Neurobiology of curiosity and learning mechanisms
Curiosity is a foundational cognitive mechanism that drives information-seeking behavior across the lifespan. Far from being a purely abstract psychological phenomenon, the drive to acquire knowledge is deeply rooted in the brain's metabolic architecture, neuromodulatory systems, and evolutionary imperative to resolve environmental uncertainty. The biological systems that reward learning rely on complex dopaminergic circuitry, hippocampal neuroplasticity, and precise metabolic homeostasis. However, as the brain ages - or as it encounters restrictive learning environments - the capacity and motivation for curiosity undergo significant physiological and structural transformations. This analysis examines the theoretical frameworks governing curiosity, the neural and metabolic circuitry that sustains it, the cognitive shifts that attenuate it during aging, and the implications for educational systems.
Theoretical Models of Information Seeking
Understanding why the brain rewards learning requires examining the theoretical models that define how knowledge gaps are perceived, processed, and resolved. Three primary frameworks dominate the current discourse: the Information Gap Theory, Active Inference, and the Goldilocks Effect.
The Information Gap and Novelty Theories
The Information Gap Theory, synthesized by George Loewenstein in 1994, posits that curiosity functions similarly to a physiological drive state, akin to hunger or thirst. According to this model, curiosity is triggered when an individual becomes consciously aware of a discrepancy between their current knowledge and a desired state of understanding 1234. This gap induces a feeling of cognitive deprivation, a mildly aversive state that the brain is highly motivated to resolve.
Empirical validation of this theory has demonstrated an inverted U-shaped relationship between an individual's confidence in their prior knowledge and their level of curiosity. In paradigm-defining studies, researchers presented subjects with trivia questions and recorded both their confidence in guessing the answer and their subjective curiosity 5. When individuals possessed zero knowledge (low confidence), curiosity was minimal because no cognitive baseline existed to form a gap. Conversely, when individuals were highly confident in their knowledge, curiosity was equally low. Curiosity peaked at intermediate levels of confidence, where the individual possessed enough foundational knowledge to recognize exactly what they did not know 5.
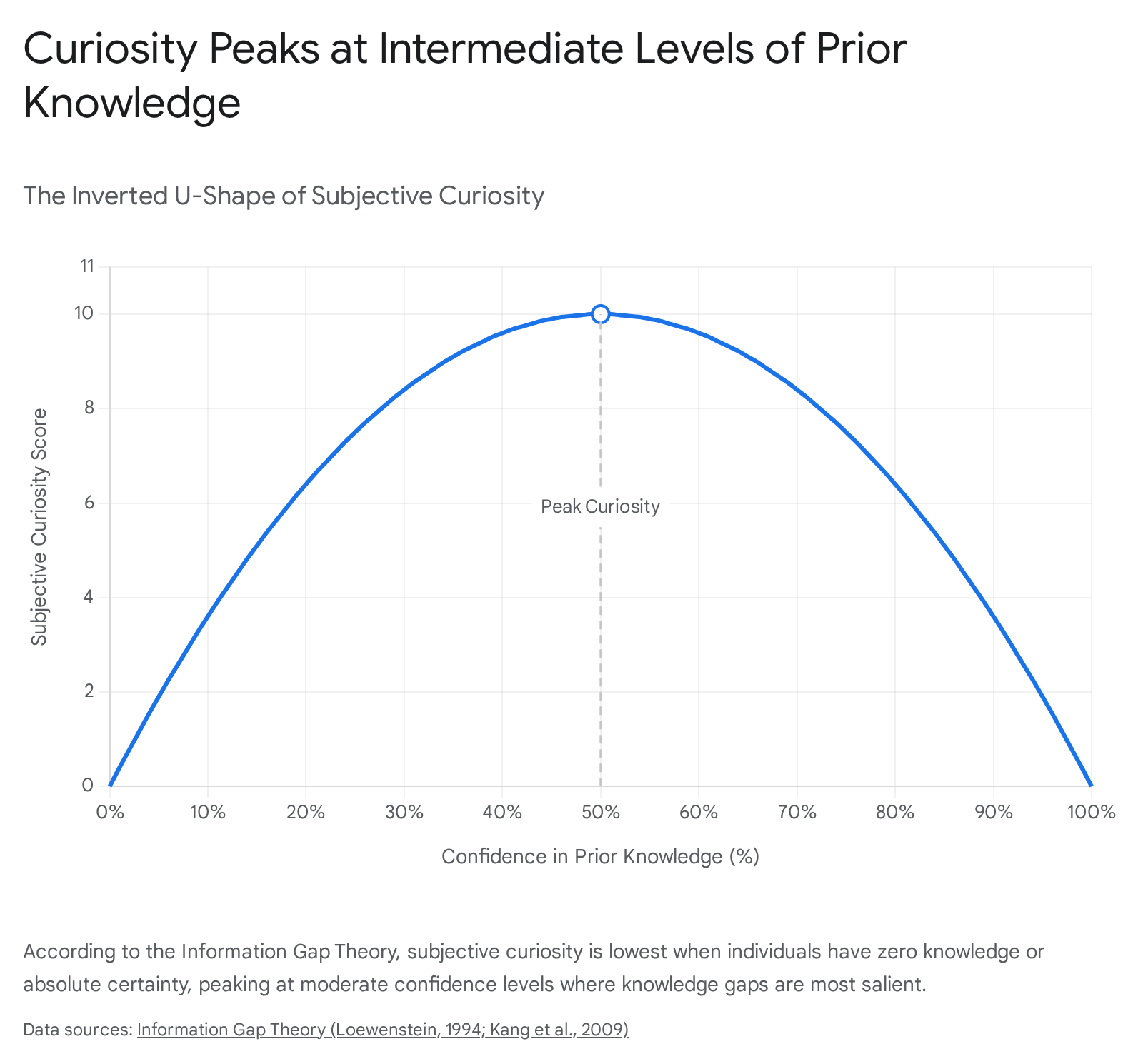
