Neural mechanisms of in-group and out-group distinctions
The human brain possesses a profound, evolutionarily conserved capacity to categorize individuals into social groups, distinguishing swiftly between an in-group and an out-group. This categorization process is fundamental to navigating complex social environments, yet it is also the cognitive foundation of intergroup bias, prejudice, and systemic discrimination. The social neuroscience of group identity investigates how these social constructs are physically encoded in neural architecture. Through decades of functional neuroimaging, electroencephalography, and neuromodulatory interventions, researchers have mapped a distributed network of brain regions - encompassing the medial prefrontal cortex, the temporoparietal junction, the amygdala, and the striatum - that collectively govern how group boundaries are perceived, evaluated, and acted upon.
However, this field is currently undergoing a significant methodological and theoretical evolution. Early localizationalist assumptions, which posited direct mappings between single brain regions and complex social behaviors (e.g., characterizing the amygdala strictly as the brain's "fear center"), have been heavily contested. Similarly, foundational behavioral findings in social neuroendocrinology, particularly regarding the role of oxytocin in parochial altruism, are being reevaluated amidst a broader scientific replication crisis. Contemporary models emphasize that the neural boundaries of group identity are highly flexible, context-dependent, deeply modulated by cultural frameworks of self-construal, and demonstrably plastic in the face of cooperative intergroup contact.
Cognitive Foundations of Social Categorization
Categorization operates as an automatic cognitive reflex. Before higher-order evaluations regarding morality, intent, or trustworthiness occur, the brain must first encode a target as a social entity and assign that entity to a heuristic category. This early-stage processing relies heavily on visual perception networks and rapid structural encoding.
Visual Perception and Facial Processing
The most immediate social cue available to humans is the face. Behavioral and neuroscientific research extensively documents the "own-race bias" (also known as the cross-race effect), wherein individuals demonstrate superior recognition accuracy and emotion identification for faces of their racial in-group compared to out-group members 12. This bias is not merely a downstream memory failure but is rooted in the earliest stages of visual processing. Electroencephalography (EEG) studies reveal that novel group memberships modulate very early components of face processing, specifically the N170 event-related potential, which is implicated in the structural encoding of facial identity 3. Subjects display greater N170 amplitudes in response to in-group faces, suggesting that social identity motivates enhanced encoding of in-group members at a perceptual level within the first few hundred milliseconds of exposure 34.
Functional magnetic resonance imaging (fMRI) studies further localize this effect to the fusiform face area (FFA). The extended face-processing network exhibits heightened activation for in-group faces, which correlates directly with behavioral indices of individuation 3. Out-group faces, conversely, are often processed in a categorical rather than individuated manner, neurologically reinforcing the out-group homogeneity effect. The neurological allocation of attentional resources also exhibits cross-cultural variability. For instance, adult perceivers in Western contexts typically attend more to the eyes of racial in-groups but fixate on the nose and mouths of racial out-groups. Conversely, East Asian perceivers often show an attentional preference for the central facial area of in-groups, indicating that the neural mechanisms of face scanning are culturally imprinted and learned early in development 1.
The Minimal Group Paradigm
To isolate the neurological impact of group categorization from confounding variables like historical conflict, physical dissimilarities, or pre-existing cultural stereotypes, social neuroscientists frequently employ the Minimal Group Paradigm (MGP) 567. Originally pioneered by Henri Tajfel, the MGP arbitrarily assigns participants to groups based on trivial criteria, such as a coin flip or an aesthetic preference between the painters Klee and Kandinsky 8. Despite the meaningless nature of these groups and the lack of past or future interaction among members, participants reliably exhibit in-group favoritism in resource allocation tasks, trait evaluation, and implicit association measures 7910.
Participants in MGP studies demonstrate a tendency to maximize relative in-group gain over out-group gain (maximum differentiation), even when adopting this strategy requires sacrificing absolute in-group resource acquisition 810. Neuroimaging applications of the MGP demonstrate that arbitrary social classifications can engender in-group preference even when pitted against highly salient, orthogonal visual categorizations, such as race. When participants are assigned to mixed-race minimal groups, the neural processing of these novel affiliations can override automatic racial biases previously observed in the amygdala and FFA 346. This neural override suggests that while race is a pervasive social classifier due to historical inequalities and visual salience, it is not an intractable biological absolute; the brain's categorization networks are highly flexible and dynamically prioritize the most immediately relevant group boundary in a given social context 34.
