Neurobiology and function of forgetting
For decades, the neurobiological study of memory was heavily biased toward understanding persistence - the cellular, systemic, and psychological mechanisms by which the central nervous system acquires, consolidates, and retrieves information. Within this classical paradigm, forgetting was primarily viewed as a passive deterioration, an inevitable failure of the memory trace, or an unavoidable consequence of biological aging and neurological disease. However, contemporary research across neuroscience, cognitive psychology, and computational modeling has catalyzed a significant paradigm shift. Evidence now demonstrates that forgetting is not a passive decay but an active, heavily regulated, and energy-consuming biological process.
The brain engages in a constant physiological competition between memory formation and active erasure. This continuous pruning of information is critical for optimizing decision-making, ensuring cognitive flexibility, and preventing the neurological equivalent of overfitting to environmental stimuli. From the molecular signaling cascades within individual dendritic spines to the psychological frameworks governing learning and recall, the capacity to forget is as essential to human intelligence as the capacity to remember.
Evolutionary and Computational Foundations
The assumption that an optimal memory system would permanently retain all incoming sensory data is challenged by both evolutionary biology and computational theory. If the central nervous system recorded and seamlessly recalled every trivial detail of daily existence, the resulting cognitive load would paralyze decision-making and environmental adaptation. William James observed that selection is the keel on which the mental ship is built, a concept illustrated by Jorge Luis Borges' fictional character Funes the Memorious, whose inability to forget rendered him incapable of generalizing or prioritizing information 1.
Memory Optimization and Decision Making
The primary evolutionary function of memory is not to serve as an infallible archival record of the past, but to act as a predictive engine that guides intelligent decision-making in dynamic environments. Neurobiological frameworks propose that an interaction between persistence (remembering) and transience (forgetting) is required to navigate a noisy world effectively 23.
Transience enhances behavioral flexibility by systematically reducing the influence of outdated information. In environments that frequently change, historical data can become a liability. By actively erasing obsolete memories, the brain prevents conflicting memory traces from competing during retrieval, thereby reducing the cognitive resources required to make an informed choice 24. Furthermore, active forgetting allows the brain to generalize past events to novel situations. By stripping away hyper-specific, irrelevant details of an episodic memory, the neural network distills the underlying schema of an experience, which is vastly more applicable to future, similar encounters 35.
Biological Parallels in Artificial Intelligence
The necessity of forgetting is tightly mirrored in the development of artificial neural networks (ANNs) and large language models (LLMs). In machine learning, an algorithm that memorizes its training data perfectly will fail to perform well on new, unseen data - a phenomenon known as overfitting 25. To counteract this, artificial intelligence models employ regularization techniques that force the network to ignore specific details and prioritize core patterns, enabling wider application 34.
Standard dropout techniques in ANNs, which randomly deactivate neurons to force the network to find redundant pathways, serve as a rudimentary form of artificial forgetting. More recent advances utilize magnitude-based synaptic pruning that closely emulates biological processes. By gradually removing weak connections based on their absolute magnitudes across layers, these models achieve higher generalization and avoid catastrophic interference - a state where learning new information completely overwrites previously learned tasks 67.
Furthermore, researchers are increasingly implementing "refresh learning" and "machine unlearning" mechanisms in LLMs. These systems selectively erase outdated knowledge while utilizing Fisher information matrices to preserve memory stability across different domains, mimicking the hippocampal trisynaptic circuit's ability to balance memory retention and active forgetting 889. These computational models demonstrate that selective forgetting separates noise from signal, fundamentally enhancing a system's adaptability to continuous data streams 57.
Cellular and Molecular Mechanisms
The physiological erasure of memory is driven by highly specific molecular, cellular, and network-level mechanisms. The central nervous system utilizes multiple, layered pathways to dissolve memory traces and modulate synaptic strength over time.
