Neural mechanisms and individual differences in color perception
The Physical Stimulus and Neurological Construct
In the disciplines of optics and physics, color does not inherently exist as an objective property of the physical environment. Objects absorb specific segments of the electromagnetic spectrum and reflect others, projecting a varying distribution of wavelengths toward the observer 123. The perception of color is entirely a neurological construct - an intricate illusion engineered by the visual cortex to distinguish surfaces, identify objects, and navigate complex spatial environments 12. The fundamental dissociation between objective physical wavelengths and subjective perceptual experiences forms the basis of contemporary visual neuroscience 3.
When light strikes a surface, such as a green leaf, the material absorbs various wavelengths while reflecting radiation concentrated in specific nanometer ranges back to the eye 3. The leaf possesses no intrinsic color; it only possesses intrinsic reflectance properties governed by its molecular composition 13. The cognitive experience of "green" arises only after the reflected wavelengths stimulate specialized photoreceptor cells in the retina, initiating a cascade of electrochemical signals that ascend through the visual hierarchy 13. The human capability to perceive roughly one million distinct colors is an evolutionary adaptation; cognitive scientists propose that trichromatic color vision developed in primates primarily to facilitate complex social communication and the identification of ripe foliage and fruit against forest backgrounds 2.
Because color is synthesized centrally within the brain rather than measured objectively at the sensory periphery, perceptual experience is highly malleable. Contextual information, spatial gradients, ambient illumination, and prior visual experiences actively shape how the brain assigns hue to a given surface 1. This top-down influence produces phenomena where identical wavelengths of light can be perceived as drastically different colors depending on their surrounding context, demonstrating that the visual system prioritizes relational consistency over absolute photometric accuracy 2. The brain relies on hierarchical stages of processing, extracting basic features like edges and local contrast in early visual areas before deriving complex surface color representations in higher-order regions 1.
Foundational Theories of Color Vision
The historical pursuit to understand how human vision decodes spectral information yielded two dominant theoretical frameworks in the nineteenth century. For decades, these theories were viewed as mutually exclusive paradigms. Modern neurobiology has since reconciled them, demonstrating that both frameworks are fundamentally correct but operate at different neuroanatomical stages of the visual processing hierarchy 47.
Trichromatic Theory
The Trichromatic Theory, originally proposed by Thomas Young in 1802 and later expanded by Hermann von Helmholtz in the 1860s, posits that human color vision is mediated by three distinct types of photoreceptors 478. Operating at the cellular level of the retina, this mechanism relies on cone cells that exhibit peak sensitivities to different segments of the visible light spectrum 9.
The human retina contains approximately six million cones, categorized into three classes based on the specific photopigments they contain 78. According to the trichromatic framework, any perceived hue is the result of the combined, proportional activation of these three cone types 49. For instance, perceiving yellow requires the simultaneous stimulation of M-cones and L-cones 89. While this theory accurately models the initial capture of light, it fails to explain specific psychophysical phenomena, such as why humans cannot perceive a "reddish-green" or a "bluish-yellow," and why staring at a red object produces a green afterimage 785.
| Cone Designation | Wavelength Sensitivity | Peak Perception | Retinal Distribution |
|---|---|---|---|
| S-Cones | Short-wavelength | Blue | ~10% of cone population |
| M-Cones | Medium-wavelength | Green | ~30% of cone population |
| L-Cones | Long-wavelength | Red | ~60% of cone population |
Opponent-Process Theory
To address the explanatory gaps of the Trichromatic Theory, German physiologist Ewald Hering proposed the Opponent-Process Theory in the late 1800s 89. Hering hypothesized that the visual system processes color not through independent receptors, but through antagonistic neural channels 79.
This theory outlines three opponent pairs: red versus green, blue versus yellow, and a black versus white achromatic luminance channel 478. In this framework, the neurological pathways for opposing colors cannot transmit signals simultaneously. Activation by red light inhibits the pathway for green, rendering the perception of "reddish-green" a biological impossibility 789. Furthermore, the opponent-process model elegantly explains negative afterimages: prolonged exposure to a red stimulus causes neural fatigue in the red-sensitive channel; when the observer subsequently shifts their gaze to a neutral white surface, the baseline firing rate of the fatigued red channel drops below its resting state. This reduction in inhibition allows the opponent green channel to dominate the perceptual output temporarily, producing a green afterimage 95.