The Information Gap Theory has been further refined by the Interest/Deprivation (I/D) model, which distinguishes two qualitatively different experiences of curiosity. I-type (interest) curiosity is driven by a positive feeling of anticipating something enriching, while D-type (deprivation) curiosity is driven by an uncomfortable awareness of missing information 36. Recent psychological evidence indicates that these states yield different epistemic behaviors. Individuals high in discovery-based I-type curiosity show greater discernment and tolerance for ambiguity. Conversely, those driven by D-type curiosity are often so eager to resolve cognitive tension that they become less discriminating, accepting implausible claims or overclaiming knowledge of nonexistent concepts simply to close the gap 6.
While highly influential, the Information Gap Theory faces modern critiques. It struggles to fully account for diversive or novelty-seeking curiosity, where organisms exhibit strong exploratory drives toward completely novel stimuli for which they possess no prior knowledge to form a gap 5. Researchers have proposed rational models combining both paradigms, suggesting that the level of curiosity is determined by the expected value of knowing the information - a calculation that weighs both the novelty of the stimulus and the probability that the information will be useful in future environments 5.
Active Inference and the Free Energy Principle
An alternative, mathematically grounded paradigm is the Free Energy Principle, pioneered by Karl Friston. This model views the brain as a Bayesian inference engine perpetually attempting to minimize "surprise" or uncertainty about the sensory inputs it receives across its Markov blanket 789. Under this framework, behavior is driven by "active inference," a process in which natural agents act to fulfill prior beliefs and minimize the difference between their internal generative model of the world and their actual sensory perceptions 810.
Within the active inference framework, curiosity is not merely a reaction to a conscious knowledge gap, but an inherent, continuous epistemic foraging 96. Agents actively sample their environment to test plausible hypotheses about causal structures. When an agent successfully tests a hypothesis and reduces the expected variational free energy of its internal model, the system experiences a Bayesian model reduction. This sudden, efficient restructuring of internal knowledge networks correlates with the neurobiological mechanisms of "aha" moments or insight 79. Unlike the Information Gap Theory, active inference suggests that intelligence inherently entails information-seeking behavior to optimize adaptive fitness and thermodynamic efficiency 67. Explainability and learning are assured because the generative model continuously updates prior beliefs (priors) based on new evidence, refining them into more accurate models (posteriors) 10.
