Neurobiology of music and brain plasticity
The intersection of auditory cognition and neurobiology has evolved over the past several decades from rudimentary localization studies into a highly sophisticated exploration of large-scale, dynamic brain networks. Foundational theoretical frameworks established by researchers such as Daniel Levitin and Robert Zatorre catalyzed a paradigm shift, definitively demonstrating that the human brain does not possess a singular, localized "music center." Instead, engaging with music recruits a widely distributed, highly interactive, and bilateral neural architecture that simultaneously encompasses sensory processing, motor planning, executive functioning, and the mesolimbic reward circuitry. In recent years, the deployment of advanced, high-resolution neuroimaging modalities - including functional magnetic resonance imaging (fMRI), diffusion-weighted imaging (DWI), and functional near-infrared spectroscopy (fNIRS) - has provided unprecedented spatial and temporal resolution, allowing researchers to elucidate the precise mechanisms underlying music-induced neuroplasticity.
This exhaustive analysis evaluates the profound manner in which music physically and functionally alters the brain. By prioritizing empirical data from recent (2023 - 2026) neuroimaging studies and rigorous clinical meta-analyses, this report delineates the stark neurobiological dichotomy between active musical performance and passive listening. It meticulously examines the profound structural neuroplasticity observed in trained musicians, the modulation of the default mode network (DMN) via the dopaminergic reward system in affective disorders, and the intense debate within cross-cultural neuroscience regarding the universality versus cultural mediation of musical syntax. Furthermore, it critically evaluates the empirical efficacy of neurologic music therapy (NMT) in treating major depression, Parkinson's disease, and dementia. In doing so, it strictly delineates verified biological mechanisms from social confounds, while systematically dismantling pervasive, oversimplified neuromyths that have historically plagued public understanding of brain function.
Deconstructing Neuromyths: Lateralization and the "Mozart Effect"
Before examining the complex neural circuitry activated by music, it is absolutely critical to dismantle the persistent misconceptions that permeate popular psychology and educational literature. Perhaps the most pervasive and resilient of these fallacies is the strict functional lateralization of the human cortex - commonly referred to as the "left-brain versus right-brain" myth. This oversimplification posits that logical, analytical, and mathematical processing occurs exclusively in the left hemisphere, while creative endeavors, intuitive leaps, and artistic processing, including music, are entirely relegated to the right hemisphere. Modern neuroimaging unequivocally and overwhelmingly refutes this model of global lateralization 12.
Massive resting-state functional connectivity magnetic resonance imaging (rs-fcMRI) studies, including investigations involving over 1,000 subjects whose brains were parceled into more than 7,000 distinct functional regions, have demonstrated absolutely no evidence of overall hemispheric dominance dictating personality types or broad cognitive styles 13. While it is a well-established neuroanatomical fact that specific, highly specialized local processes display varying degrees of lateralization - such as language syntax generally favoring the left hemisphere's perisylvian network, and pitch contour or spatial perception demonstrating a right-hemispheric advantage - complex cognitive behaviors like music processing require continuous, dense interhemispheric integration 347.
In fact, the neurobiological reality of musical creativity directly contradicts the popular lateralization myth. A seminal high-density electroencephalogram (EEG) study focusing on professional jazz guitarists undergoing complex improvisational tasks revealed a counterintuitive functional shift based on expertise. The study demonstrated that highly expert, technically proficient improvisational performances were characterized by predominant neural activity in the left hemisphere. Conversely, less experienced musicians, who were still learning the fundamental structures of jazz, relied more heavily on right-hemispheric activation 5. This empirical finding illustrates that years of rigorous practice and sensorimotor consolidation shift musical execution from a novel, exploratory, and highly effortful process (which heavily taxes right-lateralized attentional and spatial networks) to a highly structured, analytical, and automated syntactic process governed by left-lateralized executive and motor networks 5. Thus, musical creativity is not a geographically isolated right-brain phenomenon, but rather a dynamic, whole-brain capacity dependent upon robust functional integration across the entire cortex, facilitated heavily by the corpus callosum.
Similarly, the widely publicized "Mozart Effect" - the enduring claim that passive listening to classical music induces permanent structural enhancements in the brain and long-term improvements in spatial-temporal intelligence - must be rigorously discarded 6. Subsequent replication attempts and comprehensive neurobiological reviews have demonstrated that any transient cognitive enhancement observed following music listening is merely a byproduct of general physiological arousal and mood state optimization. This transient boost is mediated by the autonomic nervous system and temporary dopaminergic release, rather than any permanent alteration of cortical architecture or actual intelligence quotient (IQ) enhancement. Enduring cognitive transfer and verifiable structural neuroplasticity require the rigorous, long-term sensorimotor integration, sustained attention, and error-correction demands intrinsic to active musical training. The distinction between the passive reception of acoustic stimuli and the active generation of music is fundamental to understanding music-driven neuroplasticity.