Theoretical Synthesis and Zonal Equivalence
Contemporary visual neuroscience synthesizes these two paradigms into a cohesive hierarchical model. The physicist Erwin Schrödinger contributed to the mathematical unification of these concepts by demonstrating the common geometry of the chromaticity diagrams derived from both theories, proving they were functionally equivalent stages rather than competing facts 6.
The physiological reality is a "zone theory" of vision. The Trichromatic Theory accurately describes the earliest stage of visual processing: the absorption of light by S, M, and L cones at the retinal surface 475. As these signals propagate deeper into the retina toward bipolar and retinal ganglion cells, the neural circuitry reorganizes the raw trichromatic inputs into opponent channels 475. For example, a red-green opponent ganglion cell may receive excitatory input from L-cones and inhibitory input from M-cones. This mathematical transformation of signals from a three-receptor additive code to an opponent-channel neural code bridges the gap between wavelength detection and higher-order color perception 476.
Anatomical Pathways of Visual Processing
The transformation of electromagnetic radiation into a coherent, colorful representation of the world requires an extensive network of specialized anatomical structures. Visual processing is not restricted to the cerebral cortex; substantial computation occurs locally within the eye before information is transmitted to the brain 7.
Retinal Extraction to the Lateral Geniculate Nucleus
The visual pathway begins at the retina, a complex ten-layered structure of neural tissue 8. Rods, optimized for low-light and achromatic vision, populate the periphery of the retina, while cones, optimized for high-acuity color vision, are concentrated primarily in the central fovea 78915. Cones generate action potentials through photosensitive cycles and synapse onto bipolar cells, which in turn synapse onto retinal ganglion cells (RGCs) 78. Within this retinal microcircuitry, horizontal and amacrine cells provide lateral inhibition, establishing the initial basis for brightness discrimination, motion detection, and spatial color contrast 7.
The unmyelinated axons of the RGCs converge at the optic disc to form the optic nerve, transmitting signals from more than one million nerve fibers 789. Upon exiting the orbit, the optic nerves from both eyes converge at the optic chiasm. At this anatomical intersection, a highly specific decussation occurs: fibers originating from the nasal hemiretina of each eye cross over to the opposite side of the brain, while fibers from the temporal hemiretina remain uncrossed 78910. This structural arrangement ensures that information from the left visual field - captured by the right nasal retina and the left temporal retina - crosses at the optic chiasm, routing to the contralateral Lateral Geniculate Nucleus (LGN) before terminating in the primary visual cortex of the right hemisphere, and vice versa 7910.
The reorganized fibers, now termed the optic tract, travel to the thalamus, synapsing primarily at the LGN 781510. The LGN acts as an essential relay and gating center, maintaining the strict separation of parvocellular streams (mediating color and fine detail) and magnocellular streams (mediating motion and luminance) 7. From the LGN, optic radiations carry the visual data to the primary visual cortex (V1) located in the occipital lobe 91510.
Primary Visual Cortex and Early Processing
The primary visual cortex (V1), or striate cortex, is responsible for the initial cortical processing of visual stimuli. The foundational work of neurophysiologists David Hubel and Torsten Wiesel in the late 1950s and 1960s revolutionized the understanding of this region. Through single-unit recordings in the visual cortex of cats and monkeys, they demonstrated that V1 neurons are not simply point-to-point feature detectors for spots of light, but are highly organized structures tuned to specific line orientations, edges, and spatial frequencies 1112131415.
Hubel and Wiesel mapped ocular dominance columns and hierarchical processing sequences - moving from simple cells that respond to oriented slits of light, to complex cells that aggregate inputs from simple cells to detect motion and pattern 12131415. Regarding color processing, V1 contains specialized regions known as 'blobs' containing double-opponent cells that compute basic color contrasts 1617. However, V1 primarily registers the presence and intensity of different wavelengths and local wavelength differences; it does not achieve global color constancy 1819. The data processed in V1 is subsequently sent to V2, where early surface and texture segregation begins, featuring specific hue representations in color bands spanning the thin stripes of V2, before propagating through the ventral stream for object identification 152021.
