Neural networks and neurochemical modulators of moral judgment
1. Introduction
For centuries, the mechanisms underlying human moral reasoning were the exclusive purview of philosophers, theologians, and legal scholars. However, the advent of modern neuroimaging ushered in an empirical era, fundamentally transforming ethical deliberation into a biological phenomenon that could be measured, localized, and quantified. Early paradigms in the cognitive neuroscience of morality - spurred heavily by landmark studies in the late 1990s and early 2000s - sought to identify the specific neural substrates responsible for ethical decision-making 12. These foundational inquiries often relied on highly artificial ethical dilemmas, such as the infamous "trolley problem," to construct models of moral judgment grounded in a strict dichotomy between automatic emotional intuition and effortful rational deliberation 134.
While these pioneering studies successfully established that moral cognition is a biologically rooted, brain-based ability, they inadvertently propagated theoretical and methodological bottlenecks 2. Early functional magnetic resonance imaging (fMRI) interpretations occasionally leaned toward neo-phrenology, embarking on a misguided search for a localized "moral center" or "moral module" in the brain 25. Furthermore, the overreliance on static, text-based dilemmas administered to highly homogeneous, Western demographics resulted in models of human morality that were historically, culturally, and ecologically constrained 667.
This comprehensive report examines the contemporary landscape of moral neuroscience, focusing specifically on paradigm-shifting developments from 2020 onward. The analysis explicitly dismantles the misconception of a singular moral center, reframing morality instead as an emergent property of distributed, domain-general cognitive and emotional networks 258. By exploring recent advancements in dynamic network-level analyses - particularly the complex spatiotemporal interplay between the Default Mode Network (DMN), the Salience Network (SN), and the Executive Control Network (ECN) - this report details how the brain actively routes moral information 91011.
Furthermore, the report critically evaluates the methodological limitations of early fMRI studies, addressing the replication crisis, the unreliability of analytical workflows, and the logical fallacy of reverse inference 121413. It broadens the investigation of override mechanisms to encompass top-down executive cognitive control, situational context, and the profound influence of neurochemical modulators such as serotonin and oxytocin 141518. Finally, this analysis champions the "ecological turn" in moral neuroscience, highlighting the use of Virtual Reality (VR) to bridge the gap between moral judgment and moral action 1617, and rigorously addresses the imperative to decolonize the field by integrating cross-cultural datasets that overcome the historical WEIRD (Western, Educated, Industrialized, Rich, and Democratic) sampling bias 618.
2. Deconstructing the Myth of the Moral Center: An Emergent Property
A persistent misconception in popular science, and a trap that early neuroimaging studies occasionally fell into, is the assumption that the human brain houses a dedicated "moral module" - a specific neural substrate or exclusive network uniquely evolved to process ethical dilemmas 2519. This localizationist view was historically reinforced by famous lesion studies, most notably that of Phineas Gage, and subsequent investigations into patients with damage to the ventromedial prefrontal cortex (vmPFC), who exhibited profound deficits in real-life moral decision-making and social behavior 24.
However, vast accumulations of neuroimaging, lesion data, and large-scale structural analyses have conclusively demonstrated that no such dedicated moral center exists 520. Recent attempts to isolate a dedicated moral anatomy have proven fruitless. For example, a 2024 anatomical likelihood estimation (ALE) meta-analysis examined the theory of "morality as cooperation," which posits seven specific moral domains: family values, group loyalty, reciprocity, heroism, deference, fairness, and property rights 2125. When researchers investigated the relationship between these seven types of morality and grey matter volume in a massive neuroimaging sample of over 600 individuals, no associations between moral values and grey matter volume survived whole-brain exploratory testing 2125. The researchers concluded that whatever combinations of mechanisms are responsible for morality, they are fundamentally not neuroanatomically localized in any dedicated structural capacity 2125.
Instead, current neuroscientific consensus frames moral cognition as an emergent property resulting from the dynamic integration of multiple, broadly distributed, and domain-general neural networks 2822. When an individual evaluates a moral transgression, they are not activating a specialized "ethics circuit." Rather, they are co-opting fundamental cognitive systems evolved for other survival-oriented purposes: the valuation of rewards and punishments, the simulation of future outcomes, the attribution of mental states to others (Theory of Mind), and the top-down executive inhibition of prepotent impulses 52023.