Active Musical Performance Versus Passive Listening: A Network-Level Analysis
The neural requirements for passively perceiving a musical piece versus actively producing one represent two fundamentally distinct, albeit partially overlapping, dynamic network configurations. It is crucial to note that passive music listening is by no means a neurologically passive or dormant event; it demands the rapid, preattentive extraction of complex acoustic features, syntactic integration, and continuous predictive coding to anticipate harmonic and rhythmic resolutions. However, active performance acts as a massive multiplier for cortical engagement.
Meta-analyses of fMRI data utilizing seed-based $d$ mapping demonstrate that passive music listening bilaterally coactivates the ventral attention, somatomotor, default, dorsal attention, frontoparietal, and limbic networks ($p < 0.01$, family-wise error corrected) 789. The continuous, structured flow of acoustic information necessitates real-time auditory processing primarily in Heschl's gyrus and the superior temporal sulcus. This sensory extraction is continuously combined with temporal predictions generated by the supplementary motor area (SMA) and the cerebellum, even when the listener is entirely motionless 1011.
Interestingly, the specific pattern of functional connectivity established during passive listening is highly dependent on the subject's preexisting musical expertise, current cognitive load, and attentional state. Using advanced dynamic functional connectivity analyses such as Leading Eigenvector Dynamics Analysis (LEiDA), recent studies have mapped the transition of brain states from moment to moment during naturalistic music listening. These investigations have shown that trained musicians frequently and preferentially engage a highly integrated frontoparietal and salience network state. This state heavily involves the superior frontal gyrus (SFG), middle frontal gyrus (MFG), anterior cingulate cortex (ACC), and orbitofrontal cortex (OFC) 1213. This pattern reflects an analytical, reward-driven integration of complex auditory stimuli, wherein the musician is constantly evaluating the syntactic and structural components of the piece. Conversely, non-musicians exhibit a significantly higher probability of transitioning into, and remaining within, the Default Mode Network (DMN). This DMN dominance during listening reflects a more passive, associative state characterized by self-referential mental processes, internal mind-wandering, and autobiographical memory retrieval triggered by the emotional valence of the auditory stimulus 1213.
Active musical performance, whether singing or playing an instrument, radically amplifies this network engagement by enforcing simultaneous multimodal processing and continuous error-correction loops. An active performer must translate abstract visual symbols from sheet music, or retrieve complex sequences from working memory, and convert them into highly precise, sequenced motor commands. Simultaneously, the musician must continuously monitor the resulting auditory and somatosensory feedback to adjust micro-parameters such as pitch, velocity, and timing in milliseconds.