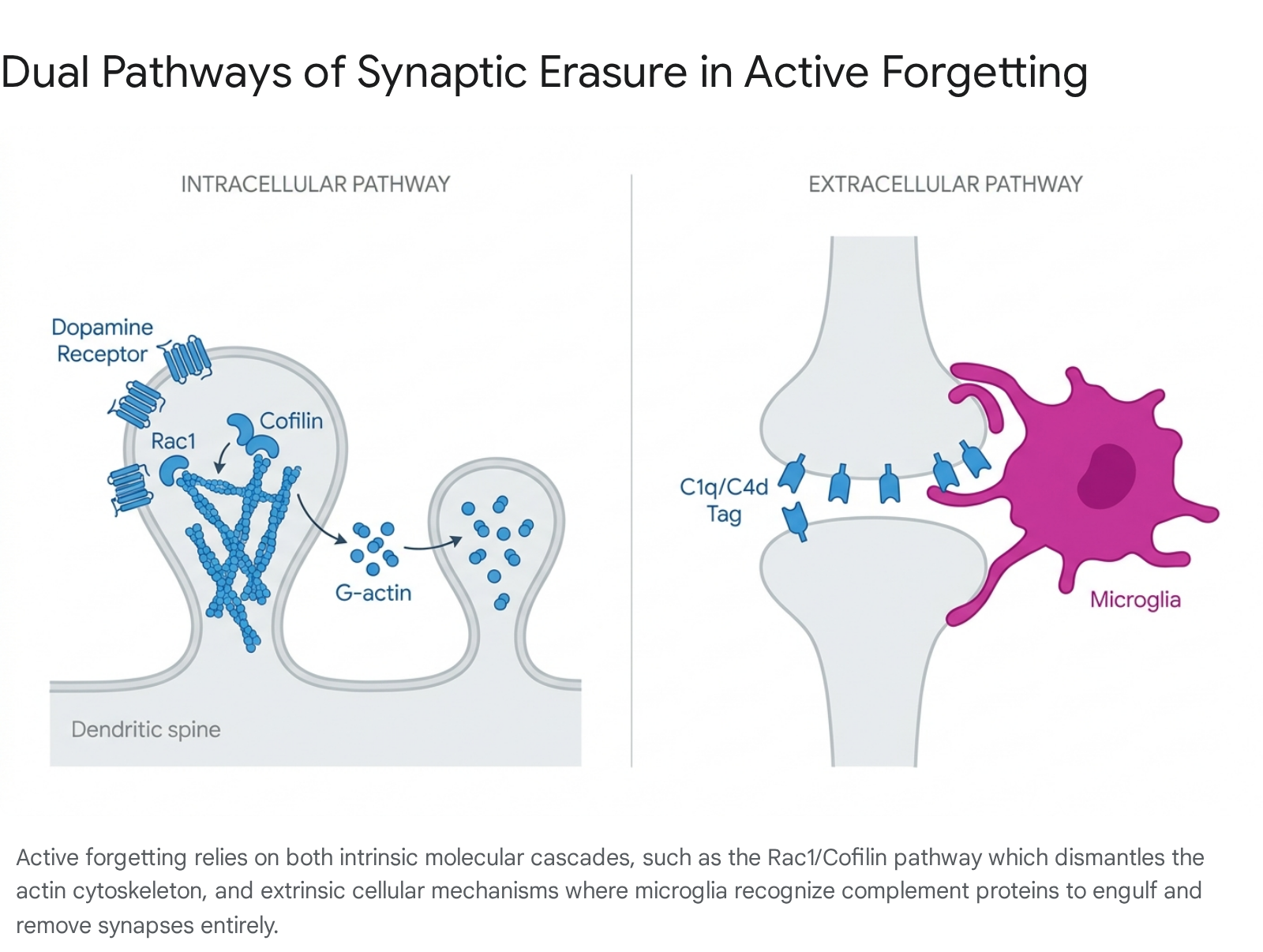
Intracellular Signaling and Actin Remodeling
At the molecular level, memory traces are physically encoded by structural changes in the actin cytoskeleton within dendritic spines. Active forgetting degrades these structural changes through intrinsic biochemical pathways. Combined behavioral and genetic studies in Drosophila and mammalian models have identified a complex system of cellular active-forgetting pathways heavily regulated by dopamine neurons 1011.
Ongoing activity in specific subsets of dopaminergic neurons works to erode newly formed memories immediately after acquisition. This signal is received by postsynaptic dopamine receptor subtypes - such as DAMB in insects or D1 homologs in mammals - mobilizing an intracellular signaling cascade 101112. The signal is communicated through a protein complex scaffolded by Scribble, which terminates in the activation of small GTPases, particularly Rac1 1112. Rac1 activation triggers cofilin, an actin-severing protein. Cofilin destabilizes filamentous actin (F-actin) and reverts it to globular actin (G-actin), thereby dismantling the structural scaffolding of the dendritic spine that houses the memory trace 131415. Conversely, LIMK-dependent inactivation of cofilin stabilizes F-actin, promoting memory consolidation 1314.