The finding that moral reasoning relies on overlapping domain-general processes fundamentally shifts the epistemological framework of the field. It implies that "morality" is not a distinct biological category, but a complex, culturally bounded application of general intelligence, social cognition, and affective processing 192328.
2.1 Systematic Mapping of Brain Structures to Moral Functions
To understand how domain-general structures coalesce to produce moral reasoning, it is necessary to map the primary brain regions implicated in ethical decision-making to their general cognitive functions and the theoretical frameworks they historically support.
| Brain Region / Structure | Primary Domain-General Function | Observed Role in Moral Cognition | Competing Theoretical Frameworks |
|---|---|---|---|
| Ventromedial Prefrontal Cortex (vmPFC) | Value representation, emotional regulation, integrating somatic signals. | Generates affective aversion to inflicting direct harm; integrates emotional signals with cost-benefit analysis. | Somatic Marker Hypothesis: Processes intuitive "gut feelings." Dual-Process Theory: Drives the automatic, deontological response 4222425. |
| Dorsolateral Prefrontal Cortex (DLPFC) | Executive control, working memory, cognitive flexibility, rule application. | Overrides prepotent emotional responses; computes abstract, utilitarian outcomes (the "greater good"). | Dual-Process Theory: Drives the slow, rational, consequentialist reasoning system 4153126. |
| Temporoparietal Junction (TPJ) | Theory of Mind (ToM), spatial attention, mentalizing. | Represents the intentions, beliefs, and mental states of agents (e.g., distinguishing intentional vs. accidental harm). | Theory of Mind / Social Brain: Highlights that morality requires evaluating mens rea (guilty mind) 42024. |
| Amygdala | Salience detection, threat processing, fear conditioning. | Generates rapid, bottom-up emotional distress and aversion in response to observing or imagining harm. | Dual-Process Theory: Initiates the "alarm bell" emotional reaction against personal harm 3427. |
| Anterior Cingulate Cortex (ACC) | Conflict monitoring, error detection, reward-based learning. | Detects the cognitive conflict between the emotional aversion to harm and the utilitarian calculation of saving more lives. | Dual-Process Theory: Acts as the conflict mediator when System 1 and System 2 generate competing outputs 82835. |
| Anterior Insula (aINS) | Interoception, empathetic pain processing, disgust. | Encodes the subjective aversiveness of moral transgressions; processes moral disgust and inequity aversion. | Social Neuroscience: Grounds morality in embodied cognition and shared empathetic pain 102329. |
3. Dynamic Network-Level Analyses: Beyond Static Snapshots
Through the 2010s, moral neuroscience was largely dominated by activation studies - identifying which specific regions exhibited increased blood-oxygen-level-dependent (BOLD) signals during a task. However, since 2020, the paradigm has shifted significantly toward network neuroscience and dynamic functional connectivity (dFC) 223038. This approach posits that complex cognitive functions cannot be understood by looking at isolated regions, but only by analyzing the continuous, time-varying interactions, directed information flow, and topological properties of large-scale brain networks 223831.
In the context of moral reasoning, the "Triple-Network Model" has emerged as the dominant framework. This model demonstrates that moral deliberation relies on three intrinsic connectivity networks operating in a highly coordinated, often hierarchical manner: the Default Mode Network (DMN), the Salience Network (SN), and the Executive Control Network (ECN) 91040.
3.1 The Default Mode Network (DMN): Moral Simulation and Self-Reference
The DMN - comprising the medial prefrontal cortex, posterior cingulate cortex, and inferior parietal lobule - was originally identified as the network active during resting states and suppressed during cognitively demanding, externally focused tasks 1132. However, in moral neuroscience, the DMN is heavily recruited when individuals deliberate over "personal" moral dilemmas 910.
Because the DMN governs self-referential mental activity, autobiographical memory, and the simulation of distal events, its activation during moral judgment represents the individual mentally simulating the proposed action and its consequences 101127. When subjects are asked to imagine a complex social transgression, the DMN facilitates the imaginative construction of that scenario, allowing the individual to project themselves into the moral context and compute the associated emotional and social outcomes 427. The profound overlap between regions driving moral cognition and regions supporting thinking about the self highlights that moral evaluations are rarely detached objective calculations; they are deeply anchored in self-projection 24.
Recent translational models have verified the neurophysiological basis of these human fMRI findings. Using fiber photometry combined with fMRI and computational modeling in rodent brains, researchers demonstrated that neuronal activity changes in the DMN are highly synchronized and subject to cyclical transitions, confirming the deep evolutionary roots of the network structures required for internal simulation and valuation 32.