Cortical Area V4 and Color Constancy
Cortical area V4, located in the mid-tier of the ventral visual pathway (specifically within regions encompassing the fusiform gyrus), is recognized as a crucial hub for intermediate shape processing and higher-order color perception 192122292324. Early neuroimaging and lesion studies associated the V4 complex with the conscious perception of color, color imagery, color ordering tasks, and the assignment of color knowledge to objects 182324.
V4 distinguishes itself from earlier visual areas primarily through the mechanism of color constancy. Color constancy is the visual system's capacity to perceive an object as maintaining a stable hue despite drastic changes in the spectral composition of the ambient illumination 192125. For example, a red apple reflects a vastly different wavelength profile under the warm light of a sunset compared to the cool light of a fluorescent bulb, yet the observer consistently perceives it as red 21. Single-unit recording studies indicate that neurons in V4 adjust their color-tuning functions dynamically in response to shifts in background lighting, aligning neuronal firing rates with the subjective psychophysical percept rather than the objective wavelength inputs 192126.
Research indicates a functional subdivision within the human V4 complex: the posterior section is referred to as V4, and the anterior section as V4α 192127. Functional MRI (fMRI) investigations show a gradient across the visual hierarchy, where early areas like V1 encode the raw physical illumination, while V4 and particularly V4α progressively encode the invariant surface color regardless of illumination 2124. Furthermore, V4 neurons represent more complex spatial geometries. Recent cortical inactivation experiments in non-human primates demonstrate that millimeter-scale domains in V4 are tuned not just to colors, but to polar and hyperbolic gratings (curvature), indicating that V4 operates as an image segmentation and grouping engine where color and shape are tightly integrated 2935.
High-resolution fMRI analyses utilizing representational similarity analysis (RSA) have also mapped functional asymmetries across the V4 area 2228. Ventral parcels of V4 exhibit distinct response profiles to chromatic and luminance contrasts compared to dorsal parcels, showing decreasing dependence on spatial frequency and increasing dependence on pure color contrast as signals ascend the hierarchy from V1 to V4 2228.
Color Vision Deficiencies and Neurological Adaptations
Dysfunctions within the retinal or cortical pathways result in color vision deficiencies, a condition commonly but inaccurately termed "color blindness." A complete inability to perceive color is extraordinarily rare; rather, color vision deficiencies generally manifest as a restricted chromatic gamut and an inability to accurately distinguish between specific hues 29303132.
Genetic Basis and Photoreceptor Dysfunction
Color vision deficiencies are predominantly congenital and genetically linked, frequently resulting from mutations on the X chromosome 2930. Because males possess a single X chromosome, a recessive mutation affecting cone photopigments inevitably results in phenotypic expression. Consequently, anomalous color vision affects approximately 1 in 12 men, compared to only 1 in 200 women 293032. Beyond genetics, acquired deficiencies can develop later in life secondary to neurodegenerative conditions, chemical exposure, or ocular pathologies such as glaucoma and macular degeneration, which disrupt the optic nerve or central retina 293032.
When a specific cone class (S, M, or L) is mutated or absent, the brain receives an incomplete trichromatic signal. Because downstream color perception relies heavily on the opponent-process comparison of cone signals, the loss of one cone type collapses the differentiation along a specific color axis, causing distinct visual confusions 3031.