Ecological Validity of Arbitrary Categorization
The MGP has faced methodological scrutiny regarding its ecological validity. Critics argue that real-world intergroup relations involve high stakes, deep historical contexts, and systemic power imbalances that artificial laboratory settings fail to replicate 510. Furthermore, while minimal group classifications can alter implicit evaluations, some neural activity - such as early event-related potentials (ERPs) - still exhibits an initial race effect before subsequent top-down processing redirects attention to the minimal group membership, indicating that physical salience retains a chronological primacy in perception 4.
Nevertheless, the MGP remains foundational because it successfully isolates the mere act of categorization from confounding sociological variables 5. A meta-analysis of over 200 studies indicated that the magnitude of bias observed in minimal groups is statistically comparable to that observed during interactions with real-world groups 5. Recent adaptations of the paradigm have further demonstrated its utility in exploring systemic issues, such as evaluating minimal group dynamics to simulate prejudice against migrants, revealing that the cognitive effort required to categorize displaced individuals independently drives lower favorability ratings 10.
Neural Architecture of Group Perception
The brain does not possess a singular "group identity" module. Instead, processing in-groups versus out-groups engages diverging pathways within the broader social cognition network, reflecting a fundamental neurobiological difference between mentalizing (attributing mental states) and salience detection.
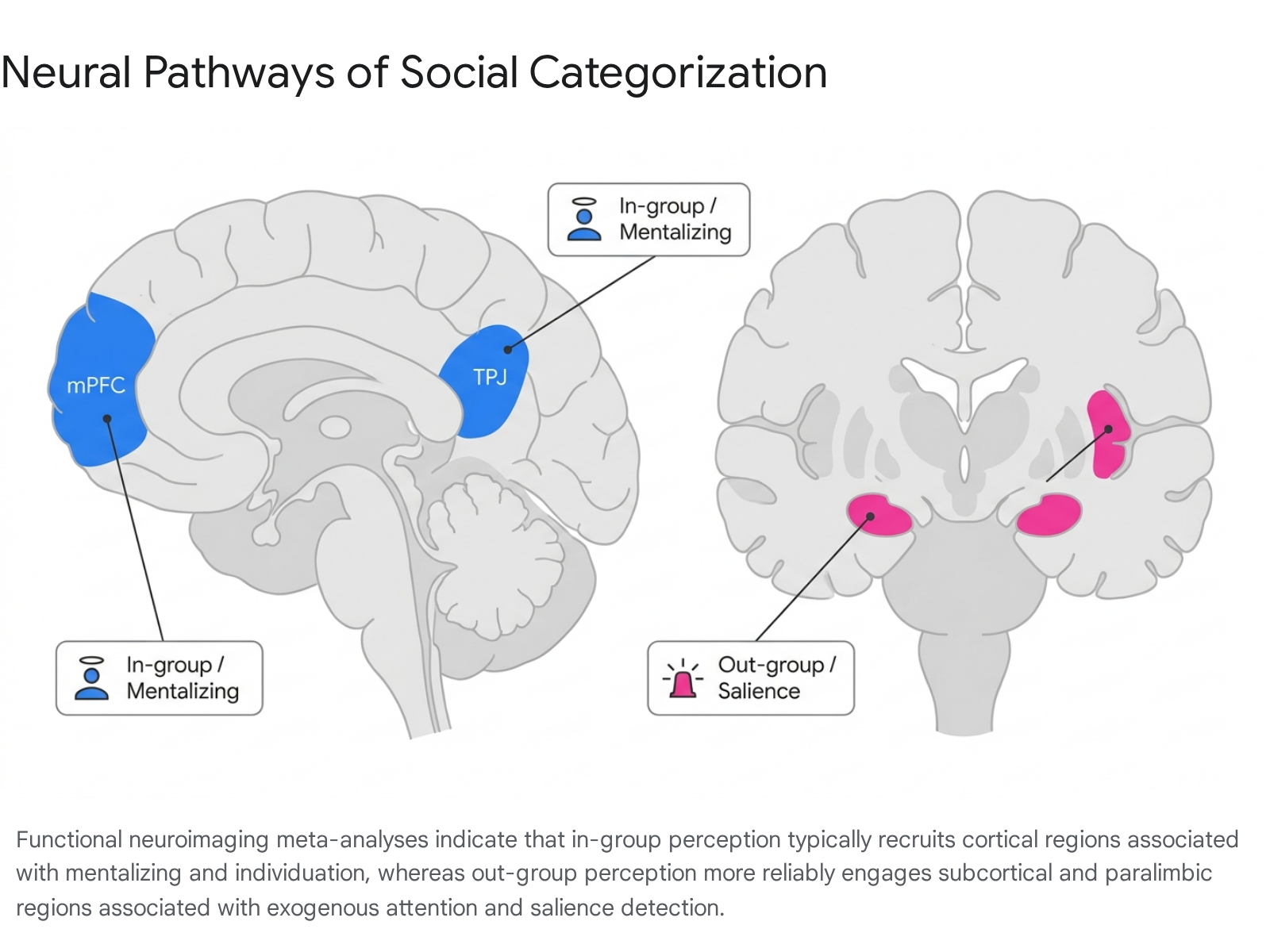
Medial Prefrontal Cortex and Temporoparietal Junction
The medial prefrontal cortex (mPFC) and the temporoparietal junction (TPJ) are the core hubs of the brain's mentalizing network, responsible for Theory of Mind (ToM) and cognitive perspective-taking 111213. Quantitative functional neuroimaging meta-analyses consistently reveal that social cognition directed toward in-group members more reliably elicits activation in these regions, particularly the dorsomedial prefrontal cortex (dmPFC), compared to out-group members 141516.
The mPFC exhibits functional subspecialization along its ventral-dorsal axis. The ventral mPFC (vmPFC) is primarily involved in bottom-up, evaluation-related processing, emotional regulation, and reward associations, while the dmPFC governs top-down, metacognitive processes, and perspective-taking 171819. When participants make judgments about themselves or in-group members, there is substantial