Neural substrates of loss aversion
Loss aversion - the empirical observation that the psychological impact of a loss outweighs that of a mathematically equivalent gain - has served as a foundational pillar of behavioral economics and cognitive psychology since its formalization in Prospect Theory. Traditionally, behavioral models have quantified this asymmetry using a loss aversion coefficient ($\lambda$), estimating that losses loom approximately 1.5 to 2.5 times larger than gains 123. For decades, researchers sought a singular, localized neural generator for this phenomenon, postulating the existence of a dedicated "fear" or "aversion" center in the brain. However, contemporary cognitive neuroscience, utilizing functional magnetic resonance imaging (fMRI), focal lesion studies, and computational neurobiology, has dismantled this localizationist perspective.
The asymmetry described by Kahneman and Tversky does not reside in a single anatomical structure. Rather, it is an emergent property arising from the continuous, dynamic integration of multiple neural systems. This architecture includes subcortical threat detection systems, striatal reward prediction mechanisms, the biophysical constraints of midbrain dopaminergic signaling, and cortical networks responsible for subjective value integration and executive control 3456. Furthermore, recent advancements in drift-diffusion modeling and cross-cultural neuroimaging have challenged the universality of loss aversion, demonstrating that the phenomenon is highly contingent on stake magnitude, contextual framing, and systemic structural biases 78910. This report systematically examines the distributed neural substrates, neurochemical mechanisms, and computational models that govern reference-dependent value processing.
The Amygdala and Aversive Valuation
Early neuroeconomic hypotheses posited that loss aversion was primarily driven by negative affective responses, making the amygdala - a medial temporal lobe structure critical for threat detection, fear conditioning, and autonomic arousal - a primary candidate for its neural origin.
Causal Evidence from Focal Lesion Studies
The most definitive evidence establishing the amygdala's causal role in behavioral loss aversion emerges from studies of rare human patients with complete bilateral focal lesions of the amygdala (e.g., patients resulting from Urbach-Wiethe disease). In standardized experimental economics paradigms, participants evaluate mixed gambles characterized by equal probabilities of winning a larger amount or losing a smaller amount (e.g., a 50% chance to win $\$15$ or lose $\$10$). Neurotypical individuals robustly reject gambles with positive expected values due to an inherent aversion to the potential loss 10.