The Goldilocks Effect in Early Development
The theoretical intersection of environmental complexity and information-seeking is empirically observable in early human development through the "Goldilocks Effect." Infants, who face strict capacity limits on computational resources, must be highly selective in sampling information to learn efficiently 8910.
In foundational behavioral studies measuring visual and auditory attention, infants actively avoid allocating cognitive resources to sequences that are overly simple (highly predictable) or excessively complex (highly surprising or unknowable) 81011. By employing ideal learner computational models, researchers quantified the complexity of stimuli using negative log probability. The data revealed a distinct U-shaped curve in the probability of infants looking away from a stimulus: attention termination was highest at the extremes of complexity and lowest at intermediate rates of information 91012.
This effect indicates that curiosity is biologically calibrated to maintain an optimal rate of information absorption. Infants implicitly seek environments that are "just right" - containing enough novelty to be engaging but sufficient predictability to be successfully integrated into existing cognitive schemas 81011. This mechanism ensures that cognitive resources are not wasted on redundant data or incomprehensible noise.
| Theoretical Framework | Primary Mechanism of Curiosity | Neurocognitive Function | Limitations / Critiques |
|---|---|---|---|
| Information Gap Theory | Conscious awareness of missing knowledge creates aversive deprivation or interest. | Motivates targeted search to resolve specific cognitive tension. | Fails to fully explain unprompted novelty-seeking and purely diversive exploration. |
| Active Inference | Continuous minimization of uncertainty (free energy) to optimize generative models. | Drives epistemic foraging and Bayesian model selection (insight). | Highly abstract; empirical translation to human psychosocial behavior is still developing. |
| Goldilocks Effect | Allocation of attention toward intermediate environmental complexity. | Prevents cognitive overload while ensuring continuous schema updating. | Primarily descriptive of attention allocation rather than higher-order epistemic drives. |
Neural Circuitry of Curiosity
The theoretical models of curiosity are physically instantiated in the brain through distinct neural circuits. The neurobiology of curiosity reveals why the acquisition of knowledge is inherently rewarding and how it fundamentally alters memory encoding.
Perceptual Versus Epistemic Curiosity Pathways
Curiosity is not a monolithic neural process; cognitive neuroscience distinguishes between perceptual and epistemic curiosity. Perceptual curiosity is an evolutionarily ancient drive aroused by novel, strange, or ambiguous sensory stimuli 131415. Functional magnetic resonance imaging (fMRI) reveals that the induction of perceptual curiosity activates the anterior insula and the anterior cingulate cortex (ACC) - regions traditionally sensitive to conflict, negative arousal, and homeostatic needs like hunger 141617. The relief of perceptual curiosity, achieved through sensory disambiguation, activates the striatum, providing a reward signal that reinforces the exploratory behavior 1417. Inter-individual variation in trait perceptual curiosity correlates with the strength of anterior insula activation, suggesting highly curious individuals experience stronger negative feelings when confronted with ambiguity 14.
Epistemic curiosity, conversely, is the desire for intellectual information and abstract knowledge 1314. While it shares common pathways with perceptual curiosity, epistemic curiosity is more profoundly linked to the anticipation of reward. It strongly activates the dopaminergic midbrain and the caudate nucleus 1416. Epistemic curiosity transforms learning from a reactive process into a proactive, pleasurable pursuit driven by the anticipation of resolving uncertainty.
Reward Prediction Error and Dopaminergic Signaling
The primary mechanism by which the brain updates its understanding of the world is the Reward Prediction Error (RPE). Historically associated with the receipt of primary rewards, modern neurobiology indicates that midbrain dopaminergic neurons in the ventral tegmental area (VTA) and substantia nigra (SN) fire not for rewards themselves, but for surprises 181925.