overlap in vmPFC activation, suggesting that in-group members are processed through a neural proxy of the self 20. The preferential engagement of the mPFC and TPJ during in-group interactions facilitates individuation - the perception of targets as unique entities with rich internal lives - while out-group members frequently fail to recruit this network to the same extent, culminating in subtle forms of neural infrahumanization 46.
The Amygdala, Salience, and Novelty Encoding
Historically, social neuroscience strongly associated the amygdala with the processing of out-group faces, often interpreting this activation as evidence of implicit fear, threat detection, or negative evaluative bias 321. This perspective was heavily influenced by the amygdala's established role in fear conditioning and its frequent correlation with implicit racial bias scores 3. Consequently, early literature frequently inferred that heightened amygdala reactivity signaled an inherent, neurologically embedded prejudice against out-groups.
Contemporary interpretations of amygdala function in social categorization have evolved significantly, moving away from strict valence-based models toward salience-based models. The amygdala is increasingly understood as an encoder of general motivational salience and novelty 3222324. Large-scale meta-analyses demonstrate that both common and unusual novel stimuli engage the amygdala and hippocampus, as the region directs exogenous attention and cognitive resources to unpredictable environmental variables 242526. Therefore, heightened amygdala activation to out-group faces may reflect the perceptual novelty or unpredictability of the out-group rather than inherent animosity 25.
This interpretation aligns with findings that amygdala BOLD (blood-oxygen-level-dependent) responses correlate with the social rank of faces and hierarchical updating, tracking individuals who represent salient shifts in social dynamics regardless of specific emotional valence 27. In animal models, changes in social status trigger severe stress responses and rapid gene expression alterations within the medial amygdala, particularly for dominant individuals dropping in rank, underscoring the amygdala's role in mapping complex social hierarchies rather than simply generating fear 28. Furthermore, ecological momentary assessment (EMA) studies combined with fMRI reveal that individuals with heightened amygdala reactivity actually experience significant modulations in high-arousal positive affect depending on social intimacy contexts, providing further evidence against a strictly negative-valence framework for amygdala function 29.