3.2 The Salience Network (SN): The Causal Switchboard
The Salience Network, anchored by the anterior insula (AI) and dorsal anterior cingulate cortex (dACC), operates as a critical detection system that filters and identifies behaviorally relevant internal and external stimuli 91032. Recent dynamic causal modeling and Granger causality analyses have revealed that the SN acts as a modulatory hub during moral reasoning 933.
When a moral transgression or dilemma is detected, the SN assigns emotional and biological salience to the event. Crucially, the SN then causally dictates the engagement or disengagement of the other networks. It acts as a "switch," down-regulating the internally focused DMN and up-regulating the externally focused, rule-applying Executive Control Network (ECN) when complex moral problem-solving is required 101132. Animal models utilizing optogenetic stimulation have confirmed this causal directionality, showing that activation of the anterior insula actively inhibits the retrosplenial cortex, a prominent node of the DMN 32.
Evidence for this hierarchical relationship in humans is drawn from clinical observations: patients with behavioral variant frontotemporal dementia (bvFTD), which primarily causes early atrophy in the SN, demonstrate impaired recruitment of the DMN 91033. Without the SN accurately flagging a situation as morally salient and modulating the DMN, these patients subsequently provide abnormally utilitarian and socially inappropriate responses to moral dilemmas, lacking the internal simulation of harm 91033.
3.3 Dynamic State Transitions in Moral Valuation
Recent advancements in time-series causal analysis and dynamic effective connectivity (dECN) suggest that the brain rapidly transitions through different states of network synchronization during ethical deliberation 3038. Moral cognition is not a single, protracted computational event, but a cyclical transition. Initially, the SN evaluates the emotional salience and potential threat of the scenario; the DMN is then engaged to simulate the social consequences and intent; finally, if a normative rule must be applied or a prepotent emotion overridden, the SN facilitates a transition to ECN-dominant processing 1134.
The balance of these networks is profoundly sensitive. For instance, studies on long-term mindfulness meditation (LTM) reveal that extensive mental training leads to increased cognitive-sensory integration by decoupling affective processes 35. Neuroimaging shows reduced connectivity between the executive network and the salience network in LTMs during pain processing, indicating an ability to experience physical salience without the typical cognitive-affective appraisal, resulting in enhanced emotional neutrality 35.
Conversely, alterations in these micro-transitions - such as hyperconnectivity in the DMN or reduced coupling between the SN and ECN - are heavily implicated in psychopathology. Artificial intelligence models, particularly deep neural networks (DNNs), analyzing fMRI data have successfully predicted the onset of major depressive disorder (MDD) with up to 89% accuracy purely by detecting hyperconnectivity patterns within the DMN and SN that precede clinical symptoms 36. Similar triple-network disruptions characterize narcissistic and antisocial personality traits, where altered resting-state functional connectivity between the DMN and cognitive networks disrupts the typical flow of empathetic moral processing 37.
4. Override Mechanisms: Top-Down Control, Context, and Flexibility
A central pillar of the traditional dual-process theory of moral judgment is the concept of the "override" - the ability of the rational, deliberative system to suppress the immediate, affective response generated by observing harm 327. Early models conceptualized this simply as "reason conquering emotion," but contemporary research reveals a vastly more complex mechanism involving value-based evidence accumulation, neuro-stimulation, and situational flexibility 253147.
4.1 Re-evaluating the Dorsolateral Prefrontal Cortex (DLPFC)
The DLPFC has long been viewed as the primary engine of top-down executive cognitive control. In classic utilitarian dilemmas, deciding to inflict harm for a greater good (e.g., pulling a switch to kill one person but save five) requires overcoming a massive, aversive emotional signal. Neuroimaging correlates these high-conflict utilitarian decisions with robust DLPFC activation 4815.
However, modern causal studies using non-invasive brain stimulation - such as transcranial magnetic stimulation (TMS) and transcranial direct current stimulation (tDCS) - have complicated this picture 1526. While the prevailing hypothesis suggested that enhancing DLPFC activity should universally increase utilitarian, "cold" reasoning by boosting cognitive control, actual findings are highly lateralized and context-dependent. Studies indicate that while the left DLPFC predominantly subserves executive rule-based control, the right DLPFC is heavily involved in affective-cognitive integration 48. Transiently disrupting the right DLPFC via rTMS actually increases the probability of utilitarian responses in objective evaluations of high-conflict dilemmas 26. This suggests that the right DLPFC is not merely a suppressor of emotion, but actively integrates contextual emotional appraisals that are critical for normative moral selection. By knocking it offline, the brain defaults to colder calculus 26.