An RPE represents the quantitative difference between an expected outcome and the actual outcome. When information is better or more novel than expected, a positive RPE generates a phasic burst of dopamine 1825. This dopamine signal does not solely encode pleasure; it functions as a critical learning signal broadcast across the brain, driving plasticity in the striatum to facilitate reinforcement learning and memory restabilization 1820. In the context of curiosity, the acquisition of targeted information acts as a reward, generating prediction errors that physically rewire synaptic connections to ensure the new data is retained 1925.
Furthermore, local striatal circuits intricately manage this dopaminergic release. Striatal acetylcholine and GABAergic systems modulate dopamine release initiated by midbrain spiking 21. Cholinergic interneurons (ChINs) directly shape dopaminergic output, creating a highly localized regulatory environment where acetylcholine and dopamine dynamics are often anticorrelated during learning tasks 21.
The PACE Framework and Hippocampal Modulation
To synthesize how curiosity affects memory, cognitive neuroscientists Mathias Gruber and Charan Ranganath developed the PACE (Prediction, Appraisal, Curiosity, and Exploration) framework 22232425. The PACE model delineates the specific neural circuitry that transforms a state of curiosity into enhanced memory retention:
- Prediction Error: The cycle begins when the hippocampus or the ACC detects a context-based or information-based prediction error 2224.
- Appraisal: The lateral prefrontal cortex (PFC) evaluates this uncertainty. If the brain determines it has the cognitive resources to resolve the gap, it triggers a curiosity response rather than an anxiety response 2224.
- Curiosity and Dopaminergic Neuromodulation: The lateral PFC signals the dopaminergic midbrain (SN/VTA). This region floods the circuitry with dopamine, creating an anticipatory state of high motivation 22.
- Exploration and Consolidation: Dopamine interacts heavily with the ventral striatum (driving exploration) and the hippocampus. The dopaminergic neuromodulation of the hippocampus significantly enhances both the immediate encoding and the long-term consolidation of the target information 22.
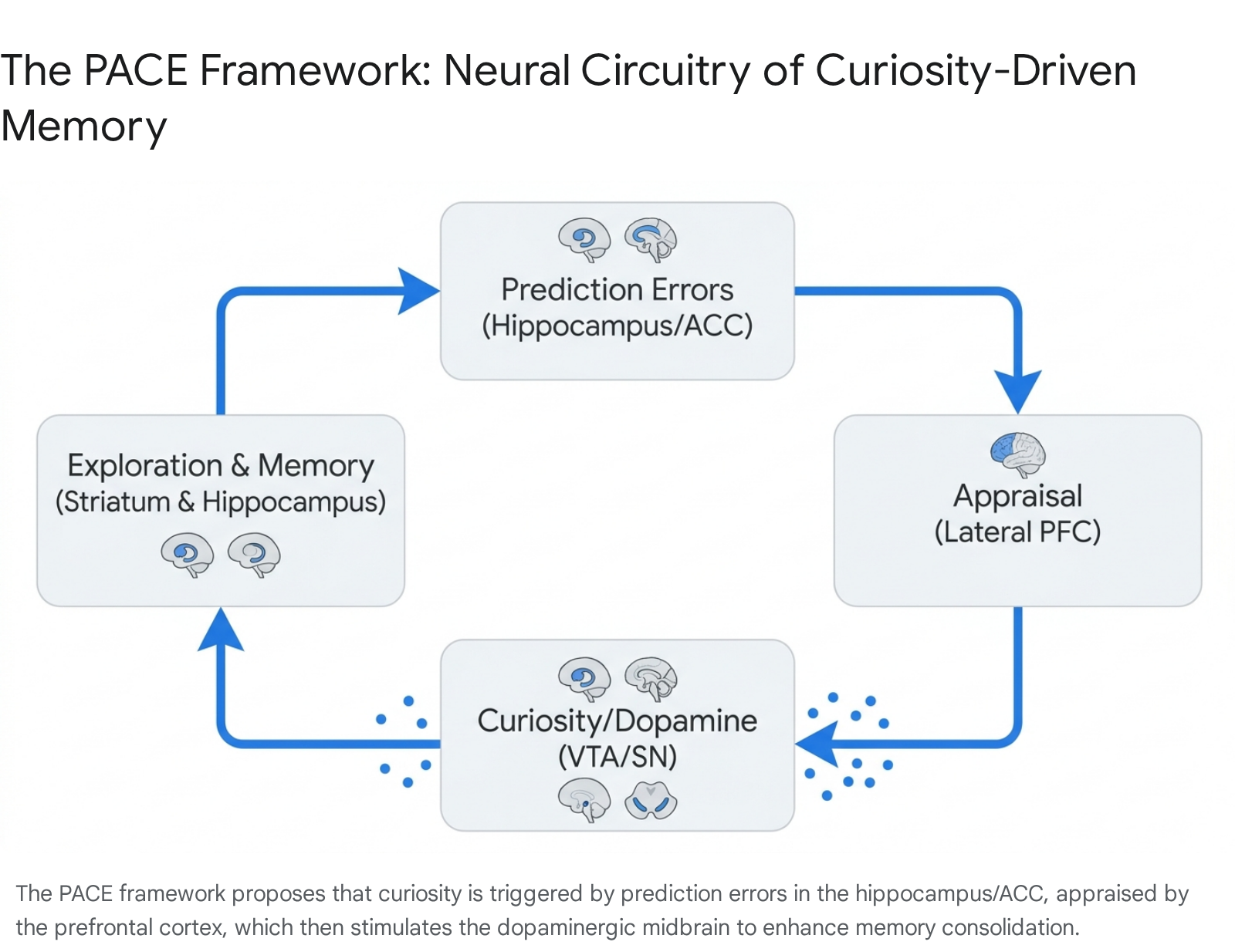
Crucially, the PACE framework explains a phenomenon where curiosity creates a "vortex" of learning. fMRI studies utilizing trivia paradigms found that when subjects were in a state of high curiosity, they not only remembered the answers to the trivia questions better, but they also exhibited enhanced memory for entirely unrelated, incidental information (such as faces) presented during the anticipatory window 25262734.