| Neural Region / Network | Primary Cognitive Function | Intergroup Processing Bias | Meta-Analytic Findings 14161724 |
|---|---|---|---|
| Dorsomedial PFC (dmPFC) | Top-down mentalizing, perspective-taking | In-Group Bias | Greater activity when attributing mental states to in-group targets; reflects cognitive individuation. |
| Ventromedial PFC (vmPFC) | Bottom-up evaluation, self-referential processing | In-Group Bias | High functional overlap with self-processing networks; integrates the affective value of the in-group. |
| Temporoparietal Junction (TPJ) | Theory of Mind, distinguishing self from other | In-Group Bias | Modulated by inter-brain plasticity during successful in-group cooperation and affective empathy. |
| Amygdala | Salience, arousal, novelty detection | Out-Group Bias | Rapidly encodes out-group boundaries due to perceptual novelty and unpredictable social salience. |
| Anterior Insula | Exogenous attention, affective resonance | Out-Group Bias | Consistently activated in contrast mappings of out-group vs. in-group social cognition tasks. |
| Ventral Striatum | Reward processing, value computation | In-Group Bias | Tracks the subjective reward value of interacting with or observing the success of in-group members. |
Ventral Striatum and Reward Circuitry
In addition to the mentalizing network, in-group favoritism is heavily supported by the brain's reward circuitry. The ventral striatum demonstrates significantly greater activity in response to in-group members, reflecting the inherently rewarding nature of in-group affiliation and interaction 1416. This dopaminergic response underlies parochial altruism - the willingness to incur personal costs to benefit the in-group - and reinforces behavioral loops that maintain group cohesion.
Cultural Modulation of Neural Encoding
The baseline neural pathways that differentiate in-groups from out-groups are not biologically fixed across all human populations. They are dynamically modulated by macro-level cultural paradigms. A significant critique of foundational social neuroscience is its overreliance on WEIRD (Western, Educated, Industrialized, Rich, Democratic) populations, which champion individualistic values and potentially skew generalized models of human cognition 3031.
Individualism and Collectivism Paradigms
Recent cross-cultural neuroimaging reveals that the cultural dimension of individualism versus collectivism fundamentally alters how the brain encodes the self and, by extension, the in-group 13132. In individualistic cultures, the self is perceived as a stable, autonomous entity, independent of social context. Conversely, in collectivistic cultures, the self is highly relational, dynamic, and defined by social obligations and group belonging 13233.
fMRI studies directly testing these populations demonstrate that cultural values modulate neural activity within the mPFC. Individualistic subjects exhibit greater activation in the anterior rostral portion of the mPFC during general self-judgments, reflecting an autonomous self-concept 3233. In contrast, collectivistic subjects show greater mPFC activation during contextual self-judgments, mirroring an interdependent self-concept 3233.
When navigating intergroup boundaries, members of collectivist societies often allocate greater neural resources to maintain group harmony. However, they may also display stronger, more inflexible relational boundaries. Studies using cyberball exclusion paradigms show that individuals from marginalized or collectivist-leaning groups often demonstrate heightened neural responses (e.g., in the insula and prefrontal networks) to out-group exclusion compared to highly individualistic subjects 34. Because the in-group is structurally integrated into a collectivist's neural representation of the self, the boundaries of in-group encoding can be deeply entrenched, yet they remain highly sensitive to situational relationship dynamics 135.
Cognitive Load and Contextual Scanning
These cultural frameworks extend to baseline cognitive processing and visual encoding. Research indicates that collectivist cognition often relies heavily on holistic and contextual processing, whereas individualist cognition relies on analytic, object-focused processing 213637. When encoding complex visual scenes, Westerners tend to focus on central objects (engaging visual regions efficiently), while East Asians spontaneously attend to backgrounds and relationships 2137. Forcing subjects to adopt the non-preferred visual scanning strategy significantly increases cognitive burden and frontal lobe activation, demonstrating that cultural paradigms physically alter the metabolic demands of basic perception 3738.
Intergroup Empathy and Neural Plasticity
The divergence in neural encoding between in-groups and out-groups has profound implications for interpersonal emotion regulation and empathy. Empathy is a multidimensional construct encompassing both an affective component (shared emotional resonance, heavily reliant on the anterior cingulate cortex [ACC] and anterior insula) and a cognitive component (perspective-taking, reliant on the mPFC and TPJ) 1119.
The Intergroup Empathy Gap
Humans exhibit a stark "empathy gap" across group boundaries. When observing an in-group member experiencing physical or emotional pain, individuals demonstrate robust, synchronous activation in both the affective and cognitive empathy networks 35. However, observing an out-group member in identical distress elicits significantly dampened empathic resonance in these specific regions 35. In competitive intergroup contexts, observing out-group suffering can even activate the ventral striatum, indicating a neural signature of schadenfreude (pleasure at another's misfortune) 35. This failure of intergroup empathy is considered a proximate neurobiological cause of out-group indifference and is theorized to facilitate intergroup aggression by bypassing the neural constraints that normally inhibit harm against others 35.
