast, decisions made to assist others activate highly specific socio-cognitive networks. Meta-analyses demonstrate that prosocial decision-making is conceptually clustered into three distinct behavioral facets: cooperation, equity, and altruism. While these behaviors share underlying neural commonalities, they maintain unique modular activations 29. The overarching prosocial network heavily recruits the dorsolateral prefrontal cortex (dlPFC) and the ventromedial prefrontal cortex (vmPFC). These regions are vital for integrating cognitive rule-sets with affective signals, allowing a leader to evaluate the moral and social utility of an action over immediate personal gratification 27.
Furthermore, leadership frequently requires the exertion of physical or cognitive effort specifically for the benefit of others, which yields a unique neural signature. During fMRI decision-making tasks, the anterior cingulate gyrus (ACCg) uniquely encodes the cost of prosocial effort 28. When an individual chooses to exert energy to secure a reward for a subordinate or peer, the ACCg exhibits robust multivariate representations of subjective value. Crucially, this specific ACCg pattern is entirely absent when individuals exert the exact same effort for personal, self-benefiting rewards 28. Stronger ACCg representation strongly correlates with higher self-reported affective empathy and an increased subsequent willingness to deploy force for the collective good.
Structural Brain Markers of Prosociality
The inclination toward prosocial leadership is not limited to transient functional activations; it is also reflected in the brain's resting-state structural and functional connectivity. Multimodal magnetic resonance imaging analyses of individuals engaging in high-stakes economic games reveal that consistent prosocial behavior correlates with macroscopic brain characteristics. These include a larger corpus callosum volume and stronger interhemispheric functional connectivity 30.
Moreover, regions comprising the broader "social brain network" - including the temporal lobe, temporoparietal junction (TPJ), insula, and inferior frontal gyrus (IFG) - display greater functional segregation and integration in highly prosocial individuals 30. Structurally, prosocial orientation is linked to fewer myelin maps combined with a thicker cortex in these areas. These physical biomarkers indicate that prosocial leadership requires extensive, high-bandwidth communication across bilateral brain hemispheres to continuously integrate complex social cues, moral constraints, and long-term planning 30.
| Brain Region / Network | Primary Activation Profile | Behavioral Correlate in Leadership |
|---|---|---|
| Lentiform & Caudate Nucleus | Personal Reward System 2629 | Self-interested decision making; immediate personal gain. |
| Anterior Cingulate Gyrus (ACCg) | Prosocial Effort Encoding 28 | Exerting physical or cognitive effort specifically to benefit others; high empathy. |
| vmPFC & dlPFC | Affective/Cognitive Integration 2729 | Evaluating equity, altruism, and the moral utility of resource distribution. |
| Temporoparietal Junction (TPJ) | Mentalizing / Theory of Mind 273031 | Inferring the emotional states, hidden needs, and intentions of group members. |
Cross-Cultural Variations in Authority Processing
The neurobiology of leadership does not operate in a vacuum; it is continuously modulated by the macro-level cultural frameworks within which a hierarchy is embedded. Cultural neuroscience demonstrates that societal norms regarding power distance - whether a society fundamentally values egalitarianism or steep vertical hierarchies - fundamentally re-wire how the brain perceives social status and processes interpersonal empathy 3132.
Social Dominance Orientation and Neural Reactivity
Individuals and cultures vary significantly in their Social Dominance Orientation (SDO), an established psychological metric that measures the preference for hierarchical social structures and the legitimization of inequality 73334. In individualist, egalitarian cultures (such as many Western democracies), neural responses to the suffering of others are generally more uniform, irrespective of the sufferer's social rank or group affiliation. However, in collectivist cultures with a high tolerance for power distance and strict social hierarchies, the brain's empathetic networks exhibit a strong ingroup and status-based bias 3134.
During cross-cultural fMRI studies where participants observed individuals experiencing emotional pain, those from hierarchical cultures exhibited significantly heightened activity in the left temporoparietal junction (L-TPJ) - a core node for mental state inference and perspective-taking - only when observing members of their own ingroup 3143. This preferential reactivity demonstrates that cultural conditioning regarding hierarchy and group boundaries actively dictates the allocation of neural resources for empathy.
Furthermore, an individual's specific SDO directly alters how they neurobiologically encode the faces of subordinates versus superiors. When highly dominant, hierarchy-preferring individuals view the faces of high-status individuals, they exhibit intense activation in the right anterior dorsolateral prefrontal cortex (aDLPFC) 7. Conversely, individuals with lower SDO scores (indicating egalitarian preferences) do not show this heightened aDLPFC response to status disparities 7. This establishes that the perception of leadership and submission is not universally hardwired but is a neuro-cultural synthesis, where ideological beliefs about power actively program the brain's visual and socio-cognitive processing centers.
Conclusion
The neuroscience of leadership reveals that power is far more than a sociological construct; it is a pervasive physiological state that dynamically alters the architecture of human cognition. The acquisition of authority fundamentally shifts the brain's operational priorities. It can adaptively suppress the mirror neuron system, creating an empathy deficit that biases leaders toward stereotyping, while simultaneously triggering dopaminergic surges that alter risk assessment and strategy generation.
However, the exact nature of this neural transformation depends heavily on the pathway to power and the stability of the environment. Dominance strategies entrench the brain in amygdala-driven threat responses and steep hormonal volatility. Conversely, prestige-based leadership leverages the medial prefrontal cortex and serotonin networks to foster prosociality and complex social learning. Furthermore, an institutionalized sense of control effectively insulates the leader from the debilitating physiological effects of chronic cortisol, providing a biological advantage. Ultimately, effective leadership requires a neurobiological balancing act: maintaining the dopaminergic drive and psychological autonomy required to execute high-stakes decisions, while actively overriding the brain's structural tendency to shut down the empathetic networks that bind a leader to their followers.