| Condition Category | Specific Subtype | Retinal Mechanism | Perceptual Outcome |
|---|---|---|---|
| Anomalous Trichromacy | Protanomaly | Mutated L-cones | Reduced sensitivity to red; reds, greens, and oranges appear muddy or brown. |
| Anomalous Trichromacy | Deuteranomaly | Mutated M-cones | Reduced sensitivity to green; the most prevalent form of "red-green" deficiency. |
| Anomalous Trichromacy | Tritanomaly | Mutated S-cones | Reduced sensitivity to blue; confusion between blue/yellow and green/blue. |
| Dichromacy | Protanopia | Complete absence of L-cones | Severe red-green confusion; reds appear dark brown or black. |
| Dichromacy | Deuteranopia | Complete absence of M-cones | Severe red-green confusion; mid-reds and mid-greens are entirely indistinguishable. |
| Dichromacy | Tritanopia | Complete absence of S-cones | Inability to distinguish blue from green, and yellow from pink/red. |
| Monochromacy | Achromatopsia | Absence or dysfunction of all cones | True grayscale vision mediated by rods, often paired with severe photophobia and low acuity. |
Cortical Responses in Anomalous Vision
Recent neuroimaging studies have investigated how the visual cortex adapts to deficient retinal inputs. In functional MRI studies comparing individuals with normal trichromatic vision, Daltonism (red-green deficiencies), and achromatopsia, researchers observed distinct cortical activation patterns 33. In normal trichromats, the hV4 region demonstrates the highest chromatic sensitivity within the ventral visual stream 33.
However, in individuals with Daltonism and achromatopsia, while cortical activation is robust under combined chromatic and luminance conditions, there is no significant color-specific activity detected in hV4 33. This indicates that color-deficient individuals rely heavily on luminance-based compensatory mechanisms to discriminate surfaces 33. The disruption of chromatic pathways forces the visual cortex to adapt, utilizing brightness gradients and textures to maintain functional object recognition despite the absence of a stable hue representation 33.
Visual Ambiguity and Color Constancy Phenomena
The evolutionary advantage of color constancy is profound - it ensures that object recognition remains stable across shifting diurnal and environmental conditions. However, because color constancy relies on the brain making subconscious assumptions about the primary light source (the illuminant), ambiguous visual stimuli can cause the system to generate conflicting perceptual conclusions across different observers 343536.
The Mechanics of Discounting the Illuminant
To maintain a constant perception of surface color, the visual system must subtract the spectral contribution of the illuminating light 3738. When an observer looks at an object outdoors at midday, the brain detects a blue bias from the sky and neurologically "subtracts" blue from the visual field, adding yellow tones to compensate 37. If the same object is viewed indoors under incandescent lighting, the brain detects a yellow bias and subtracts it, adding blue 37.
In natural environments, the visual scene contains ample context - shadows, horizons, and familiar objects - to accurately estimate the illuminant. However, when an image is tightly cropped or artificially filtered, the visual system is forced to estimate the nature of the light source based on internal prior probabilities 3637.
The Cyan Strawberries Illusion
A vivid demonstration of this compensatory mechanism is the "Cyan Strawberries" illusion, created by Japanese psychologist Akiyoshi Kitaoka 3839. The image features a plate of strawberries that clearly appear red. However, digital color analysis reveals that the image contains zero red pixels; the pixels composing the strawberries are entirely cyan (a mixture of blue and green) and gray 383940.
The brain perceives the strawberries as red due to two overlapping mechanisms. First, the entire image is washed in a heavy blue-green filter. Recognizing this pervasive tint as an unnatural illuminant, the color constancy algorithms in V4 automatically discount the cyan light 363840. When cyan is subtracted from gray pixels, the perceptual result is pushed toward the opposite end of the opponent-process spectrum - red 3840. Second, the human brain holds strong prior associations regarding the memory color of familiar objects. Recognizing the geometric shape, dimples, and texture of strawberries, the brain expects them to be red, further accelerating the top-down color-correction process 3840.