When subjected to these mixed gambles, patients with bilateral amygdala damage demonstrate a profound and specific disruption in choice behavior. While demographically matched healthy control participants exhibit standard loss aversion coefficients ($\lambda \approx 1.52$ to $1.76$), amygdala-lesioned patients display an almost complete elimination of loss aversion, yielding coefficients approaching strict loss neutrality ($\lambda \approx 0.76$ to $1.06$) 10.
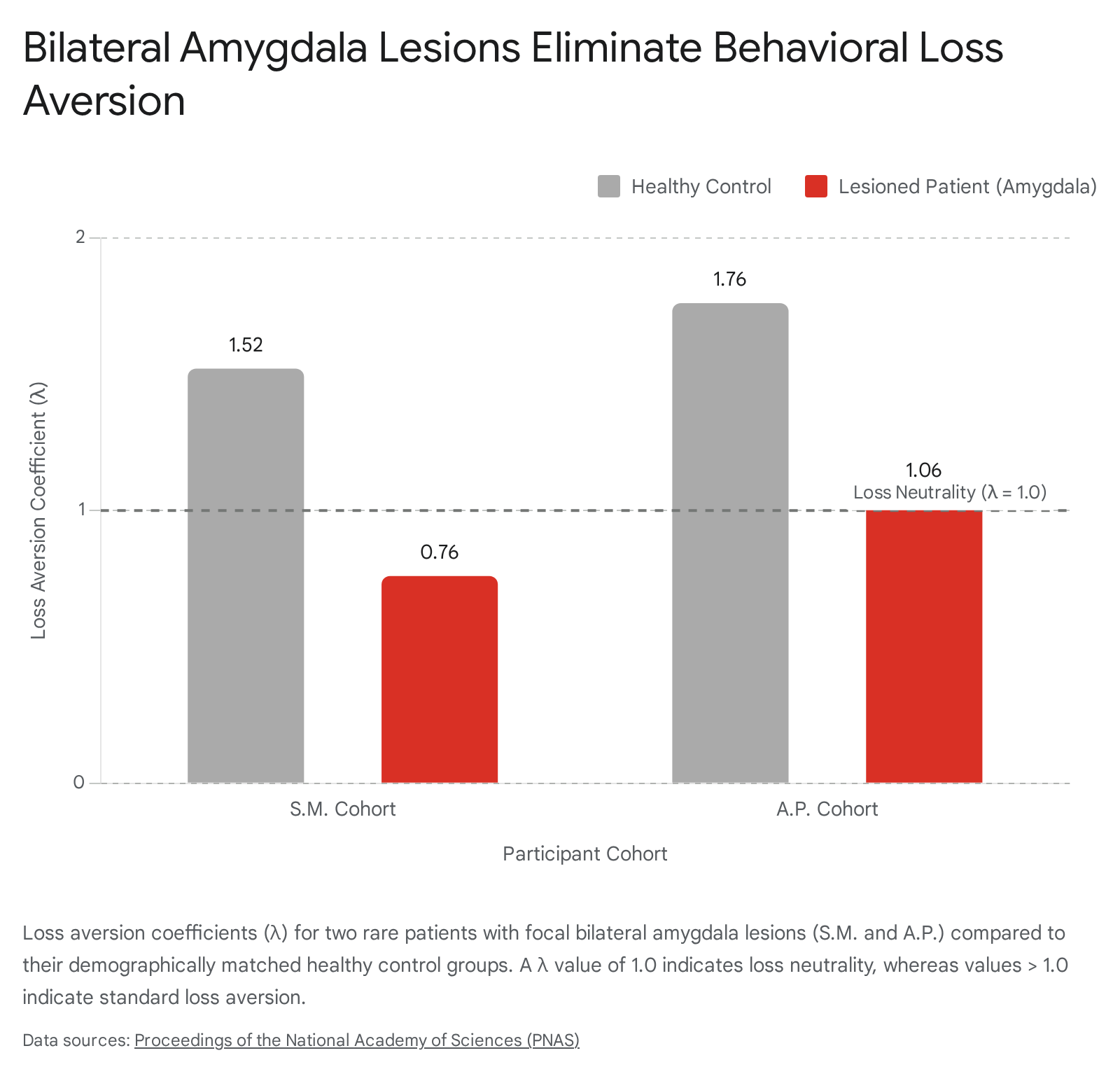
Crucially, this disruption is not indicative of a generalized deficit in risk assessment or value computation. These patients retain a normal, mathematically sound ability to adjust their choices based on changes in expected value and outcome variance. Their impairment is highly selective: they fail to generate the asymmetric affective penalty typically assigned to prospective financial losses, suggesting the amygdala operates as an essential cautionary brake on behavior involving deleterious outcomes 510.
Functional Neuroimaging and Emotional Regulation
In neurotypical populations, blood oxygen level-dependent (BOLD) responses measured via fMRI reinforce the amygdala's role in scaling loss severity. During the decision-making phase, amygdala activation increases parametrically with the magnitude of potential losses 3511. The strength of this neural response directly predicts individual differences in behavioral loss aversion, linking the physiological arousal response directly to economic choice.
This amygdala-mediated mechanism is also highly sensitive to top-down emotional regulation. When individuals utilize cognitive reappraisal strategies - deliberately reframing the context of a gamble to minimize emotional reactivity - they exhibit concurrent reductions in both behavioral loss aversion and loss-related amygdala activation 25. This covariation confirms that the asymmetric weighting of losses is not a strictly mathematical computation, but rather an affective one, relying on the amygdala to translate potential financial deprivation into a biological signal of threat or negative arousal.
Ventral Striatum and the Unified Valuation Network
If the amygdala provides an affective penalty signal for losses, the striatum operates as the primary engine for computing expected utility and reward prediction. Early neuroeconomic frameworks hypothesized a dual-system architecture: an appetitive system (centered in the striatum) that exclusively processed gains, and an independent aversive system (centered in the amygdala and anterior insula) that processed losses.