Comparative independent component (IC) analyses of fMRI data reveal that while both passive and active states recruit auditory and temporoparietal language networks, active performance uniquely and heavily recruits the left dorsolateral prefrontal cortex (DLPFC), the anterior cingulate cortex (ACC), and extensive, bilateral sensorimotor networks 1415.
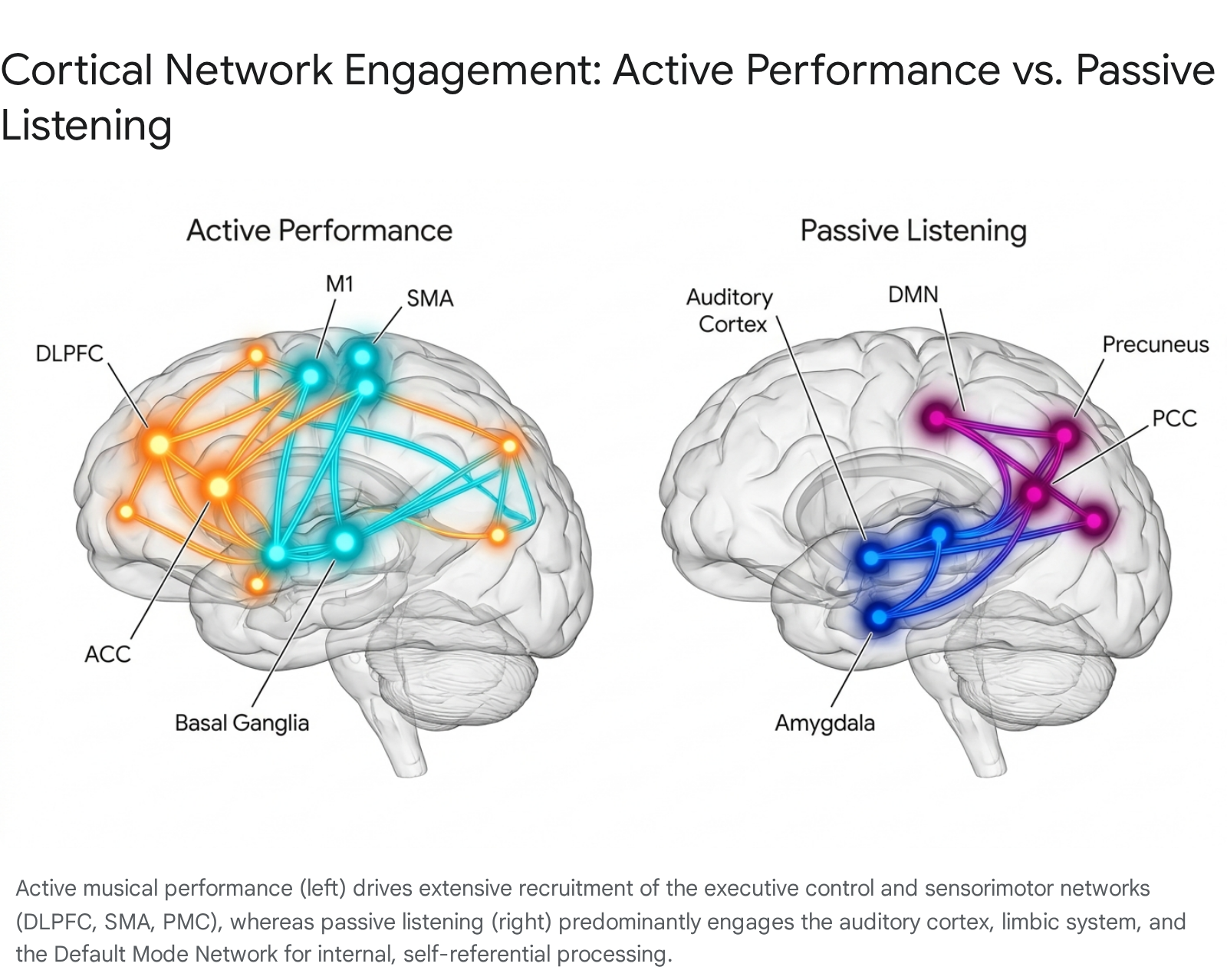
Active rhythm production enforces a tight cortical auditory-motor coupling that spans the basal ganglia, SMA, premotor cortex (PMC), and inferior parietal regions 10. The brain must continuously compute the difference between the intended acoustic output and the actual acoustic feedback, utilizing the cerebellum as a forward-model predictor to adjust motor commands before the next note is even struck.
| Brain Network / Region | Neurobiological Function in Passive Music Listening | Neurobiological Function in Active Musical Performance |
|---|---|---|
| Auditory Cortex (Heschl's Gyrus, STG) | Feature extraction (pitch, timbre, spectral density), contour analysis, and preattentive syntactic processing 41216. | Real-time auditory feedback monitoring, active pitch-matching, and acoustic error detection for immediate motor correction 1617. |
| Sensorimotor Network (SMA, PMC, M1) | Covert beat simulation and temporal prediction; often exhibits negative connectivity (suppression) if overt movement is consciously inhibited in a scanner 1116. | Execution of highly sequenced fine-motor commands; spatial-motor translation; somatosensory integration from instrument contact 1014. |
| Dorsolateral Prefrontal Cortex (DLPFC) | General attention maintenance to the auditory stream and transient working memory encoding 414. | Extreme cognitive load management: sight-reading, working memory maintenance of upcoming phrases, and executive motor planning 1415. |
| Anterior Cingulate Cortex (ACC) | Affective evaluation and salience detection of sudden harmonic transitions or dynamic shifts 12. | Sustained focused attention, performance monitoring, and real-time conflict resolution during complex bimanual motor execution 1214. |
| Default Mode Network (DMN) | High engagement probability in non-musicians, facilitating mind-wandering, episodic memory retrieval, and personal emotional association 1218. | Generally strongly suppressed in favor of the Executive Control Network to focus entirely on external task execution and strict temporal constraints 1219. |
| Cerebellum & Basal Ganglia | Rhythmic entrainment, internal beat perception, and continuous temporal expectation generation 710. | Deep motor sequence learning, micro-timing precision, forward-model error prediction, and fine bimanual coordination 1023. |
Structural Neuroplasticity: The Musician's Brain as an Experience-Dependent Model
The rigorous, daily demands of active musical performance serve as one of neuroscience's premier models for investigating experience-dependent structural neuroplasticity. Longitudinal and cross-sectional neuroimaging studies utilizing Voxel-Based Morphometry (VBM) and Diffusion-Weighted Imaging (DWI) consistently and robustly demonstrate that extensive musical training fundamentally alters both gray matter macrostructure and white matter microstructural integrity across the human brain 17202122. The brains of expert musicians are anatomically distinct from those of non-musicians, reflecting the extreme computational requirements of their profession.