Furthermore, computational modeling via attribute-based neural drift-diffusion models suggests that DLPFC response during moral choice depends more on value-based evidence accumulation than the mere inhibition of "baser instincts" 31. The DLPFC calculates the cumulative value of a decision over time, modulating its activity based on the subjective weight of the overarching goal. Rather than simply acting as an emotional brake, it actively constructs the subjective value of the normative choice 31.
4.2 Metacognitive Sensitivity and Ideological Neural Signatures
Moral reasoning is profoundly flexible and intimately tied to an individual's ideological framework and metacognitive abilities. The Moral Foundations Theory (MFT) posits that human morality relies on several distinct intuitions: care/harm, fairness/cheating, loyalty/betrayal, authority/subversion, and sanctity/degradation 4938. For years, it was debated whether these foundations were processed uniformly. However, a landmark 2023 study published in Nature Human Behaviour utilized machine-learning algorithms to decode moral judgments directly from brain activation patterns. The researchers found that different moral foundations elicit highly distinct neural signatures, proving that morality is not processed in a single "moral hotspot" but via multiple, distinct pathways 4938.
This study also revealed striking neurological differences driven by political ideology. Liberals exhibited more pronounced neural responses to moral transgressions related to care and fairness (individualizing foundations), whereas conservatives demonstrated elevated Late Positive Potentials (LPPs) and unique neural processing when evaluating binding foundations like loyalty and authority 493851.
The speed and rigidity of these moral judgments are heavily influenced by metacognitive sensitivity - an individual's self-awareness of their own decision accuracy. Research demonstrates that individuals with strong moral convictions process moral decisions more rapidly, showing heightened activation in the SN (anterior insula, dACC) 5239. Crucially, individuals with low metacognitive sensitivity exhibit even stronger neural responses to moralized issues, relying heavily on fast, emotional signals 5254. This neural rigidity, wherein the valuation process is overwhelmed by unquestioned moral conviction, sheds light on the biological underpinnings of political polarization, dogmatism, and the overriding of natural harm aversion to justify ideological violence 523954.
4.3 Situational Context and Acute Stress
The ability to flexibly adapt moral strategies based on situational context is a hallmark of healthy cognition. Neuroimaging studies utilizing varied behavioral tasks (e.g., Cost-Benefit Analysis versus Mitigating Inevitable Misconducts) reveal that neurotypical individuals seamlessly adjust their moral frameworks depending on whether a situation demands strict utilitarian calculation or empathetic mitigation 47. In contrast, individuals with Autism Spectrum Disorder (ASD) demonstrate significantly lower behavioral flexibility during cost-benefit analyses, correlated with reduced activation in the left inferior frontal gyrus 47.
Furthermore, under conditions of acute stress, the influx of neuroendocrine modulators (such as cortisol and adrenaline) directly impairs the functional connectivity between the prefrontal control networks (DLPFC, VMPFC) and the amygdala. This chemical inundation shifts the brain from slow, ethical reasoning to base, instinctual reactions favoring self-preservation, a critical finding for understanding ethical failures in high-pressure professions 40.
5. The Neurochemical Architecture of Moral Choice
While functional networks provide the macro-architecture for moral reasoning, these networks operate within a highly sensitive neurochemical bath. Recent psychopharmacological fMRI studies have demonstrated that altering specific neurotransmitter systems can reliably and predictably shift moral judgments, proving that ethical values are highly susceptible to molecular modulation 5641.

5.1 Serotonin: Harm Aversion and the Economics of Fairness
Serotonin (5-HT) is traditionally associated with mood stabilization, but it plays a profound role in delimiting antisocial behaviors and shaping moral values related to harm and fairness 4142. In experimental settings, boosting serotonin function via citalopram (a selective serotonin reuptake inhibitor, SSRI) drastically alters responses in highly emotionally salient personal moral dilemmas. Subjects become significantly less likely to endorse harming one person to save many others (the utilitarian choice), demonstrating that serotonin directly enhances harm aversion 144243.