Experimental manipulation underscores the precise control this pathway exerts: expressing constitutively active mutant Rac1 in the mouse hippocampus accelerates the decay of object-recognition memory threefold. In contrast, inhibiting Rac1 activity - or inactivating cofilin - effectively prevents natural memory decay and blocks interference-induced forgetting 121314. Thus, intrinsic active forgetting serves as the default, energy-consuming state of the brain, constantly promoting memory erasure to balance processes like consolidation 1215.
Extracellular Synaptic Pruning
Beyond intracellular actin remodeling, the brain utilizes immune-mediated cellular mechanisms to eliminate entire synapses. Synaptic pruning is an essential neurodevelopmental and maintenance process that removes weak or redundant connections to increase the efficiency of neural circuits. This process follows a use-dependent rule; heavily utilized pathways are strengthened, while inactive pathways are chemically flagged for removal 61617.
Microglia, the resident immune cells of the brain, play a central role in this process by engulfing and degrading synaptic material. Recent neurobiological debate focuses on the precise conceptual framework of this interaction, resulting in two primary models of microglial pruning.
| Pruning Framework | Biological Mechanism | Microglial Role | Therapeutic Implication |
|---|---|---|---|
| Synapse Culling | Microglia act as active hunters that autonomously select, cleave, and degrade intact but unwanted synapses based on external chemical tags. | Active initiator and executioner of synapse destruction. | Blocking microglial activity directly halts the destruction of live synapses. |
| Synapse Scavenging | Neurons internally determine which synapses to remove and physically shed the material. Microglia subsequently arrive to clean up the debris. | Passive waste-management and phagocytosis of pre-shed material. | Blocking microglial activity causes a buildup of necrotic synaptic debris, but does not save the functional synapse. |
The mechanisms driving both potential pruning models rely heavily on the complement cascade - an innate immune surveillance system. Signal proteins such as C1q and C4d act as tags on weak synapses, while CD47 acts as a protective signal preventing engulfment 171819. Dysregulation of these signals has profound developmental and pathological consequences. Under-pruning is implicated in delayed development and autism spectrum disorders, while excessive pruning is linked to schizophrenia and difficulty processing information 1617.
Pathological states can hijack these normal forgetting mechanisms. In Alzheimer's disease models, the buildup of amyloid beta and chronic brain inflammation converge on a specific receptor, LilrB2. This interaction erroneously triggers neurons to upregulate complement cascade proteins like C4d, flipping a molecular switch that causes neurons to actively invite the destruction of their own connections, leading to severe memory loss 18. Similarly, neurotropic viral infections can disrupt synaptic pruning by overactivating the complement system and inflammatory cytokines, driving aberrant synaptic elimination 20.
Neural Activity Decay and Human Development
The macro-level effects of synaptic pruning on cognitive efficiency have been successfully mapped in human development. Utilizing electroencephalography (EEG), researchers investigated the developmental timelines of synaptic pruning and cognitive control during adolescence. A recent study recorded brain activity from 179 adolescents performing an adaptive response inhibition task (the Eriksen flanker) at ages 12, 15, and 18, generating 331 recordings across 104 electrodes 21.
The findings indicate that neural activity decay - the speed at which dynamic neural activity returns to a baseline unperturbed state - is significantly slower in early adolescence compared to late adolescence. As the overall number of synapses is reduced through natural pruning between ages 12 and 18, the removal of recurrent, inefficient processes enhances the speed of information transfer. This structural refinement correlates directly with increased behavioral cognitive control, validating theoretical models that posit forgetting and pruning as fundamental drivers of adolescent cognitive maturation 21.
Engram Dynamics and Network Topology
Memories are physically encoded in "engrams" - distributed, sparsely populated networks of neurons that undergo long-lasting modifications in gene expression and synaptic connectivity during learning, and are subsequently reactivated during memory retrieval 11222324. The dynamic interaction between memory consolidation and forgetting continuously alters the topology and accessibility of these engram networks.