Neural Loss Aversion in the Striatum
Subsequent high-resolution fMRI studies overturned the strict dual-system hypothesis, identifying instead a unified valuation system primarily located within the ventral striatum (VS) and the ventromedial prefrontal cortex (vmPFC). In this unified framework, activity in the VS encodes both potential gains and potential losses on a single, bidirectional continuum. Striatal BOLD responses increase parametrically as the magnitude of potential gains increases, and decrease parametrically as the magnitude of potential losses increases 351011.
Crucially, this system exhibits an inherent functional asymmetry. The slope of neural deactivation for increasing losses is significantly steeper than the slope of neural activation for mathematically equivalent increasing gains 3111213. This phenomenon, termed "neural loss aversion," functions as the direct neural correlate of the behavioral $\lambda$ coefficient. Individuals who exhibit severe behavioral loss aversion display correspondingly steeper asymmetric BOLD responses in the ventral striatum, confirming that Kahneman's asymmetry is deeply embedded within the brain's fundamental reward circuitry.
Dynamic Reference Point Switching
The striatal encoding of value is not absolute; it is highly dependent on situational reference points. Investigations into motor task performance under financial pressure have revealed that the VS dynamically shifts its framing between incentive presentation and task execution.
When a participant is initially presented with a high monetary incentive for successfully completing a skilled task, the VS encodes the incentive as a potential gain, marked by robust neural activation 14. However, as the participant begins to execute the highly demanding task, cognitive framing shifts. Striatal activity rapidly inverts, switching from activation to deactivation, effectively encoding the scenario no longer as the pursuit of a reward, but as the active avoidance of a substantial loss if they fail 1415.
This reframing mechanism explains the phenomenon of "choking under pressure." Decrements in motor performance under high financial stakes correlate strongly with these mid-task striatal deactivations, and the magnitude of the performance drop is directly predicted by an individual's baseline behavioral loss aversion trait 1415. Thus, the VS translates prospective incentives into performance-modulating signals that are inherently skewed by a loss-averse reference frame.
Dopaminergic Mechanisms of Asymmetric Encoding
The fundamental driver of striatal valuation is the neurotransmitter dopamine, primarily synthesized in the Ventral Tegmental Area (VTA) and Substantia Nigra pars compacta (SNc). The computation of value relies on Reward Prediction Errors (RPEs) - the mathematical discrepancy between expected outcomes and actual outcomes, serving as the biological instantiation of temporal difference (TD) learning models 16171819. The roots of loss aversion can be traced directly to the biophysical and anatomical constraints of these midbrain dopamine neurons.
Firing Rate Asymmetries: Spikes versus Dips
Dopaminergic neurons typically maintain a low, steady tonic baseline firing rate of approximately 4 to 6 spikes per second (Hz) 202122. When an unexpected reward is encountered (generating a positive RPE), these neurons fire a rapid burst of action potentials, temporarily increasing their rate up to 20, 30, or even 50 Hz. Conversely, when an expected reward is omitted, or an unexpected loss occurs (generating a negative RPE), the neurons pause their firing, resulting in a "dip" 17232425.
This physiological reality creates an unavoidable biological bound. The dynamic range for signaling a positive RPE is vast - expanding from 5 Hz up to the physiological maximum of the neuron. However, the dynamic range for signaling a negative RPE is strictly floored at 0 Hz 1922. Consequently, a mathematical doubling of a positive reward can elicit a doubling of the neuronal spike rate, but a mathematical doubling of a severe loss cannot double the magnitude of a neuronal dip if the neuron is already completely silent.
Downstream Post-Synaptic Scaling
If raw dopamine firing rates are fundamentally truncated by a zero-bound floor, the brain requires downstream mechanisms to accurately represent severe losses and drive avoidance behavior. This is achieved through asymmetric post-synaptic receptor sensitivities in the striatum.