This molecular effect translates clearly into social economic interactions, such as the Ultimatum Game. Normally, individuals will reject highly unfair financial offers to punish the proposer, even at a cost to themselves - an act of "moral punishment." However, enhancing serotonin makes subjects less likely to reject unfair offers 43. This counterintuitive finding indicates that serotonin amplifies the aversiveness of harming others; because punishing the proposer inflicts financial harm, the serotonin-enhanced subject avoids the punitive action 5642.
Furthermore, these effects are highly state-by-trait dependent; serotonin's amplification of harm aversion is significantly stronger in individuals who possess high baseline trait empathy 1443. Neurobiologically, serotonin depletion blunts responses in the ventral striatum and medial PFC during fair exchanges, confirming its role in encoding the value of social fairness 41. Groundbreaking research involving awake brain surgery in Parkinson's disease patients has allowed scientists to directly measure these neurotransmitters in real time, confirming that sub-second fluctuations in dopamine and serotonin uniquely drive the emotional and cognitive processes underlying our sense of social justice 60.
5.2 Oxytocin: Moral Emotions and Network Flexibility
Oxytocin (OT) is widely recognized as a prosocial neuropeptide facilitating attachment, trust, and in-group cooperation 184244. However, its role in moral cognition extends far beyond generalized prosociality. Recent studies demonstrate that intranasal oxytocin specifically amplifies "moral emotions" such as guilt and shame, and decreases the willingness to cause deliberate harm for personal benefit 1862. Interestingly, this effect is distinct from other social hormones like vasopressin, indicating that the neurochemical pathways for general prosocial behavior and specific moral judgments are dissociable 1862.
At the network level, oxytocin exerts extensive modulatory effects on dynamic brain connectivity 6364. Co-activation pattern (CAP) analysis of resting-state fMRI data reveals that administration of oxytocin alters the topology of the DMN and frontoparietal networks, increasing the frequency of state transitions, in-degree, and out-degree, thereby enhancing overall network flexibility 6465. It specifically targets top-down control over emotional processing hubs, such as the amygdala, altering its connectivity with the salience network and the medial prefrontal cortex 6364.
This increased dynamic flexibility enables the brain to rapidly switch states to adapt to complex social and emotional contexts. Notably, age-stratified analyses reveal that while young adults exhibit more pronounced modulation of dynamic flexibility, older adults show more sustained engagement with emotion-related states post-oxytocin administration 64. In clinical contexts, such as Autism Spectrum Disorder (ASD), oxytocin has been shown to target widespread enhancement of local neural network processing, modulating local integration within brain regions involved in emotional regulation, positioning it as a powerful therapeutic tool 45.
6. Methodological Critiques: Reverse Inference and the Replication Crisis
Despite the rapid advancement of moral neuroscience, the field has been subject to intense methodological scrutiny. The transition from the foundational studies of the early 2000s to the rigorous data science standards of the 2020s has exposed several critical vulnerabilities in how moral neuroimaging is conducted and interpreted 121446.
6.1 The Fallacy of Reverse Inference
The most pervasive epistemological flaw in cognitive neuroscience, heavily scrutinized in recent literature, is the problem of "reverse inference" - a logical fallacy highlighted prominently by researchers like Russell Poldrack 146847. In standard forward inference, a researcher induces a cognitive process (e.g., moral deliberation) and observes which brain regions activate. Reverse inference occurs when a researcher observes activation in a specific region (e.g., the amygdala) and deduces that a specific cognitive state (e.g., fear or emotional distress) must be occurring 1447.
This reasoning is an instance of affirming the consequent, functioning as a non-deductive abductive logic. It is logically invalid because brain regions are invariably multi-functional 144748. The anterior cingulate cortex, for example, is involved in conflict monitoring during moral dilemmas, but it is also engaged during physical pain, task switching, and reward anticipation 82848. Entire theories of moral cognition have historically been built on reverse inferences, assuming that because the vmPFC lit up, the subject was primarily experiencing a "gut emotion." Modern neuroinformatics, utilizing databases like NeuroSynth and BrainMap, are actively seeking to quantify and mitigate reverse inference by calculating precise positive predictive values for specific neural-cognitive mappings, shifting the field toward formal ontologies 4748.
6.2 The Replication Crisis and Analytical Flexibility
Moral neuroscience has not been immune to the broader replication crisis affecting psychology and biomedicine 124649. Many foundational fMRI studies on moral dilemmas relied on critically small sample sizes (often fewer than 20 participants), which drastically reduces statistical power and inflates the likelihood of false positives 4649.