Systemic Reorganization and Neurogenesis
At extended timescales, forgetting is heavily modulated by systemic neural reorganization, notably through adult neurogenesis in the dentate gyrus of the hippocampus. As new neurons are continuously generated and integrated into existing hippocampal circuits, they form new connections that compete with older synapses. This constant physical integration rewires existing neural networks, which enhances the capacity to encode novel information but simultaneously degrades the structural integrity and accessibility of pre-existing memory traces 10122526.
Neurogenesis-based forgetting provides a leading biological explanation for infantile amnesia - the inability of human adults to recall episodic events from before the age of three or four. During infancy and early childhood, the rate of hippocampal neurogenesis is at its absolute lifetime peak, resulting in rapid circuitry rewiring that overwrites or functionally isolates the engrams holding early autobiographical memories 425. In adulthood, as neurogenesis slows, memories become more stable, though the hippocampus continues to undergo "representational drift," where the neuronal representation of task variables gradually changes over days to weeks due to stochastic synaptic fluctuations and continuous learning 2427.
Engram Core Contamination and Retroactive Interference
Engrams are not static biological records; they are highly dynamic entities vulnerable to structural reorganization. A major driver of active forgetting is retroactive interference (RI), a phenomenon where the acquisition of new information disrupts recently consolidated memories.
Recent network-level analyses utilizing mouse models of object location memory reveal that retroactive interference induces forgetting by physically infiltrating the "engram core" during a discrete biological consolidation window. Memory consolidation requires de novo protein synthesis. If a novel experience occurs within a few hours of the initial learning event, the neurons activated by the new experience intermingle with the original engram, destabilizing the structured coactivity network 1528.
This "engram core contamination" reduces the probability of reactivating the original cells and degrades network topology by increasing synaptic edge turnover. Over time, however, if the engram is allowed to consolidate without interference, it undergoes progressive maturation. It acquires greater network density, increased similarity, and k-core robustness. Once this maturation occurs, subsequent interference is confined to the network periphery, rendering the memory resistant to catastrophic forgetting 28.
Storage Failure Versus Retrieval Failure
A central controversy in the cognitive neuroscience of forgetting revolves around whether a forgotten memory is physically eradicated (storage failure) or simply rendered temporarily inaccessible (retrieval failure). Traditionally, forgetting was viewed as an irreversible storage degradation caused by synaptic decay or the death of engram cells. However, advanced neuro-technologies, including optogenetics, chemogenetics, and in vivo calcium imaging, have provided compelling evidence that natural forgetting is predominantly a retrieval failure 293031.
Researchers have repeatedly demonstrated that in cases of natural forgetting, retroactive interference, and even in transgenic mouse models of early-stage Alzheimer's disease, the original engram cells persist in a latent state 232831. While a subject may appear entirely amnesic behaviorally, artificially stimulating the originally tagged engram cells with light (optogenetics) or designer drugs (DREADDs) is sufficient to induce full memory retrieval 23242632.
These findings indicate that active forgetting mechanisms degrade the accessibility of the trace - perhaps by weakening specific synaptic retrieval pathways, altering AMPA receptor trafficking, or masking the engram with inhibitory GABAergic inputs - without completely erasing the core biological encoding 11233134.
| Paradigm | Storage Failure (Passive Erasure) | Retrieval Failure (Active Forgetting) |
|---|---|---|
| Primary Mechanism | Physical degradation of synapses; loss of engram cells over time. | Active modulation of synaptic strength; inhibitory masking; cue mismatch. |
| Trace Status | The memory trace is permanently lost, overwritten, or structurally corrupted. | The trace exists in an intact, latent, but inaccessible state. |
| Reversibility | Irreversible. Requires relearning the information from environmental input. | Reversible. Can be recovered via optimal contextual cues or artificial optogenetic stimulation. |
| Clinical Presentation | Late-stage neurodegeneration (e.g., severe Alzheimer's volumetric loss), traumatic brain injury. | Early-stage amnesia, retroactive interference, state-dependent forgetting. |
Cognitive Psychology and Learning Frameworks
While neurobiologists observe forgetting at the synaptic and network levels, cognitive psychologists study its functional utility in learning and behavioral adaptation. The most prominent pedagogical framework bridging forgetting and learning is the "New Theory of Disuse," pioneered by cognitive psychologists Robert and Elizabeth Bjork.
Storage Strength and Retrieval Strength
The Bjork framework postulates that human memory does not function as a simple recording device where repeated exposure guarantees retention. Instead, every memory possesses two theoretically independent dimensions: Storage Strength and Retrieval Strength 333435.