White Matter Architecture and Interhemispheric Communication
Instrumental musicians, particularly those relying on complex, asynchronous bimanual coordination such as pianists and guitarists, exhibit profound structural adaptations in the corpus callosum (CC). The corpus callosum is the brain's largest interhemispheric white matter fiber bundle, absolutely critical for rapidly integrating sensory and motor information between the left and right cerebral hemispheres. High-resolution DWI and tractography reveal significantly greater fractional anisotropy (FA) - a highly sensitive metric indicating robust white matter tract integrity, fiber density, and heavy myelination - specifically in the anterior portion and the splenium of the CC in professional musicians compared to their non-musical peers 172123.
This microstructural enhancement directly facilitates decreased interhemispheric transfer times and significantly stronger interhemispheric inhibition (IHI). IHI is a neurophysiological mechanism that allows one hemisphere to momentarily suppress the motor cortex of the opposite hemisphere, a capability that is essential for maintaining independent, highly asymmetric control of the hands (e.g., a pianist playing a rapid staccato melody with the right hand while playing a slow, legato bassline with the left) 23. Crucially, these white matter adaptations are heavily age-dependent. The most dramatic callosal hyper-development is consistently observed in musicians who commenced formal, rigorous training before the age of seven. This finding underscores a critical developmental sensitive period for use-dependent white matter myelination 17212324. Furthermore, structural connectivity extends beyond the corpus callosum; both vocalists and instrumentalists show increased microstructural integrity in the arcuate fasciculus, the primary tract connecting temporal auditory regions with frontal motor regions, which facilitates the rapid auditory-motor integration essential for immediate pitch matching and vocal correction 1725.
Gray Matter Hypertrophy and Cortical Remapping
Beyond the white matter highways, VBM analyses reveal that musicians possess significantly higher gray matter volumes and increased cortical thickness in several discrete, specialized cortical and subcortical regions 202426.
Within the primary motor and somatosensory cortices, enlarged volumes are noted in both the precentral and postcentral gyri. Notably, the typical leftward asymmetry found in the motor cortex of right-handed individuals is significantly reduced in musicians, reflecting the heavy, symmetric bimanual demands of instrumental play 2426. In string players, a remarkable phenomenon of cortical remapping occurs, wherein the brain allocates a significantly greater cerebral representation to the specific digits most actively engaged - specifically the left-hand fingers used for rapidly pressing the strings - compared to non-musicians 24.
The cerebellum, which is absolutely essential for fine motor control, precise millisecond timing, and the continuous updating of motor sequence learning, displays significantly increased gray matter density in professional musicians 232427. In the auditory domain, Heschl's gyrus and the planum temporale show marked hypertrophy. Cortical thickness in these superior temporal regions can be up to 30% greater in highly trained musicians, a structural adaptation that directly correlates with augmented pitch resolution, complex timbre discrimination, and an expanded auditory working memory capacity 2426. Furthermore, structural asymmetry and increased overall volume are frequently noted in the hippocampus of musicians. This hippocampal enhancement is believed to facilitate the encoding, spatial mapping, and rapid retrieval of complex, multi-layered episodic memories associated with memorizing extensive musical scores and performance sequences 202426. Finally, practicing musicians exhibit greater gray matter volume in the left inferior frontal gyrus (including Broca's area) and appear neurologically less sensitive to typical age-related volumetric decline in this region, suggesting a neuroprotective effect of lifelong musical engagement 2426.
Dopaminergic Reward Pathways and Default Mode Network Modulation in Affective Disorders
To fully comprehend how music functions as an affective regulator and a potent clinical intervention for mental health, it is imperative to trace its mechanistic interaction with the brain's reward architecture and the Default Mode Network (DMN).
The Neurobiology of Musical Reward and Predictive Coding
Despite lacking any obvious biological necessity for human survival, music is an extraordinarily potent activator of the brain's dopaminergic reward system. The primary mechanism driving this reward response relies on predictive coding inherent in musical syntax. As a listener engages with a piece of music, their auditory cortex continuously generates subconscious predictions regarding the upcoming trajectory of harmony, rhythm, and melody. The subsequent fulfillment, delay, or deliberate violation of these expectations by the composer drives intense phasic dopamine release in the mesolimbic pathway 1628.