Dopamine acts primarily on two receptor families: D1 receptors (which facilitate the direct, "Go" pathway) and D2 receptors (which suppress the indirect, "No-Go" pathway). Research indicates that decreases in baseline dopamine (the dip) cause exponentially greater disinhibition of D2-receptor-bearing neurons than equivalent increases in dopamine (the burst) cause excitation in D1-receptor-bearing neurons 2022. Thus, a relatively small physiological pause in midbrain dopamine release is amplified at the synaptic level into a massive signal to halt action and avoid the loss. This disparity in receptor efficacy biologically enforces the behavioral reality that losses exert a heavier subjective weight than gains.
Distributional Reinforcement Learning
Recent advancements in computational neuroscience have revealed that dopamine neurons do not broadcast a single, monolithic prediction error. Instead, the brain employs a mechanism known as Distributional Reinforcement Learning. Different subpopulations of midbrain dopamine neurons maintain distinct baseline firing rates and varying asymmetric scaling factors for positive versus negative errors 162022.
Some dopamine neurons operate as "optimists," scaling positive RPEs more steeply than negative ones. Conversely, other neurons operate as "pessimists," reacting with disproportionate severity to negative RPEs. By maintaining a diverse population of neurons with differing scaling asymmetries, the brain learns a complete probability distribution of future outcomes (represented mathematically as expectiles), rather than a single mean expected value 1622. Individual differences in trait loss aversion are theorized to stem from variations in the population density of these neuronal types; an individual with a higher proportion of "pessimistic" dopamine neurons will inherently compute a skewed, loss-averse behavioral policy.
| Mechanism of Asymmetry | Neurobiological Constraint | Functional Consequence on Valuation |
|---|---|---|
| Spike/Dip Floor Limit | Tonic baseline firing is ~4-6 Hz. Negative prediction errors cannot drop below 0 Hz, capping the raw signal amplitude 202225. | Demands downstream computational adaptation; raw firing rates alone underestimate severe losses. |
| Receptor Efficacy (D1R vs. D2R) | Drops in dopamine cause rapid unbinding at D2 receptors, amplifying the physiological impact of a "pause" 2022. | A biological amplification of negative outcomes, driving aversive learning and status quo maintenance. |
| Distributional Reinforcement Learning | Sub-populations of dopamine neurons possess varying ratios of learning rates for positive versus negative prediction errors 1620. | Generates diverse expectiles of reward; a structural skew toward "pessimistic" neurons biases choice toward loss aversion. |
Ventromedial Prefrontal Cortex in Value Integration
While the striatum and amygdala process the immediate predictive and affective components of gambles, these disparate signals must be integrated to facilitate complex decision-making. The ventromedial prefrontal cortex (vmPFC) fulfills this role, computing the subjective value of available options and translating them into a "common neural currency" that allows individuals to weigh differing types of rewards and punishments 11262728.
Impairments in Subjective Valuation and Utility Curvature
The necessity of the vmPFC in rational economic choice is highlighted by studies of patients with focal bilateral lesions to this region. These patients demonstrate profound deficits in value-based decision-making, frequently engaging in paradoxical risk-seeking behaviors and failing to properly weigh future consequences against immediate prospects 27293031.
When the choice behavior of vmPFC-lesioned patients is modeled using Cumulative Prospect Theory (CPT), a distinct computational phenotype emerges that contrasts sharply with the phenotype of amygdala-lesioned patients. While amygdala damage primarily flattens the loss aversion parameter ($\lambda$), vmPFC damage fundamentally alters the curvature of the utility function for both gains ($\alpha$) and losses ($\beta$) 2731. Patients with vmPFC lesions exhibit significantly steeper utility curves, indicating a severe, generalized insensitivity to outcome magnitudes. Concurrently, their probability weighting functions become notably more linear (higher $\gamma$), reflecting a failure to appropriately weight low-probability risks 2731.
Although these patients display a trend toward reduced loss aversion, their primary deficit lies in the failure to integrate the absolute magnitude of the gamble and to exert top-down executive inhibition over impulsive reward-seeking behavior 272931.