Recent advancements in understanding event cognition further highlight the hippocampus's role in this process. Functional connectivity between the hippocampus and the neocortical posterior medial network (PMN) peaks at "event boundaries" - moments of transition or uncertainty in naturalistic observation 3528. Hippocampal-PMN connectivity at these boundaries strongly predicts the subsequent recall of complex narrative events, suggesting that the brain segments information based on moments of resolved uncertainty 28. Furthermore, researchers are actively investigating the sub-second rhythmicity of memory behavior, mapping optimal and suboptimal phases of attention and memory encoding to specific phases of hippocampal theta oscillations 29.
Memory Interference and Complex Scholastic Encoding
While the "vortex" effect of curiosity suggests broad educational applications, recent neurobiological research reveals distinct limitations. Curiosity does not universally enhance all forms of learning.
Studies by Stare, Dunsmoor, and colleagues tested the limits of incidental memory enhancement by replacing simple visual stimuli (faces) with complex, unrelated scholastic facts presented during states of high trivia-induced curiosity 3031. Contrary to the standard PACE predictions, high curiosity interfered with the encoding of these complex scholastic facts, resulting in poorer memory retention 30.
This interference suggests a resource competition model within working memory. When the brain is highly aroused and awaiting the resolution of a specific, high-value prediction error, the cognitive load is substantial. While simple stimuli like faces can be passively encoded via dopaminergic diffusion, complex semantic information requires active executive processing. Because executive resources are already allocated to the primary object of curiosity, concurrent complex learning is degraded 3031. Thus, curiosity operates as an exceptionally powerful, yet highly targeted, cognitive resource rather than a universal learning enhancer.
Metabolic Regulation of Exploratory Behavior
The shift between learning (exploration) and relying on prior knowledge (exploitation) is heavily governed by the brain's metabolic state. Exploration is an energetically expensive neurological process, tightly regulated by systemic energy homeostasis.