Furthermore, the sheer complexity of fMRI data analysis introduces massive "analytical flexibility." The raw blood-oxygen-level-dependent (BOLD) signal requires extensive preprocessing, spatial smoothing, and statistical thresholding 1213. A landmark meta-analytical study recently highlighted this vulnerability by distributing the exact same raw fMRI dataset to 70 independent research teams, asking them to test nine identical hypotheses. Because no two teams used the exact same analytical workflow, they routinely produced widely divergent results 1213.
Additionally, longitudinal studies have questioned the reliability of fMRI in teasing out individual differences during emotion-processing tasks, demonstrating that brain activity patterns often do not hold up when people are scanned multiple times months apart 13. Combined with historical embarrassments - such as the infamous study finding fMRI signals in a dead salmon due to failure to correct for multiple comparisons, or the realization that simple breathing patterns can distort scan results - these methodological revelations demand modern preregistration, robust sample sizes, and algorithmic transparency in current neuro-moral investigations 134649.
6.3 Critiques of Early Dual-Process Paradigms
At the behavioral level, the reliance on extreme, artificial dilemmas - like the trolley problem - has drawn sharp criticism 350. Critics argue that categorizing responses purely as "utilitarian" versus "deontological" oversimplifies human ethics. Furthermore, early paradigms by Greene and colleagues asked subjects if an action was "appropriate" or "inappropriate," words that are inherently ambiguous and conflate moral permissibility, legal requirement, and social convention 350. The highly contrived nature of these dilemmas - often involving bizarre scenarios that participants find humorous or absurd rather than morally gripping - severely limits the generalizability of the findings to everyday moral transgressions, failing the test of ecological validity 565051.
7. The Ecological Turn: Virtual Reality and Naturalistic Paradigms
Recognizing the limitations of static, text-based dilemmas, the neuroscience of morality is undergoing an "ecological turn" 165152. To accurately map how the brain computes moral decisions in the real world, researchers require stimuli that evoke genuine emotional, physiological, and cognitive engagement.
Over the past decade, Virtual Reality (VR) has emerged as a revolutionary tool in moral psychology and neuroscience, particularly when integrated with fMRI 1653. Immersive VR simulations transport participants into ethically challenging scenarios - such as driving an autonomous vehicle facing an unavoidable collision, or navigating a triage situation - that would be impossible or highly unethical to recreate in physical reality 16.
Crucially, VR research highlights a profound gap between moral judgment (what we say we would do) and moral action (what we actually do) 1652. When reading a text-based trolley dilemma, participants engage heavily in semantic processing and abstract mental simulation (reliant on the DMN). However, when placed in a fully immersive VR trolley scenario - complete with spatial realism, auditory cues (screaming), and the physical requirement to execute a motor action - participants exhibit drastically different behavioral and neural profiles. VR studies consistently reveal a general trend toward utilitarian choices compared to text-based paradigms. This shift is heavily modulated by the visceral, emotional salience of the environment, the perceived age of victims, and extreme time pressure 1617.
By incorporating multimodal data such as eye-tracking, galvanic skin response, and real-time hemodynamic tracking, VR enables researchers to parse the temporal dynamics of moral choice 1617. It shifts the experimental focus from cold, cognitive evaluations of hypothetical rules toward the hot, embodied, and highly contextual nature of true moral decision-making, allowing researchers to simulate scenarios directly relevant to modern challenges, such as programming ethics into autonomous systems 1651.
8. Decolonizing Moral Neuroscience: Overcoming the WEIRD Bias
Perhaps the most significant limitation in the history of moral neuroscience is its reliance on WEIRD populations - Western, Educated, Industrialized, Rich, and Democratic 66. Approximately 96% of subjects in top behavioral science and psychology journals are drawn from WEIRD demographics, even though these populations constitute only 12% of the global population 654.