Causal Neuromodulation
The causal role of the vmPFC in mitigating loss aversion and framing effects has been further substantiated by non-invasive brain stimulation studies. Excitatory transcranial direct current stimulation (tDCS) or magnetic stimulation applied to the vmPFC effectively alters risk-taking behavior. Excitation of the vmPFC decreases risk-seeking when the probability of a loss is high, while simultaneously decreasing irrational risk-aversion when the probability of a gain is high. By artificially enhancing the regulatory capacity of the vmPFC, individuals make choices with higher overall expected values and demonstrate a heightened resistance to cognitive biases and maladaptive framing effects 2829.
Network Connectivity and Psychiatric Implications
Loss aversion is an inherently plastic computation that fluctuates based on emotional state, acute stress, and situational context. These dynamic shifts are governed by real-time alterations in functional connectivity between the frontostriatal and amygdala-striatal networks.
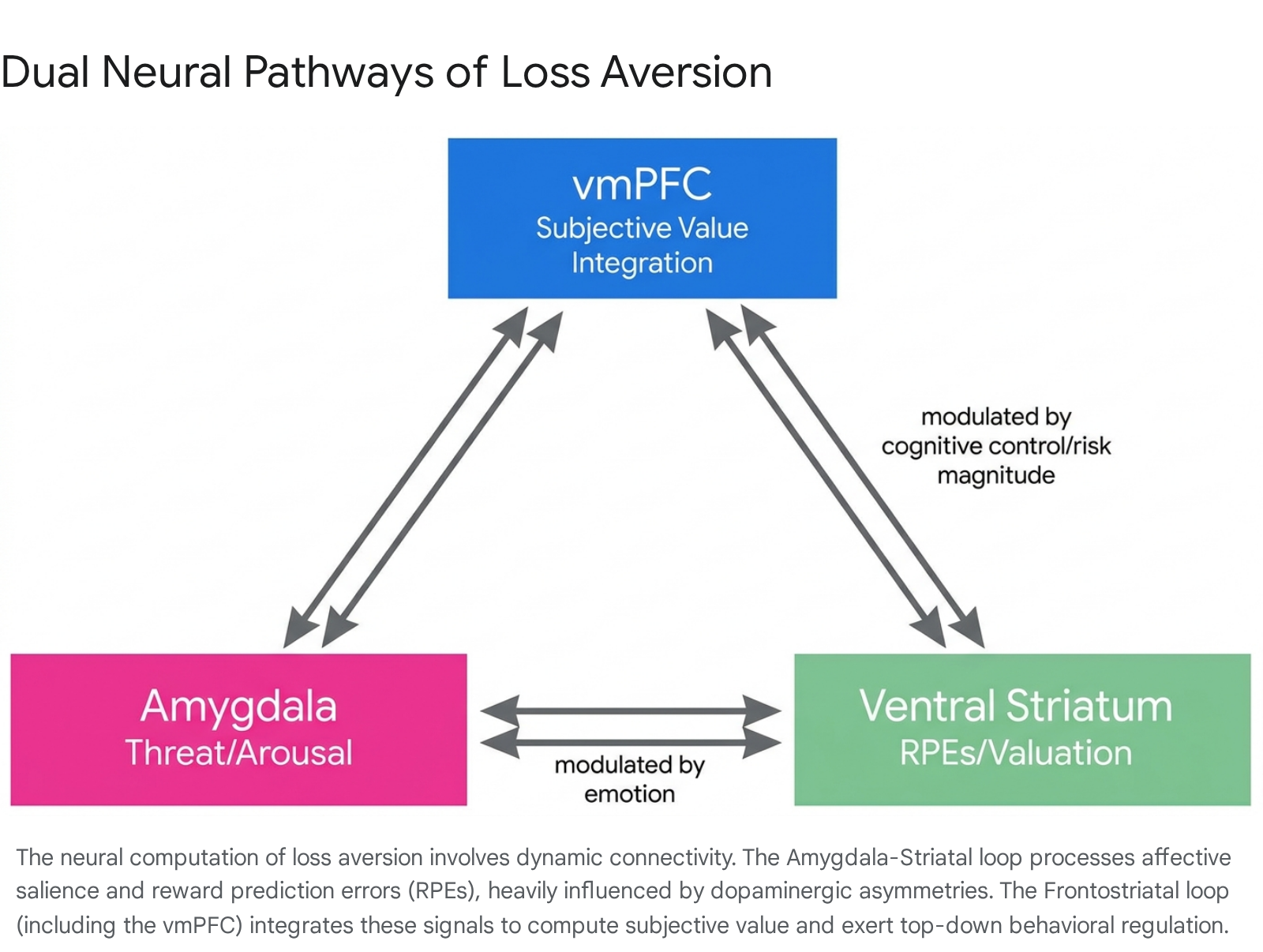
Amygdala-Striatal Coupling
The degree to which an individual overweighs a loss depends heavily on functional coupling between the amygdala and the ventral striatum. In fMRI paradigms incorporating emotional priming - such as presenting participants with subliminal fearful faces prior to an economic choice - significant shifts in valuation occur. Specifically, individuals with low baseline trait anxiety exhibit a transient but marked increase in behavioral loss aversion following exposure to threat cues 1213.