Energy Homeostasis and Striatal Foraging Circuits
Organismic states of energy depletion profoundly alter exploratory behavior. Fasting and hunger reliably increase risk-taking and exploratory foraging, overriding standard risk aversion to ensure survival 3233. The neural coordination of this metabolic shift originates in the hypothalamus, specifically via agouti-related peptide (AgRP) neurons, commonly known as "hunger neurons" 3233.
These AgRP neurons project polysynaptically through the lateral hypothalamus, central amygdala, and periaqueductal gray to dopaminergic neurons projecting to the tail of the striatum (TOS). In states of caloric depletion, this circuit suppresses standard dopamine signaling in the TOS, which structurally alters the animal's behavior to favor wide-ranging exploration over localized exploitation 3233. Furthermore, extracellular dopamine dynamics differ wildly across the striatum; the ventral striatum maintains persistent, minute-long dopamine tone linked to broad motivational outputs, whereas the dorsal striatum exhibits rapid, transient fluctuations 2134. This indicates that the brain's metabolic reserves directly tune dopaminergic circuits to calibrate the cost-to-benefit ratio of curiosity.
Brain Insulin Signaling and Reward Salience
Insulin, traditionally viewed strictly as a peripheral metabolic hormone, plays a critical, distinct role in the central nervous system. Brain insulin signaling operates independently of blood glucose levels and is heavily concentrated in regions associated with memory and reward, such as the hippocampus and striatum 353637.
In the striatum, insulin acts as a powerful reward signal. An elevation in brain insulin promotes increased stimulated dopamine release by acting on cholinergic interneurons and nicotinic acetylcholine receptors on dopamine axons 213738. This insulin-dopamine axis is essential for flavor-nutrient learning and general motivational salience 37. Furthermore, insulin in the hippocampus stimulates dendritic spine formation and long-term potentiation (LTP) via PI3K/Akt signaling pathways, creating the cellular foundation for memory encoding 3639.
Consequently, metabolic pathologies fundamentally impair curiosity and cognition. Brain insulin resistance (BIR) - increasingly recognized as a hallmark of aging and Alzheimer's disease - deprives the prefrontal cortex and hippocampus of necessary metabolic signaling. This leads to diminished synaptic plasticity, depressive-like behaviors, hyperphagia, and a stark reduction in exploratory drives 35394041. The metabolic stress forces a shift into a hypervigilant, sympathetic-dominant state that precludes complex learning 40. Promising modern therapeutics, such as GLP-1 receptor agonists, are currently being investigated not just for weight loss, but for their ability to restore mitochondrial structure, improve oxidative phosphorylation, and re-regulate dopaminergic reward salience in the ventral striatum 41.
Mitochondrial Neoteny and Neurodevelopment
At the cellular level, the tempo of neurodevelopment and the capacity for learning are set by mitochondrial metabolism. The human brain consumes 20% of the body's resting energy, demanding highly active oxidative phosphorylation (OxPhos) 4243.
Recent comparative neurobiology reveals that human cortical neurons exhibit significantly slower mitochondrial development and lower metabolic activity in early life compared to other mammals 44. This uniquely prolonged mitochondrial immaturity dictates the species-specific neoteny of the human brain. By delaying the stabilization of neural circuits, human mitochondria allow for an extended period of childhood neuroplasticity and curiosity-driven exploration 44. Genetic pathways governing mitochondrial function, such as those involving the SLC33A1, SLC13A5, and SLC25A1 genes which transport acetyl-CoA to the endoplasmic reticulum, are vital for this developmental pacing. Mutations in these metabolic pathways lead to premature neuronal maturation and altered exploratory behaviors, often manifesting as autism-like traits and repetitive behavior in animal models 45. In adulthood, chronic stress and aging induce mitochondrial dysfunction, shifting cellular energy away from restorative learning processes and toward allostatic survival responses, further suppressing epistemic curiosity 4243.