Mental Training and Structural Plasticity
Despite the automaticity of these biases, the neural architectures underlying group identity exhibit high plasticity. Intergroup contact theory posits that sustained, cooperative contact with out-group members reduces prejudice, a phenomenon observable at the neurobiological level.
Longitudinal fMRI studies evaluating mindfulness and compassion-based mental training interventions (MBIs) demonstrate significant structural and functional plasticity within the social brain network 13. Training programs focused specifically on affective empathy have been shown to enhance fronto-insular connectivity, fostering greater emotional resonance. Conversely, training modules focused on cognitive perspective-taking selectively boost plasticity in the TPJ, inferior frontal gyrus (IFG), and mPFC 13.
As individuals practice perspective-taking with out-group members, the functional connectivity between the prefrontal cortex and the amygdala undergoes dynamic shifts. Top-down regulatory signals from the dmPFC can modulate the amygdala's initial salience response, dampening heuristic fear or novelty responses and allowing the perceiver to bypass category-based encoding to engage in target individuation 364142. Neuromodulatory interventions, such as transcranial direct current stimulation (tDCS) targeting the mPFC, have been shown to experimentally alter the representation of shared-goal beliefs, further implicating the mPFC's causal role in generalizing positive actions across group members 36. Furthermore, studies on adolescents with histories of childhood maltreatment reveal that structured interventions can induce functional developmental plasticity in cortical midline structures (such as the retrosplenial cortex and vmPFC), suggesting that the neural basis of social resilience and in-group attachment remains malleable well into adulthood 37.
Inter-Brain Plasticity and Hyperscanning
The frontier of social neuroscience is moving beyond the isolated individual to examine interacting dyads and groups. "Inter-brain plasticity" refers to the ability of interacting brains to modify their neural coupling over time in response to repeated social exchanges 1138. Using hyperscanning methodologies (recording simultaneous brain activity from multiple subjects via EEG or fNIRS), researchers have observed that successful group cooperation induces significant neural synchronization, particularly in the orbitofrontal cortex (OFC) and dorsolateral prefrontal cortex (DLPFC) 39. Stronger group identification correlates with increased individual DLPFC activation and subsequent alignment of inter-brain networks, suggesting that shared social identity actively coordinates multi-brain neurocognitive states to optimize collective performance 39.
Methodological Constraints in Neuroimaging
Our understanding of the social neuroscience of group identity is intricately bound to the limitations of the tools and methodologies used to measure it. The field must carefully navigate the inherent physical constraints of neuroimaging technology to draw accurate inferences regarding rapid social categorization.
Spatial and Temporal Limitations of fMRI
fMRI remains the dominant modality for mapping the social brain due to its excellent spatial resolution, allowing precise localization of activity in deep subcortical structures like the amygdala and striatum 4047. However, fMRI relies on the blood-oxygen-level-dependent (BOLD) signal, an indirect hemodynamic response that peaks several seconds after the underlying neural firing. This results in notoriously poor temporal resolution 40474142.
The temporal lag of the BOLD signal poses a significant challenge for social categorization research, which involves neurocognitive processing occurring within milliseconds 34143. For example, determining whether the amygdala's response to an out-group face chronologically precedes or follows top-down modulation from the prefrontal cortex is mathematically complex when data is aggregated over the sluggish hemodynamic response 3. To address this, researchers apply advanced modeling techniques, such as spatiotemporal constrained principal component analysis (ST-fMRI-CPCA), to portion variance predictable from finite impulse response models and extract precise task-based networks 51.
Addressing Voxel Averaging
Spatial constraints present another challenge: a single fMRI voxel represents millions of neurons. Activation within a voxel might obscure distinct, overlapping neural subpopulations. For instance, overlapping activation for different social categories within the mPFC might be misconstrued as identical processing. To circumvent this, researchers increasingly employ repetition suppression paradigms (fMRI adaptation). This technique exploits the biological phenomenon that stimulus-tuned neurons reduce their firing rate upon repeated exposure to their preferred stimulus. By measuring reduced BOLD responses to repeated distinct stimuli, researchers can infer that both stimuli share a cognitive property processed by the same sub-voxel neuronal population, allowing for sub-voxel resolution in social categorization tasks 44.
To bridge the spatiotemporal gap entirely, the field is rapidly moving toward multimodal integration, simultaneously combining fMRI with EEG or functional near-infrared spectroscopy (fNIRS). This integration captures the high temporal resolution of electrical activity alongside the high spatial resolution of hemodynamics, tracking the rapid, sequential cascade of social encoding from initial visual perception to complex moral judgment 4546.