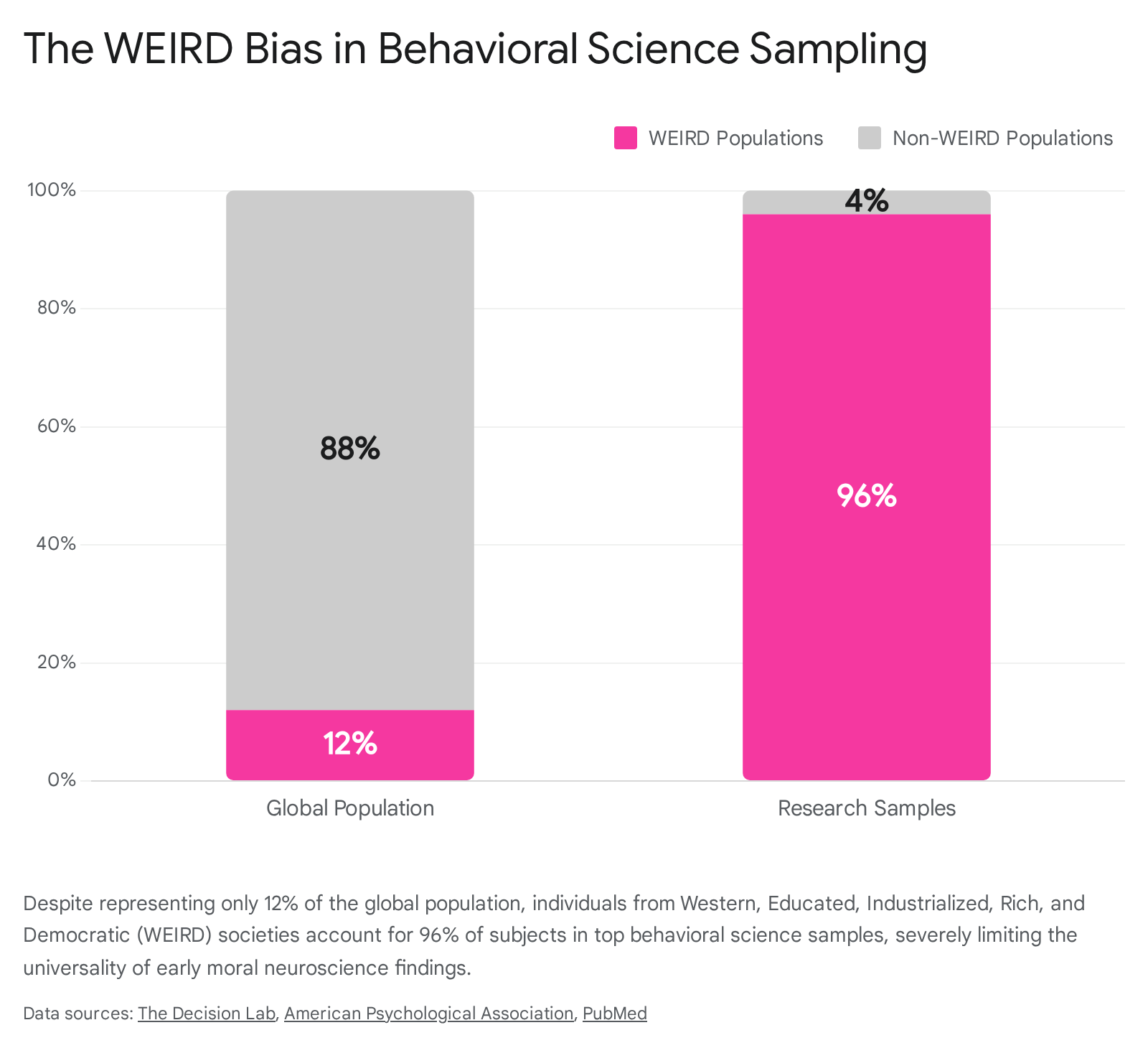
Historically, researchers implicitly assumed that basic cognitive and moral processes were universal, treating the WEIRD brain as the standard human prototype 66.
8.1 Cross-Cultural Variability in Moral Cognition
As neuroscientists and anthropologists peek beyond the WEIRD bubble, they find that moral cognition is profoundly shaped by cultural norms, values, and ecological variables 654. For instance, Western moral and legal systems place a paramount emphasis on mens rea - the intentionality and mental state of the actor. In WEIRD populations, accidental harm is judged far less severely than intentional harm, reflecting a high degree of "mind-mindedness" 7.
However, this is not universally dominant. Cross-cultural research reveals substantial variability. Studies involving Indigenous iTaukei Fijians and other non-Western societies demonstrate a "mental opacity" norm, where moral judgments are highly outcome-oriented 7. In some communities, a person who causes harm by an honest mistake is judged nearly as harshly as someone who harms intentionally 7. While some elements of moral reasoning - such as the basic aversion to direct harm - appear relatively conserved across diverse populations (e.g., Mayan communities exhibiting similar responses to certain trolley variations), the weighting of intentions, purity, in-group loyalty, and social hierarchy varies vastly 1.