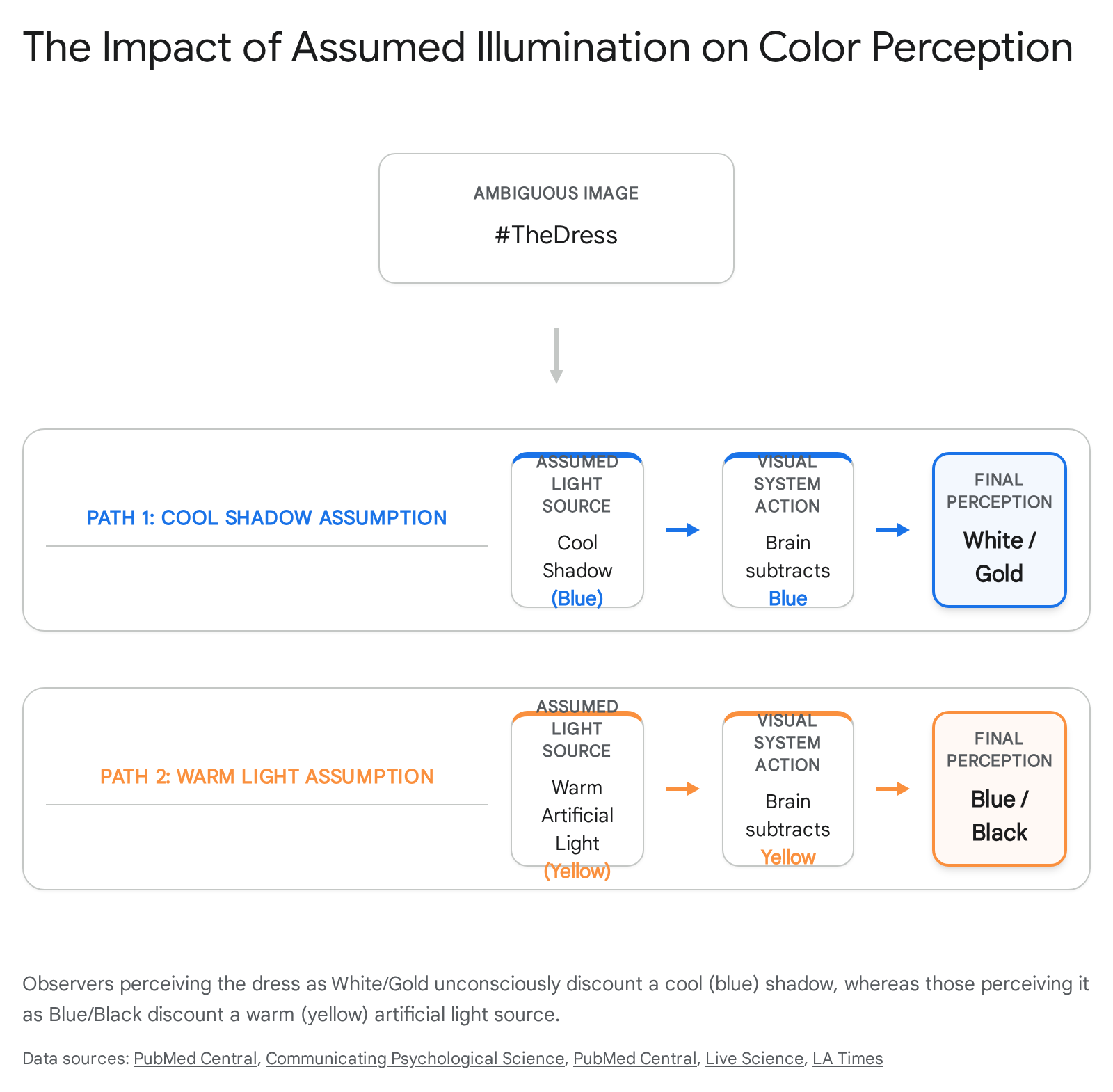
The Dress and The Shoe Case Studies
In February 2015, a photograph of a striped dress (#theDress) triggered a global debate over its color, marking a rare instance of a naturally occurring, deeply bimodal visual illusion 344142. Roughly 57% of observers perceived the garment as blue and black, while 30% perceived it as white and gold, with a small minority seeing blue and brown 4243. A similar phenomenon emerged in 2017 with a photograph of a shoe (#theShoe), which observers split between perceiving as pink/white or teal/gray 34374445.
These images represent extreme challenges in illuminant estimation. Both photographs were poorly lit, overexposed, and lacked broad contextual cues, rendering the illumination source fundamentally uncertain 3746. Consequently, the brain is forced to rely on internal priors to solve the inverse optics problem 3547.
If an observer's visual system unconsciously assumes the dress is located in a cool, blue-lit shadow, it discounts the short-wavelength (blue) light, resulting in a perception of a white and gold dress 34354344. Conversely, if the observer's brain assumes the dress is under warm, artificial incandescent lighting, it discounts the long-wavelength (yellow) light, leaving the true colors of the pixels: blue and black 343543.