This context-induced spike in loss aversion is statistically mediated by an acute increase in amygdala-striatal functional connectivity 111213. In this state, the amygdala shifts the baseline calibration of the striatal valuation system, integrating external threat cues with the mathematical expected value of the gamble to engage adaptive, harm-avoidant behaviors.
Pathological Disruptions in Network Connectivity
The structural and functional integrity of frontostriatal circuits - linking the prefrontal cortex (including the vmPFC and dorsolateral prefrontal cortex) to the basal ganglia - is critical for the regulation of standard loss-averse behavior. In psychiatric conditions marked by poor decision-making, excessive risk-taking, and heightened impulsivity, severe dysregulation within these networks is consistently observed.
Systematic reviews of risk processing in psychiatric cohorts identify frontostriatal dysregulation as a transdiagnostic feature, typically characterized by prefrontal hypoactivation paired with striatal hyperreactivity 3233. * Bipolar Disorder: During the manic phase, individuals exhibit severe disruptions in ventral prefrontal-subcortical connectivity, resulting in exaggerated reward-seeking and a stark reduction in loss aversion. Longitudinal studies indicate that clinical remission correlates with the normalization of connectivity between the medial prefrontal cortex, thalamus, and inferior frontal gyrus 323334. * Major Depressive Disorder (MDD): MDD frequently involves pronounced anhedonia and altered risk processing, linked to aberrant functional and effective connectivity between the ventral striatum and the medial prefrontal cortex. This connectivity blunts typical reward valuation, distorting standard loss aversion metrics 323335. * Behavioral Addictions: Conditions such as Internet Gaming Disorder (IGD) and Short-Video Addiction (SVA) are associated with hypersensitivity within the mesolimbic reward salience network. Addicted individuals display diminished loss aversion, increased impulsivity, and altered amygdala-striatal morphology, prioritizing immediate gratification over the avoidance of long-term negative consequences 36373839.
| Clinical Condition | Primary Neural Disruption | Impact on Loss Aversion and Risk Behavior |
|---|---|---|
| Bipolar Disorder (Mania) | Ventral prefrontal hypoactivation; striatal hyperreactivity 323334. | Drastic reduction in loss aversion; excessive risk-seeking; failure to weight negative consequences. |
| Major Depressive Disorder | Blunted ventral striatum activity; altered vmPFC-striatum effective connectivity 323335. | Anhedonia; disrupted subjective valuation; altered baseline risk sensitivity. |
| Behavioral Addictions (IGD/SVA) | Hypersensitivity in reward salience networks; DMN hyper-connectivity 36373839. | Diminished loss aversion; temporal discounting; preference for immediate over delayed rewards. |
Computational Decomposition and Behavioral Critiques
While neuroimaging and lesion studies have effectively mapped the brain's asymmetric valuation networks, a growing contingent of behavioral economists and cognitive psychologists argue that the fundamental premise of ubiquitous loss aversion is flawed.
The Magnitude Dependence Critique
Researchers such as Gal and Rucker (2018) assert that the principle of loss aversion has achieved a "rule-like" status that outpaces the empirical evidence 4942. They argue that many phenomena historically attributed to an asymmetric value function - such as the status quo bias or the endowment effect - can be more parsimoniously explained by cognitive inertia or a general bias against taking action, rather than an explicit overweighting of losses 3479.