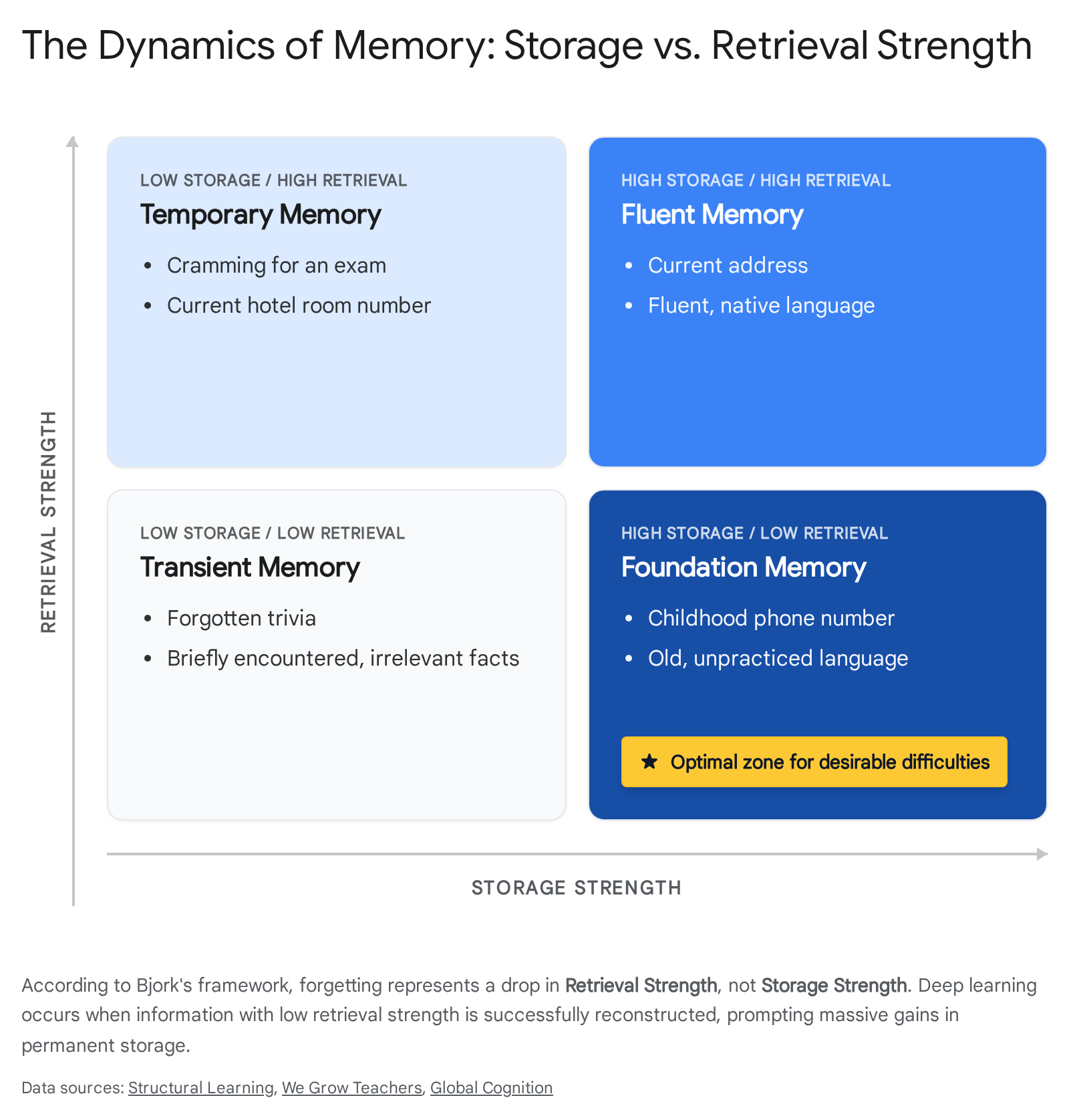
Storage Strength measures how deeply a piece of information is encoded in long-term memory. It accumulates gradually over time and is highly durable. The framework posits that once storage strength reaches a high threshold, it is permanent and never decreases - meaning well-encoded information is never truly "lost" 3334. In contrast, Retrieval Strength measures the immediate accessibility of a memory at a specific moment in time. Retrieval strength is highly volatile, heavily influenced by recency, and deeply dependent on environmental cues 3435.