| Metabolic Regulator | Primary Locus of Action | Impact on Curiosity and Exploration | Pathological Outcomes |
|---|---|---|---|
| AgRP "Hunger" Neurons | Hypothalamus to Tail of Striatum (TOS) | Suppresses TOS dopamine to trigger broad foraging and risk-taking. | Maladaptive feeding behaviors if chronic. |
| Brain Insulin | Striatum and Hippocampus | Enhances dopamine release via ChINs; promotes dendritic spine formation. | Brain Insulin Resistance (BIR) correlates with cognitive decline and apathy. |
| Mitochondrial Metabolism | Cortical Neurons (Global) | Sets tempo of neurodevelopment; delayed OxPhos allows extended plasticity. | Dysfunction forces energy diversion to allostasis, halting learning. |
Adult Cognitive Aging and Information Processing
As the brain ages, biological and metabolic shifts alter how individuals interact with information. The psychological manifestation of this transition is known as the exploration-exploitation tradeoff, representing a shift from seeking novel information (exploration) to relying on accumulated knowledge (exploitation) 464748.
The Exploration and Exploitation Tradeoff
The tension between exploring unknown possibilities and exploiting known rewards is pervasive across biological systems 4647. In young adulthood, cognitive control processes facilitate active exploration to map the environment. However, the cognitive control hypothesis suggests that as executive functions naturally decline with age, older adults transition toward an exploitative mental mode 4648.
Pharmacological studies reveal that this tradeoff is tightly governed by systemic catecholamine levels. Systemically increasing dopamine activity decreases exploration by increasing "stickiness" or choice precision, while decreasing dopamine activity increases exploration 49. Norepinephrine also modulates exploration, though it does so by altering outcome sensitivity in a sex-dependent manner 49.
Structural Neuroplasticity and Synaptic Shifts
Historically, it was assumed that neuroplasticity ceased in midlife. Modern neuroscience refutes this, demonstrating that adult neurogenesis continues in hippocampal memory centers well into old age 5051. However, the efficiency and structural nature of plasticity shift.
With advancing age, the dorsolateral prefrontal cortex experiences a significant loss (up to 46%) of "thin spines" - the highly plastic dendritic spines responsible for rapid, novel learning and cognitive flexibility 52. Conversely, larger "mushroom spines," which stabilize established networks and crystallized knowledge, remain largely intact 52. This structural alteration biologically biases the older brain toward exploitation. Reductions in synaptic turnover and slower neurotransmitter signaling make the encoding of entirely novel information slower and more energetically demanding, thereby favoring the utilization of preexisting, highly efficient distributed neural networks 5152.
Trait Versus State Curiosity in Later Life
This neurobiological shift results in observable changes in psychological curiosity profiles. Longitudinal data indicates that general trait curiosity (a pervasive desire to seek out new information across domains) declines across the adult lifespan 625354.
However, state curiosity - a situational, targeted interest in specific information - often increases sharply after middle age and remains robust into late adulthood 6254. This aligns with Socioemotional Selectivity Theory, which posits that as time horizons shorten, older adults optimize their cognitive resources, shedding broad, indiscriminate exploration in favor of highly selective, emotionally meaningful knowledge acquisition 4854. The shift to heuristic reliance is not strictly a deficit; it is an efficient evolutionary adaptation that maintains high decision-making accuracy without massive metabolic expenditure 4651. Maintaining this targeted state curiosity is neuroprotective, with profound disinterest and apathy acting as early clinical biomarkers for cognitive decline and Alzheimer's disease 6255.
Educational Frameworks and Curiosity Retention
The neurobiology of learning establishes that human brains are naturally engineered to seek out intermediate complexity and process it via intrinsic dopaminergic reward pathways. However, formal educational environments can either leverage or suppress these biological imperatives.
Impacts of Standardized Assessment Models
The dominant paradigm in global education relies heavily on standardized testing. While standardized models offer administrative efficiency, scalability, and objective comparability, they frequently conflict with the neurobiology of curiosity 566757.
Standardization inherently prioritizes the exploitation of predetermined facts over the exploration of novel concepts. By narrowing curricula to match rigid test outcomes, these systems rely on rote memorization and immediate recall 675859. From a neurobiological perspective, this approach bypasses the prediction-error mechanisms of the hippocampus and the intrinsic dopaminergic reward systems. Instead of allowing learners to formulate hypotheses and experience the biochemical reward of resolving an information gap, standardizing knowledge reduces learning to a compliance task, shifting motivation from intrinsic to extrinsic 6757. Furthermore, the chronic stress associated with high-stakes testing elevates systemic glucocorticoids, which can impair hippocampal function and shift the brain out of a curious state into a sympathetic-dominant survival mode 4356.