Predictors of Perceptual Divergence
Extensive neuro-ophthalmological and psychological research into the #theDress phenomenon reveals that the choice of illuminant prior is not random; it is influenced by an individual's biology, daily environment, and age.
Chronotype and Lifestyle Context: Studies suggest a strong correlation between an individual's circadian rhythm (chronotype) and their perception of ambiguous colors 354748. "Larks" (individuals who wake early and spend a larger proportion of their waking hours exposed to natural blue-sky daylight) are statistically more likely to assume a blue illuminant, automatically discounting it to see the dress as white and gold 354648. In contrast, "owls" (individuals who stay up late and are predominantly immersed in warm, long-wavelength artificial lighting) are more likely to assume a yellow illuminant, discounting it to perceive the dress as blue and black 354648.
Macular Pigment Optical Density (MPOD) and Age: The anatomical composition of the eye also introduces hard-wired variations in signal processing before the visual cortex even receives data. The human macula contains a yellow pigment that naturally absorbs short-wavelength (blue) light 4149. Individuals with a higher Macular Pigment Optical Density (MPOD) filter out more blue light before it reaches the photoreceptors 4950. Because MPOD gradually increases with age and is augmented by the ingestion of carotenoid-rich foods (such as green leafy vegetables), demographic shifts in perception are observable 495152. Studies analyzing cohorts ranging from young children to older adults demonstrate that older individuals, and those with a higher dietary green-leaf preference, are significantly more likely to perceive the dress as white and gold 42434951.
Genetics versus Environment: The extent to which perception is hardwired versus learned was explored in twin studies. By comparing the perceptual similarities between identical and non-identical twins viewing #theDress, researchers estimate that approximately 34% of the variance in color perception is attributable to genetic differences (which likely dictate baseline ocular anatomy such as pupil size and inherent neural architecture) 4150. The remaining 66% is driven by environmental factors, including learning, geographic lighting exposure, and accumulated lifelong visual experiences 41.
| Predictive Factor | Mechanism of Action | Correlated Perception of The Dress |
|---|---|---|
| Chronotype (Larks) | High exposure to natural daylight; visual system defaults to assuming cool (blue) illuminants. | White and Gold |
| Chronotype (Owls) | High exposure to artificial light; visual system defaults to assuming warm (yellow) illuminants. | Blue and Black |
| Increased Age & MPOD | Higher macular pigment density absorbs short-wavelengths, physically filtering blue light. | White and Gold |
| Professional Art Experience | Increased cognitive flexibility in interpreting visual stimuli; higher likelihood of category switching. | Capable of switching perceptions |
Linguistic Relativity and Categorical Perception
Beyond the lower-level mechanics of the retina and the mid-level color constancy computations of the visual cortex, the subjective experience of color is also heavily modulated by higher-order cognitive faculties, specifically language. The Sapir-Whorf hypothesis, or linguistic relativity, posits that the structure and vocabulary of a person's language actively shape how they perceive and categorize reality 5354. In visual neuroscience, this raises a fundamental question: does the lack of a specific color word alter the neurological capacity to discriminate that hue?
The Russian Blue Boundary
The Russian language lacks a single generic term equivalent to the English word "blue." Instead, it enforces an obligatory boundary between light blue (goluboy) and dark blue (siniy) 545556. Researchers investigated whether this deep-seated linguistic categorization provides Russian speakers with a measurable perceptual advantage in color discrimination 5556.
In speeded visual discrimination tasks, Russian speakers were significantly faster than English speakers at distinguishing two shades of blue if those shades crossed the linguistic boundary (one siniy and one goluboy) compared to when the two shades fell within the same category (both siniy or both goluboy) 5657. This speed advantage occurred even when the objective colorimetric distance between the paired shades was mathematically identical 5657. Crucially, this reaction-time advantage was entirely eliminated when the subjects were required to perform a simultaneous verbal interference task (e.g., repeating a string of numbers), but it was not affected by a spatial interference task 5556. This experimental outcome indicates that the language centers of the brain actively intercede in perceptual decision-making; the linguistic label acts as an online cognitive tool that accelerates visual categorization across borders that the language deems significant 545558.