Functional imaging consistently demonstrates that the nucleus accumbens (NAcc), the ventral tegmental area (VTA), and the caudate nucleus exhibit heightened centrality and synchronized functional connectivity with the auditory cortices during the experience of pleasurable music listening 1624. In extensively trained musicians, the NAcc forms exceptionally strong intrinsic resting-state connections with ventromedial frontal areas and the temporal pole. This dense wiring allows for a much deeper, more complex integration of acoustic features with top-down emotional valuation and cognitive appraisal 1224.
Modulating the DMN in Psychopathology
The Default Mode Network - comprising core hubs such as the anterior medial frontal cortex, ventral medial prefrontal cortex, posterior cingulate cortex (PCC), and the precuneus - is a large-scale brain network that becomes highly active during states of wakeful rest, self-referential thought, and autobiographical memory recall 1819. However, in major depressive disorder (MDD) and various stress-related psychiatric pathologies, the DMN frequently becomes highly dysregulated.
Pathological hyperconnectivity within the DMN, and particularly between the DMN and limbic regions such as the amygdala and the subgenual prefrontal cortex, correlates heavily with clinical severity 1829. This hyperconnectivity manifests behaviorally as clinical rumination, severe anhedonia, and repetitive negative thought (RNT) 18293035. In these depressive states, the patient's brain is essentially trapped in an internally directed, highly insulated loop of negative self-reference 1835. Modern neuromodulation techniques, such as non-invasive transcranial focused ultrasound (tFUS) targeting the anterior medial prefrontal cortex, actively seek to disrupt this hyperconnectivity to alleviate depressive symptoms 30.
Music therapy operates on a remarkably similar functional principle, serving as a powerful, non-invasive exogenous modulator capable of disrupting this pathological hyperconnectivity. When subjects actively engage with music - particularly highly structured, predictable, or heavily rewarding music - the Salience Network (SN) and the Executive Control Network (ECN) must be recruited to process the complex external auditory stimuli. Due to the brain's intrinsic network dynamics, the activation of the ECN naturally downregulates and decouples the hyperactive DMN, pulling the individual out of the internal ruminative loop and forcing attention outward 1929. Conversely, in specific therapeutic contexts, listening to highly emotionally resonant or nostalgic music can selectively and safely engage specific hubs of the DMN. This targeted engagement facilitates adaptive emotion regulation, memory reconsolidation, and psychological catharsis, actively altering the baseline ratio of within-DMN connectivity to extra-DMN functional connectivity 121935. It is this precise, bidirectional capability to "tune" the functional connectivity between the SN, ECN, and DMN that positions music as a uniquely potent neuromodulatory intervention for affective disorders 71829.
Cross-Cultural Neuroscience: Universality vs. Cultural Mediation
A central, historically contentious debate within auditory cognitive neuroscience and ethnomusicology is the extent to which human brain responses to musical syntax, rhythmic structures, and harmony are biologically hardwired (universal) versus acquired via lifelong enculturation (mediated entirely by regional familiarity). Recent massive, global data sets utilizing advanced computational modeling have provided highly nuanced answers, rejecting extreme relativism while acknowledging the profound, localized impact of cultural mediation.
Form-Function Universality
A landmark 2018 ethnomusicological study (the Natural History of Song project) analyzed the psychological responses of thousands of naive listeners across 60 countries to brief, 14-second excerpts of vocal music drawn from 86 highly diverse, small-scale human societies (including isolated hunter-gatherers and pastoralists). The computational data unequivocally confirmed the existence of a universal biological link between acoustic form and social function.
Independent of any prior cultural exposure or linguistic comprehension, human listeners worldwide could accurately and reliably differentiate lullabies from dance songs, healing chants, and love songs 313233. The biological imperative drives this universality: lullabies universally feature slower tempi, descending pitch contours, and melodic simplicity to actively downregulate autonomic arousal in infants, whereas dance songs universally utilize sharp rhythmic punctuation and higher temporal density to drive motor entrainment and synchronous group arousal 313233.
Rhythmic Priors: A Universal Biological Bias with Cultural Dialects
In 2024, researchers from MIT and the Max Planck Institute conducted a massive cross-cultural rhythm perception study involving 39 distinct societal groups across 15 countries, actively moving away from traditional reliance on Western, Educated, Industrialized, Rich, and Democratic (WEIRD) undergraduate populations 3440. The researchers utilized an elegant iterative tapping task to uncover the internal mental representations - known as Bayesian "priors" - of rhythm.
The neurological mechanism revealed by the study highlighted a profound universal biological foundation: the human brain inherently exhibits a strict bias toward rhythmic structures composed of simple integer ratios (e.g., forming a 1:1:1 or 2:1 temporal relationship) 34. This bias acts as an evolutionary, preattentive error-correction mechanism. When humans hear imprecise timing, their brains naturally "snap" the incoming auditory signals into predictable, integer-based mathematical grids. This neuro-computational feature is essential for synchronized group movement, information transmission, and social cohesion 3435.