Recent neuroimaging studies explicitly exploring East Asian populations have revealed divergent neural patterns. Using functional near-infrared spectroscopy (fNIRS) alongside the Consequences, Norms, Inaction (CNI) model, researchers found significant cross-cultural and gender-based differences in prefrontal cortex (PFC) activity during moral judgments between Japanese and Chinese individuals 77. These findings confirm that cultural emphasis on interdependence and collective responsibility structurally influences how the prefrontal cortex processes and resolves moral dilemmas 7755.
8.2 Structural Inequities in Global Neuroscience
Addressing the WEIRD bias requires more than simply translating surveys into new languages; it demands addressing structural and technological inequities 1856. As the Middle East and Africa begin to establish state-of-the-art neuroimaging facilities - such as the establishment of multi-nuclear MRI facilities in Saudi Arabia - researchers are uncovering unique epidemiological and cognitive data specific to these populations 8057. For example, analyses utilizing the National Health Interview Survey (NHIS) reveal that the prevalence of cognitive limitations among foreign-born Arab Americans and broader Middle Eastern/North African (MENA) populations often diverge significantly from US-born non-Hispanic Whites, indicating that the cognitive health trajectories of non-Western groups require localized, specific study 57.
However, neuroethics in the Global South faces unique challenges. Technologies developed in the West often fail to account for phenotypic diversity - for example, standard EEG caps frequently fail to accommodate Afro-textured hair. This seemingly minor technical oversight acts as a form of "techno-chauvinism," systemically excluding specific populations from foundational neurophysiological research 18.
Furthermore, African bioethicists warn against "ethics dumping" - the practice wherein Western researchers export ethically questionable or high-risk neuroscientific research to African countries with less stringent regulatory oversight 1856. A truly universal cognitive neuroscience of morality requires empowering local researchers to develop culturally grounded neuro-narratives. For instance, recent theoretical reviews have analyzed the works of Nigerian author Amos Tutuola as an "indigenous cognitive archive," demonstrating how Yoruba folklore intuitively encodes predictive reward models, fear learning, and moral conditioning that align perfectly with modern behavioural neuroscience, yet operate outside Western hyper-individualist models 82. Respecting local philosophical frameworks, such as Ubuntu - which emphasizes relational identity and interconnectedness - is crucial for generating ecologically valid neuro-moral paradigms 5682.
9. Conclusion and Future Directions
The cognitive neuroscience of morality has matured remarkably over the past two decades. The field has moved decisively past the neo-phrenological search for a localized "moral module," recognizing instead that human morality is an emergent, highly complex property born from the dynamic interaction of domain-general brain networks 2522. By mapping the continuous interplay between the Default Mode Network, the Salience Network, and the Executive Control Network, researchers now understand moral deliberation not as a static event, but as a fluid, time-varying integration of internal simulation, threat detection, and cognitive control 9113038.
Simultaneously, the recognition of powerful override mechanisms - driven by value accumulation in the prefrontal cortex and modulated intricately by neurochemicals like serotonin and oxytocin - illustrates that moral values are biologically flexible and acutely sensitive to context, stress, and pharmacological intervention 31404264. Advanced machine-learning tools, such as deep neural networks and novel algorithms like the "Krakencoder," are now allowing researchers to map the relationships between the structural connectome (the brain's physical wiring) and the functional connectome (activity patterns), bringing us closer to understanding exactly how anatomy gives rise to ethical behaviors 5859.
Looking forward, the future of moral neuroscience relies heavily on an ongoing methodological and ecological evolution. The field must definitively shed its reliance on artificial, text-based dilemmas that fail to capture the visceral reality of ethical conflict, leveraging Virtual Reality to probe the urgent gap between moral reasoning and moral action 1652. Furthermore, resolving the replication crisis demands rigorous statistical power, algorithmic transparency, and an absolute rejection of unfounded reverse inferences 4647. Most importantly, cognitive neuroscience must actively work to decolonize its datasets. By recognizing the profound impact of cultural frameworks on neural processing and investing in diverse global research infrastructure, the field can finally construct a truly comprehensive, species-wide understanding of the moral brain 61855.