The Himba Tribe Categorization Debate
One of the most widely discussed - and occasionally misrepresented - studies on linguistic relativity involves the Himba people, a semi-nomadic tribe in northern Namibia 535960. The Himba language utilizes five basic color terms, which partition the visual spectrum quite differently than Western languages 5359. Notably, they possess a single word (buru) that encompasses various shades of green and blue, but have multiple distinct terms differentiating what English speakers would classify simply as different shades of green (dambu vs. vapa) 5960.
Initial cross-cultural experiments by Debi Roberson and Jules Davidoff utilizing an odd-one-out paradigm demonstrated that Himba participants were remarkably fast at identifying a subtly different shade of green among a circular array of otherwise identical green tiles - a task English speakers found highly difficult due to lacking a specific vocabulary to differentiate the greens 596069. Conversely, when presented with a circle of green tiles and one clearly blue tile, Himba participants took a longer time to identify the blue oddball than their English-speaking counterparts 596069.
However, public reporting of these findings occasionally evolved into hyperbole. Certain media representations, including a widely circulated BBC documentary segment, implied that the Himba were biologically or perceptually "blind" to the color blue, fabricating a dramatized experimental sequence where tribe members seemingly failed to see a blue square entirely 6961. The original researchers subsequently refuted this framing, clarifying that the Himba never failed to see the blue tile; the dependent variable in the rigorous scientific studies was solely reaction time and memory retention, not absolute visual capacity or failure to detect the stimulus 6162. The Himba visual cortex processes blue and green wavelengths identically to any neurotypical human; however, the lack of a distinct linguistic boundary slows the cognitive categorization required to rapidly complete the experimental sorting task 596162. This underscores the precise nuance of linguistic relativity: language does not alter lower-level photometric absorption in the retina, but it profoundly sculpts higher-level perceptual categorization and visual memory 5362.
Computational Models of Visual Cortex and Color Processing
The intricate hierarchy of color processing - from the cone-opponent signals in the retina to the complex shape-color integration in cortical area V4 - has increasingly become a focal point for researchers developing artificial intelligence 161935. Biologically inspired computational models are now being leveraged to solve complex machine vision challenges, while simultaneously offering a computational sandbox to test and validate hypotheses about the human brain.
Biologically Inspired Architectures
Historically, computer vision systems struggled immensely with color constancy, easily failing when ambient lighting shifted in a scene 166364. To overcome this, researchers have developed Spiking Neural Networks (SNNs) and Hierarchical Convolutional Energy (HCE) models mapped directly to the known anatomy of the primate visual system 3565.
In models such as Bio-CC (Biologically inspired Color Constancy), algorithms are structured to mimic the exact physiological pathway from the retina to the LGN, V1, and V4 1666. These models incorporate simulated double-opponent cells and compute spatial average color gradients across simulated "resistive grids" akin to V4 neural tissue 63. Remarkably, when these biologically constrained architectures are exposed to naturalistic video stimuli or challenged with color assimilation illusions, they replicate the behavior of the human visual system, experiencing the same illusions and exhibiting robust, human-like color constancy in spatially varying illuminants 166566.
Deep Neural Networks as Digital Twins
Accessing the exact neurophysiological mechanics of area V4 in living subjects is constrained by the inherent limitations of fMRI spatial resolution and the invasive nature of single-unit neuronal recordings. Consequently, researchers have deployed deep neural networks (DNNs) to serve as "digital twins" of the visual cortex 6467.
By training sequential convolutional networks like DeepCC on millions of natural images and matching their hidden output layers to wide-field calcium imaging data from macaque monkeys, researchers can analyze the artificial network to deduce biological truths 646768. These deep learning models confirm that high levels of color constancy naturally emerge in intermediate network layers (analogous to V4) only when the system is allowed to utilize complex, long-range spatial inhibition and memory priors - validating the psychophysical theories derived from human illusions like the cyan strawberries and the viral dress 17646869.
The convergence of deep learning and visual neuroscience reinforces a central tenet of color perception: color is not a passive reception of environmental data, but an active, highly computed prediction of the world, reliant on a synthesis of immediate retinal wavelengths, learned lighting probabilities, and linguistic categorization 352868.