The critical insight of this framework is the inverse relationship between the two metrics during the learning process. If retrieval strength is artificially high - such as immediately after reading a text, or when practicing the exact same mathematical formula repeatedly - re-studying the material produces negligible gains in storage strength because no cognitive reconstruction is required 343536. Conversely, when retrieval strength naturally decays - when a student has partially forgotten the material - the substantial cognitive effort required to successfully reconstruct the memory triggers a massive gain in permanent storage capacity 3437.
In this model, forgetting is fundamentally necessary for deep learning. The functional suppression of unused information decreases retrieval strength, which serves as the required biological catalyst for maximizing subsequent storage gains 3435.
Desirable Difficulties in Practice
To leverage the neurocognitive reality of storage and retrieval dynamics, educators and instructional designers employ "desirable difficulties" - strategies that intentionally inject friction into the learning process to stimulate active memory reconstruction. While these tasks initially slow down progress, decrease confidence, and increase error rates during practice, they profoundly enhance long-term retention and the transferability of knowledge 34363839.
The application of desirable difficulties manifests through several highly empirically supported techniques. Distributed practice, or the spacing effect, involves allowing sufficient time to pass between study sessions to induce natural forgetting. The subsequent struggle to recall the information exercises the brain's retrieval pathways, forming multiple cognitive routes to the data and reducing contextual dependency 343640. Interleaving instruction forces the brain into discrimination learning by mixing different topics or problem types within a single session. Instead of blindly applying a blocked strategy, the learner must constantly recall the appropriate rule from memory, effectively building out-of-context retrieval strength 343641.
Furthermore, the testing effect (retrieval practice) demonstrates that the act of generating an answer from memory modifies the underlying memory trace, rendering it more robust than passive re-reading. Re-reading provides a false illusion of fluency because the text is processed in short-term working memory; forced retrieval, however, requires the reconstruction of the trace from long-term storage, leveraging the "generation effect" 33363742. Crucially, the desirable difficulties framework notes a caveat: difficulties are only desirable if they are achievable. If retrieval strength falls to zero and the student lacks the foundational background knowledge to reconstruct the memory, the generation fails entirely, leading to frustration and cognitive overload rather than durable learning 33363941.
Extremes of Memory Retention and Loss
Examining the pathological extremes of memory - individuals who cannot forget, alongside aging populations experiencing accelerated forgetting - provides profound insights into the baseline necessity of memory erasure for normal psychological and neurological function.
Highly Superior Autobiographical Memory
Highly Superior Autobiographical Memory (HSAM), clinically termed hyperthymesia, is an extraordinarily rare condition in which individuals can recall an abnormally large number of personal life experiences in vivid, exhaustive detail. First documented in 2006 with the case of Jill Price (Patient AJ), fewer than 100 individuals have been identified globally with this condition 434445.
While often perceived culturally as a "superpower," individuals with HSAM frequently describe the condition as a severe psychological burden. Because they lack the active forgetting mechanisms that normal brains use to filter out irrelevant or painful information, HSAM individuals are often trapped in a perpetual, unbidden relationship with their past 454649. The inability to fade the emotional sting of negative memories leaves them highly prone to anxiety, depression, and obsessive-compulsive tendencies. Clinical assessments demonstrate that HSAM subjects score significantly higher on the Leyton Obsessional Inventory (a standard measure of OCD traits) than the general population, suggesting a deep phenomenological link between an inability to let go of thoughts and the inability to lose memory traces 45465047.
Neuroanatomical imaging reveals that HSAM individuals possess structural differences in the brain, including increased coherence of white matter tracts like the uncinate fasciculus (connecting the anterior temporal cortex with the orbital frontal cortex) and forceps major, alongside an enlarged parahippocampal gyrus 14849. Data from longitudinal studies of older HSAM individuals (such as 75-year-old patient GC) indicate that these enhanced memory capabilities manifest without apparent decline in physiological aging, utilizing wide networks of prefrontal and temporo-parietal regions during retrieval 50.