Inquiry-Based Learning and Systemic Comparisons
Conversely, educational systems that de-emphasize rigorous standardization often align more closely with cognitive neuroscience. The Finnish education system serves as a prominent modern example. Finland minimizes standardized testing (employing only one mandatory exam at the end of upper secondary school), shortens formal school hours to allow for adequate rest, and emphasizes teacher autonomy and inquiry-based learning 606162.
Inquiry-based models initiate learning with a question rather than a fact, actively leveraging the Information Gap Theory to induce a state of mild cognitive deprivation that intrinsically motivates the student 160. By allowing students to explore at their own pace, these systems respect the Goldilocks Effect, enabling learners to interact with material at a complexity level that is optimally engaging for their specific cognitive baseline 1060.
However, this model is subject to intense academic debate. Critics note that while Finland historically performed exceptionally well, its scores on the international PISA benchmarks have slipped in recent years 6364. Some researchers argue that Finland's initial success was a byproduct of its older, more traditional centralized system, and that highly rigorous East Asian systems - which lean heavily on continuous, competitive repetition - produce consistently superior measurable outcomes 6364. Conversely, global analysts warn against mimicking East Asian models entirely, arguing that uncompromising, test-driven milieus may sacrifice long-term creative flexibility and well-being for short-term metric gains 6364.
Indigenous Pedagogies and Holistic Paradigms
Alternative, non-Western educational frameworks offer sophisticated models for embedding curiosity into learning by centering contextual relevance. Indigenous pedagogies, such as the Australian Aboriginal "8-Ways of Learning" framework, explicitly reject the compartmentalized, linear delivery of isolated facts 656667.
The 8-Ways framework focuses on eight interconnected pedagogies: narrative-driven learning (Story Sharing), spatial visualization (Learning Maps), non-verbal kinesthetic skills, the use of symbols and metaphors, place-based relevance (Land Links), non-linear lateral thinking, deconstruct/reconstruct modeling, and connectedness to community 666779.
Neurobiologically, this approach is highly effective for retention. By embedding new information within complex community narratives and physical environments, indigenous pedagogies generate rich, multi-sensory prediction errors that engage wide networks of the brain 6768. The "Deconstruct/Reconstruct" methodology - watching a whole process before breaking it down into component parts - directly supports the brain's natural active inference processes, allowing students to build complete generative models rather than struggling to integrate disparate, isolated variables 6681. Integrating these pedagogical models validates multiple ways of knowing and directly aligns with the brain's evolved preference for contextually grounded, relationally rich exploration 697071.
Conclusion
The science of curiosity reveals that the brain does not merely store information; it metabolically requires and actively seeks it. The reward of learning is orchestrated by an elegant neurobiological sequence where the detection of uncertainty generates prediction errors, prefrontal appraisal authorizes exploration, and dopaminergic cascades secure the resulting memory in the hippocampus. Theoretical frameworks like Active Inference and the Information Gap Theory demonstrate that the minimization of uncertainty is a fundamental driver of intelligence.
As the organism ages, systemic constraints - ranging from localized brain insulin resistance to mitochondrial down-regulation and shifting dendritic spine morphology - force a strategic pivot. The brain transitions from the energy-intensive exploration characteristic of youth to the highly efficient exploitation of established knowledge. While general trait curiosity may wane, selective, state-based curiosity remains a vital neuroprotective mechanism against cognitive decline. Ultimately, the environments society constructs, from formal educational curricula to the design of pedagogical frameworks, must be calibrated to these biological realities. Systems that artificially restrict exploration or rely entirely on rote standardization suppress the brain's natural reward mechanisms, while those that embrace optimal complexity, active inquiry, and contextual relevance harness the profound, innate human drive to understand the world.