Extensive empirical re-evaluations indicate that loss aversion is fundamentally magnitude-dependent. In experimental designs utilizing very small stakes (e.g., $6 gains and losses), participants frequently fail to exhibit loss aversion, instead demonstrating loss-gain neutrality or even gain-seeking behaviors (averaging $\lambda < 1.0$) 34041. Strong, classic loss aversion ($\lambda \approx 1.5$ to $2.0$) reliably emerges only when absolute financial stakes increase past a subjective threshold of meaningful consequence 184042. This evidence directly challenges the core tenet of prospect theory that the value function is universally steeper for losses across all micro and macro scales.
Decomposing Valuation and Pre-Valuation Biases
To reconcile robust fMRI findings of "neural loss aversion" with behavioral data highlighting context-dependent fragility, computational neuroscientists have applied drift-diffusion models to separate decision-making into discrete temporal and cognitive phases. This decomposition successfully identifies two independent psychological mechanisms that contribute to what is broadly observed as loss aversion:
- Valuation Bias: This mechanism aligns with the traditional prospect theory definition, wherein the subjective utility of a loss is mathematically overweighted relative to a gain during the evaluation phase of decision-making. This bias correlates with localized BOLD responses in the reward circuitry, specifically the ventral striatum and vmPFC 79.
- Pre-Valuation Bias: This mechanism constitutes a structural response bias favoring the rejection of gambles (maintaining the status quo) that occurs prior to the actual cognitive calculation of the gamble's expected value. This bias reflects an initial heuristic expectation regarding the desirability of the choice. It is marked by different physiological markers - such as distinct pupil dilation patterns - and functions independently of the brain's core valuation centers 7894344.
These models suggest that the widespread observation of behavioral loss aversion is frequently a conflation. It is a mixture of true neural value asymmetry (which activates robustly only at higher stakes or under stress) and a baseline heuristic to reject complex or ambiguous changes to one's current state 8943.
Contextual and Cross-Cultural Modulators
The assumption that "losses loom larger than gains" constitutes an invariant, hardwired rule of human neurobiology is further challenged by cross-cultural psychological and neuroimaging studies. The vast majority of foundational loss aversion literature relies heavily on participants from Western, Educated, Industrialized, Rich, and Democratic (WEIRD) societies 4546.
When identical risky choice and endowment effect paradigms are administered globally, significant behavioral variance emerges. Studies involving East Asian populations (e.g., China, Japan, Korea) often document substantially weaker endowment effects and altered loss aversion baselines compared to North American and European samples 104748. These behavioral divergences are heavily mediated by disparate cultural frameworks regarding self-concept, regulatory focus (promotion versus prevention), and cognitive consistency 1048. The divergence is even more stark in studies of non-market populations; for example, the Hadza hunter-gatherers in Tanzania exhibit essentially no endowment effect or traditional loss aversion in domains lacking deep integration into modern market economies 10.
Despite these behavioral shifts, the underlying neural architecture appears remarkably conserved. Cross-cultural fMRI data demonstrate that the core affective and valuation networks - specifically the amygdala, insula, and striatum - are universally recruited when evaluating gains and losses across Western and non-Western cohorts 4649. What differs across cultures is the calibration of the set-point. Environmental volatility, societal market integration, and cultural attitudes toward risk modulate the resting functional connectivity within these neural networks, ultimately shifting the psychological threshold at which a potential loss triggers a biologically asymmetric response 464750.
The neuroscience of loss aversion reveals a highly dynamic, distributed architecture rather than a static, localized bias. At the cellular level, the asymmetry is grounded in the physiological limits of dopaminergic firing and the scaling principles of distributional reinforcement learning. At the subcortical level, the amygdala provides a necessary affective penalty to prevent biological or financial ruin. At the cortical level, the ventromedial prefrontal cortex integrates these discrete signals to compute a subjective value, susceptible to top-down cognitive control and context-dependent framing. As advanced computational models isolate true valuation asymmetries from heuristic response biases, the field continues to refine the neurobiological boundaries of Kahneman's foundational principle.