The Replication Crisis in Social Neuroscience
Perhaps the most disruptive event in contemporary social psychology and neuroscience has been the replication crisis, which has forced a severe reevaluation of foundational claims regarding the neuroendocrinology and cognitive priming of group behavior 4748. Sparked largely by the Open Science Collaboration in 2015, widespread efforts to reproduce classic psychological findings revealed that a substantial portion of studies failed to replicate, suffering from low statistical power, positive effect size bias, and questionable research practices (such as p-hacking and selective data omission) 47575849.
The Reevaluation of Oxytocin
A prominent casualty of the replication crisis within social neuroscience is the literature surrounding oxytocin (OT). In the mid-2000s, high-profile studies published in premier journals claimed that the administration of intranasal oxytocin fundamentally altered intergroup dynamics 5061. OT was widely branded as the "trust hormone," with researchers asserting it operated via "parochial altruism" - increasing in-group trust and cooperation while simultaneously driving defensive aggression, derogation, or preemptive strikes against out-groups 506151.
Subsequent large-scale, highly powered replication attempts spanning the last decade have systematically failed to reproduce these effects 616352. In one notable example, the "Envelope Task" - a behavioral paradigm where trust is measured by the degree of openness of an envelope containing confidential information, previously cited as showing OT's massive effect on trust - completely failed to replicate across multiple independent labs with robust sample sizes 635253. Meta-analyses of OT's effect on trust in behavioral economic paradigms yield combined effect sizes that are small and not reliably different from zero 53.
The persistence of the original oxytocin claims in popular science, despite failed replications, highlights the impact of publication bias (the "file-drawer" effect), wherein high-impact journals heavily favor publishing novel, statistically significant findings while rejecting null replications 4961. If oxytocin does influence group identity, recent rigorous studies suggest its effects are highly conditional, interacting with pre-existing individual differences rather than acting as a universal neurochemical switch. For instance, OT's modulation of in-group favoritism during economic cooperation appears to depend heavily on an individual's cognitive processing style, increasing in-group bias in intuitive thinkers but actively decreasing it in reflective thinkers 54. Similarly, OT has been shown to reduce feelings of agency in anxiously attached individuals, suggesting complex, context-dependent outcomes 67.
| Methodological Vulnerability | Impact on Social Neuroscience | Corrective Action / Current Standard |
|---|---|---|
| Low Statistical Power & Publication Bias | High rates of false positives; inflated effect sizes; persistence of discredited theories (e.g., universal Oxytocin trust effects) 584961. | Pre-registration of hypotheses; large-scale, multi-site collaborative sample collections; publication of null results 4955. |
| Temporal Resolution of fMRI | Inability to track millisecond-level directional flow between the mPFC and Amygdala during rapid social categorization 4741. | Multimodal imaging (fMRI + EEG/fNIRS); application of advanced latency structure models and ST-fMRI-CPCA 40514546. |
| Spatial Resolution (Voxel Averaging) | Overlapping activation across categories within a single brain region obscures distinct neural subpopulations 44. | Repetition suppression paradigms; Multi-voxel pattern analysis (MVPA) to decode specific representations 44. |
| WEIRD Sample Bias | Neural mechanisms of self and group construed as universal, ignoring cultural plasticity 131. | Cross-cultural neuroimaging directly testing Individualist vs. Collectivist cohorts on cognitive load and self-construal 32333437. |
Conclusion
The social neuroscience of group identity has revealed that the human brain possesses distinct, highly specialized mechanisms for distinguishing in-groups from out-groups. The neural architecture leans heavily on the medial prefrontal cortex and temporoparietal junction to cognitively individuate and empathize with in-group members. Conversely, out-groups are frequently processed through salience and novelty networks centered on the amygdala and anterior insula.
Crucially, these neural boundaries are neither biological imperatives nor rigidly fixed. They are highly dependent on immediate context, deeply modulated by macro-cultural frameworks of self-construal, and demonstrably plastic in the face of cooperative intergroup contact and targeted mental training. As the field actively overcomes its historical replication crises by adopting open science frameworks, and embraces sophisticated network-level and multimodal analyses, it moves closer to providing a robust, highly accurate framework for understanding - and potentially mitigating - the deep-seated neural mechanisms that drive intergroup bias.