However, which specific integer ratios dominate a particular listener's mental grid is entirely and deeply determined by cultural exposure. While listeners from North America and Western Europe universally gravitated toward standard 4/4 ratios due to ubiquitous pop music exposure, traditional musicians in Turkey, Bulgaria, and Botswana defaulted to a complex 2:2:3 prior 3440. Even more distinctly, Malian drumming groups exhibited intense biases toward complex 7:2:3 subdivisions 3440. Thus, the neurobiological mechanism for rhythmic categorization (integer ratio bias) is universal, but the specific syntactic vocabulary utilized by the brain is culturally acquired 40.
Harmony and Affective Valuation
Neuroscientific investigations into the perception of acoustic consonance and dissonance have similarly refined Western-centric assumptions. Aversion to extreme acoustic dissonance - which is caused mechanically by sensory roughness and beating frequencies competing on the basilar membrane in the cochlea - appears to be a cross-cultural biological universal, as it triggers brainstem-level alarm responses 36.
However, the specific emotional valence attached to broader harmonic modes - most notably the fundamental Western paradigm that major chords map to "happy/positive" emotions while minor chords map to "sad/negative" emotions - is an entirely learned cultural construct 36. Cross-cultural data indicates that this strict affective distinction is not neurologically intrinsic to human auditory processing; in fact, the major triad only became conspicuously more prominent than the minor triad in Western polyphony during the 15th and 16th centuries 36. This finding highlights the brain's extraordinary plasticity in assigning complex emotional meaning to arbitrary acoustic phenomena based on repeated cultural exposure.
Clinical Interventions: Efficacy, Mechanisms, and Methodological Limitations
The direct translation of music neuroscience into clinical practice - specifically formalized as neurologic music therapy (NMT) - has gained substantial empirical traction. However, while the outcome data is compelling, differentiating the specific neurobiological mechanisms of the music itself from the general therapeutic benefits of social stimulation remains a profound methodological challenge in the field.
Depression and Affective Disorders in Dementia
Recent, highly rigorous network meta-analyses (2024-2026) evaluating music therapy for severe depression, particularly in elderly cohorts grappling with dementia and subjective cognitive decline, demonstrate robust clinical efficacy. A major network analysis incorporating 14 randomized controlled trials (RCTs) comprising 1,080 participants identified that active music therapy combined with singing (AMT + Sing) is by far the most effective intervention for ameliorating severe depressive symptoms.
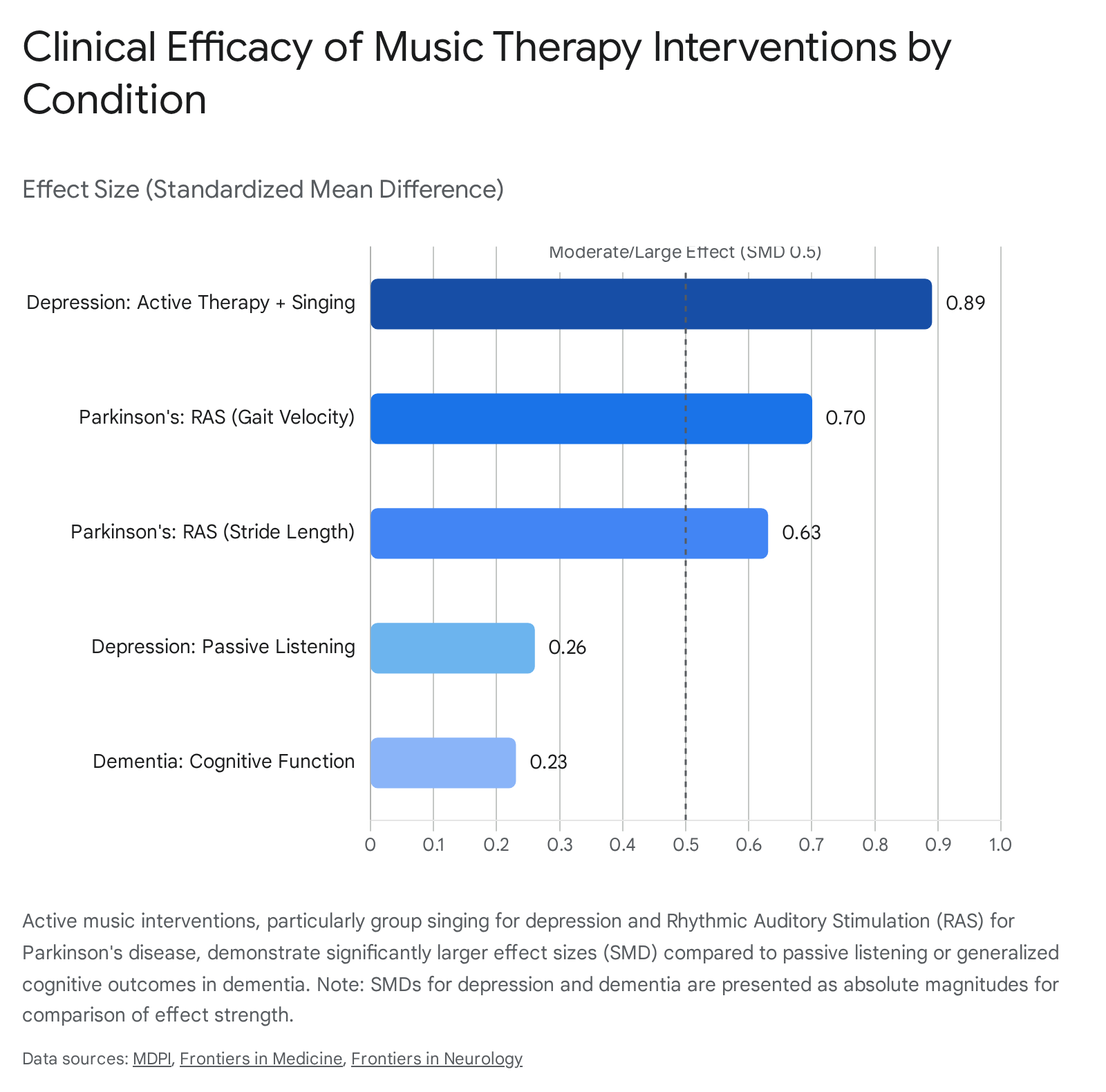