Interestingly, while HSAM individuals remember their own lives flawlessly, they do not possess superior memory for arbitrary laboratory data, distinguishing them from trained mnemonists. Furthermore, despite their vast recall, HSAM individuals are equally susceptible to the "misinformation effect" and false memories as neurotypical individuals. In laboratory settings using the Deese-Roediger and McDermott (DRM) paradigm and the nonexistent news-footage paradigm, HSAM subjects falsely recalled seeing footage of a plane crash at the same 20% to 29% rate as controls. This critical finding proves that even in brains with extraordinary retention, memory retrieval remains an inherently reconstructive process utilizing fallible mechanisms, rather than a perfect reproductive video recording 454951.
| Clinical Feature | Highly Superior Autobiographical Memory (HSAM) | Trained Mnemonists (e.g., Luria's "S", Patient TE) |
|---|---|---|
| Domain of Expertise | Personal life events, specific dates, and related public events. | Arbitrary data, long lists of numbers, words, or complex images. |
| Mechanism of Recall | Involuntary, automatic, effortless, and highly emotional. | Deliberate, effortful, utilizing highly practiced mnemonic techniques. |
| Standardized Memory Tests | Average or below-average performance on general memory tasks. | Exceptionally high performance on targeted memorization tasks. |
| Use of Synesthesia or Eidetic Imagery | Rare; relies on vivid episodic re-experiencing. | Common (e.g., time-space synesthesia, "Memory Palaces"). |
| Psychological Profile | High obsessive-compulsive traits; prone to absorption and fantasy; high burden of negative recall. | Varies widely, but often requires intense deliberate focus; memory does not intrude unbidden to the same degree. |
Accelerated Forgetting and Environmental Factors
At the opposite extreme, excessive or accelerated forgetting highlights vulnerabilities in the aging brain, providing data on how external variables influence intrinsic memory decay. Recent large-scale international neuroimaging mega-analyses reveal that age-related memory decline is not triggered by the failure of a single brain region or a single gene (like APOE ε4), but is instead the result of widespread structural changes and tissue shrinkage distributed across both cortical and subcortical regions 52.
Geographical, environmental, and socioeconomic diversity play a massive role in the rate of this cognitive decline. Analyses of global populations across 5,306 participants demonstrate that individuals in Latin American and Caribbean (LAC) countries exhibit significantly "older" brain ages compared to matched individuals in non-LAC countries (such as the US, UK, Japan, and China). This accelerated brain aging, primarily localized to frontal-posterior networks critical for memory and goal-driven tasks, is driven by environmental inequalities, including high pollution levels, disparate disease burdens, gender bias, and severe socio-economic stress 535854.
Furthermore, while high levels of education are traditionally thought to build a protective "cognitive reserve," recent analyses of over 15,157 MRI scans challenge this assumption. Education is strongly associated with a larger maximum intracranial volume developed early in life, slightly larger volumes of memory-sensitive regions, and better baseline episodic memory. However, education does not slow the actual rate of age-related memory decline or confer biological resilience against structural brain atrophy later in life. It merely provides a higher cognitive starting point from which the inevitable decline begins, emphasizing that the mechanisms of forgetting and neural decay operate independently of intellectual accumulation 5455.
Conclusion
The science of forgetting reveals a deeply elegant biological paradox: to construct a durable, adaptive, and intelligent mind, the brain must be uniquely equipped to dismantle its own architecture. Forgetting is not an error code within the human operating system; it is a meticulously engineered, energy-consuming feature that prioritizes generalization and decision-making over exhaustive archiving.
At the cellular level, intrinsic signaling cascades utilizing dopamine, Rac1, and cofilin work alongside the extrinsic immune activities of microglia to constantly sculpt the synaptic landscape, ensuring that only the most relevant neural connections survive 11131456. At the network level, engrams undergo discrete periods of vulnerability where retroactive interference rewires the topology of a memory trace, actively suppressing outdated information to prevent cognitive catastrophic interference 71528. Psychologically, the volatile decay of retrieval strength is the exact mechanism required to force the brain into deep, reconstructive processing, embedding critical information permanently into the architecture of thought through desirable difficulties 34353642.
By analyzing the pathological extremes - the paralyzing psychological burden of individuals with HSAM who cannot shed the weight of their past, and the devastating cognitive loss of those whose neurobiological pruning mechanisms run unchecked due to disease or environmental stress - it becomes evident that human intelligence exists in the delicate equilibrium between persistence and transience. As computational neuroscience advances, leveraging biological principles for machine unlearning and targeted memory modulation, the fundamental truth remains unchanged: optimal cognition requires the ability to let go.