This approach yielded a highly significant, massive Standardized Mean Difference (SMD) of $-0.89$ 3738. In stark contrast, passive music listening alone produced a much smaller, albeit still positive, clinical effect (SMD = $-0.26$) 37.
The biological mechanism driving this profound efficacy in active therapy is multifaceted. It involves the direct dopaminergic regulation of the reward circuit to override clinical anhedonia, alongside the aforementioned decoupling of pathological DMN hyperconnectivity 1830. Furthermore, the physical act of active group singing induces the measurable release of oxytocin and endogenous opioids, directly mediating social bonding, reducing feelings of isolation, and effectively downregulating the hypothalamic-pituitary-adrenal (HPA) stress axis 353940.
Parkinson's Disease and Rhythmic Auditory Stimulation
Parkinson's disease (PD) results from the progressive degeneration and death of dopaminergic neurons in the substantia nigra pars compacta. This cellular loss severely impairs the basal ganglia's ability to internally sequence and initiate motor actions, leading to the hallmark symptoms of severe resting tremor, rigidity, bradykinesia, and debilitating gait disturbances 41.
Rhythmic Auditory Stimulation (RAS) - a highly specific, evidence-based NMT technique - leverages the brain's intact auditory-motor entrainment pathways to mechanically bypass the damaged basal ganglia circuitry. By utilizing an external, highly predictable auditory beat (such as a metronome or heavily rhythmic music), the auditory cortex projects timing signals directly to the supplementary motor area and premotor cortex via robust cerebellar pathways, effectively establishing an exogenous timing cue for continuous motor execution 104042. Meta-analyses of older adults with PD confirm that RAS training significantly and durably improves fundamental motor parameters, specifically gait velocity (SMD = $0.70$) and stride length (SMD = $0.63$) 41. However, the clinical data reveals highly mixed or non-significant effects on cadence, overall balance, and the complex phenomenon of freezing of gait. This indicates that while RAS excels at establishing continuous, linear temporal parameters for locomotion, it is markedly less effective for supporting the complex, multidirectional postural adjustments required for balance 41.
Dementia and Alzheimer's Disease Management
In the context of Alzheimer's disease and related dementias, music therapy is primarily deployed not as a cure, but as a critical tool to manage Behavioral and Psychological Symptoms of Dementia (BPSD) and to temporarily mitigate the outward signs of cognitive decline. The processing of highly familiar music relies heavily on the medial prefrontal cortex and the hippocampus 2040. Because musical memory is often encoded alongside intense emotional valence during youth, these specific neural networks exhibit remarkable resilience against early-stage neurodegenerative tau and amyloid pathology.
Clinical interventions demonstrate moderate but reliable efficacy in improving short-term global cognitive function (SMD = $-0.23$), increasing verbal fluency, and significantly reducing episodes of agitation and generalized anxiety in care home settings 43444546. A recent Cochrane review reinforces that music therapy offers tangible benefits beyond standard group activities by providing an engaging, non-pharmacological avenue to support mood, which is vital given the severe adverse effects often associated with antipsychotic medications in this population 45.
| Clinical Condition | Primary Intervention Type | Target Clinical Outcome | Effect Size (SMD) | Proposed Neurobiological Mechanism |
|---|---|---|---|---|
| Depression (Elderly/Dementia) | Active Music Therapy + Singing | Severe reduction of depressive symptoms and clinical anhedonia. | $-0.89$ (Large) 37 | Disruption of pathological DMN hyperconnectivity; direct dopaminergic reward activation; oxytocinergic mediation of social bonding 183040. |
| Depression (Elderly/Dementia) | Passive Music Listening | General mood stabilization. | $-0.26$ (Small) 37 | Autonomic arousal regulation; parasympathetic activation; limbic system soothing 37. |
| Parkinson's Disease | Rhythmic Auditory Stimulation (RAS) | Improvement in Gait Velocity. | $0.70$ (Moderate) 41 | Auditory-motor entrainment mechanically bypassing the damaged basal ganglia; direct cerebellar-SMA temporal cueing 4042. |
| Parkinson's Disease | Rhythmic Auditory Stimulation (RAS) | Improvement in Stride Length. | $0.63$ (Moderate) 41 | Continuous sensorimotor timing phase-locking 4041. |
| Dementia | General Music Therapy | Global Cognitive Function enhancement. | $-0.23$ (Small) 43 | Engagement of preserved, emotionally valenced autobiographical memory networks (hippocampal-mPFC axes) 4044. |
| Dementia | Group Receptive Therapy | Reduction in Caregiver Burden. | $0.82$ (Large) 53 | Shared emotional resonance and mutual reduction in sympathetic nervous system arousal between patient and caregiver 53. |
Methodological Limitations and Competing Views in Music Therapy Research
While the clinical data supporting NMT is highly promising, maintaining rigorous neuroscientific integrity requires a frank acknowledgment of the significant limitations and competing views within current literature. First and foremost, there is high heterogeneity in study design across the literature. Varying exposure times (from single 30-minute sessions to 24-week longitudinal programs), inconsistent frequency of therapy, and a widespread lack of standardized, manualized protocols frequently dilute statistical power and complicate cross-study comparisons 414748.
A critical, almost intractable challenge in assessing active music therapy (such as group drumming or choir singing) is the immense difficulty of isolating the specific acoustic variable of music from the powerful confounding variables of intensive social engagement, structured routine, sustained attention, and the high empathy of the clinical therapist 4648. Critics rightly argue that some of the massive effect sizes seen in active therapies may be attributed more to the alleviation of severe social isolation than to the neurobiology of auditory processing itself.
Furthermore, true double-blinding in these RCTs is inherently problematic, if not impossible. Patients, caregivers, and assessors are usually acutely aware of the intervention assignment, introducing significant expectation bias and the placebo effect into self-reported or observer-rated scales 41. Finally, while transient functional connectivity changes, behavioral improvements, and mood stabilization are exceptionally well-documented post-intervention, there is currently a stark lack of robust longitudinal MRI evidence proving that late-life music interventions induce the kind of disease-modifying structural neuroplasticity capable of genuinely halting or reversing neurodegeneration 40.
Conclusion
The contemporary neuroscience of music demonstrates unequivocally that engaging with organized, rhythmic sound is one of the most computationally demanding, multimodal tasks the human brain can perform. The data thoroughly debunks archaic left/right hemispheric dichotomies, revealing instead a highly dynamic, globally distributed network dependency that shifts and refines itself through years of expert practice. The act of passive listening requires intricate predictive coding, rapid syntactic analysis, and limbic valuation, whereas active musical training physically sculpts the brain's macro- and micro-architecture, profoundly augmenting white matter interhemispheric communication and gray matter sensorimotor networks.
Crucially, the brain's response to music is neither entirely innate nor entirely a product of cultural blank-slate learning. Rather, evolutionary biology provides a universal computational scaffolding - evidenced by integer-ratio rhythmic priors and brainstem-level dissonance aversion - upon which local culture maps highly specific syntactic rules and emotional vocabularies. As clinical neurology continues to integrate these neurobiological findings, targeted therapies like Rhythmic Auditory Stimulation and DMN modulation via active singing are successfully transitioning from adjunctive, alternative treatments to empirically validated, mechanistic interventions. To fully realize this profound therapeutic potential, future psychiatric and neurological research must prioritize large-scale, rigorously controlled, and structurally imaged longitudinal studies capable of isolating acoustic mechanisms from psychosocial variables, ultimately charting the precise therapeutic thresholds of human musicality.