Neural mechanisms of aesthetic perception
Foundations of Neuroaesthetics
The biological and cognitive mechanisms governing the human appreciation of beauty constitute the foundation of neuroaesthetics, an interdisciplinary field situated at the convergence of cognitive neuroscience, evolutionary psychology, and art history 123. While aesthetic inquiry was historically confined to philosophical discourse regarding the nature of beauty and subjective taste - dating back to Aristotelian concepts of proportion and Kantian notions of the sublime - the advent of advanced neuroimaging techniques has facilitated the empirical measurement of aesthetic experiences 343. Utilizing functional magnetic resonance imaging (fMRI), electroencephalography (EEG), magnetoencephalography (MEG), and functional near-infrared spectroscopy (fNIRS), researchers can now quantify the temporal dynamics and spatial architecture of human aesthetic perception 2475.
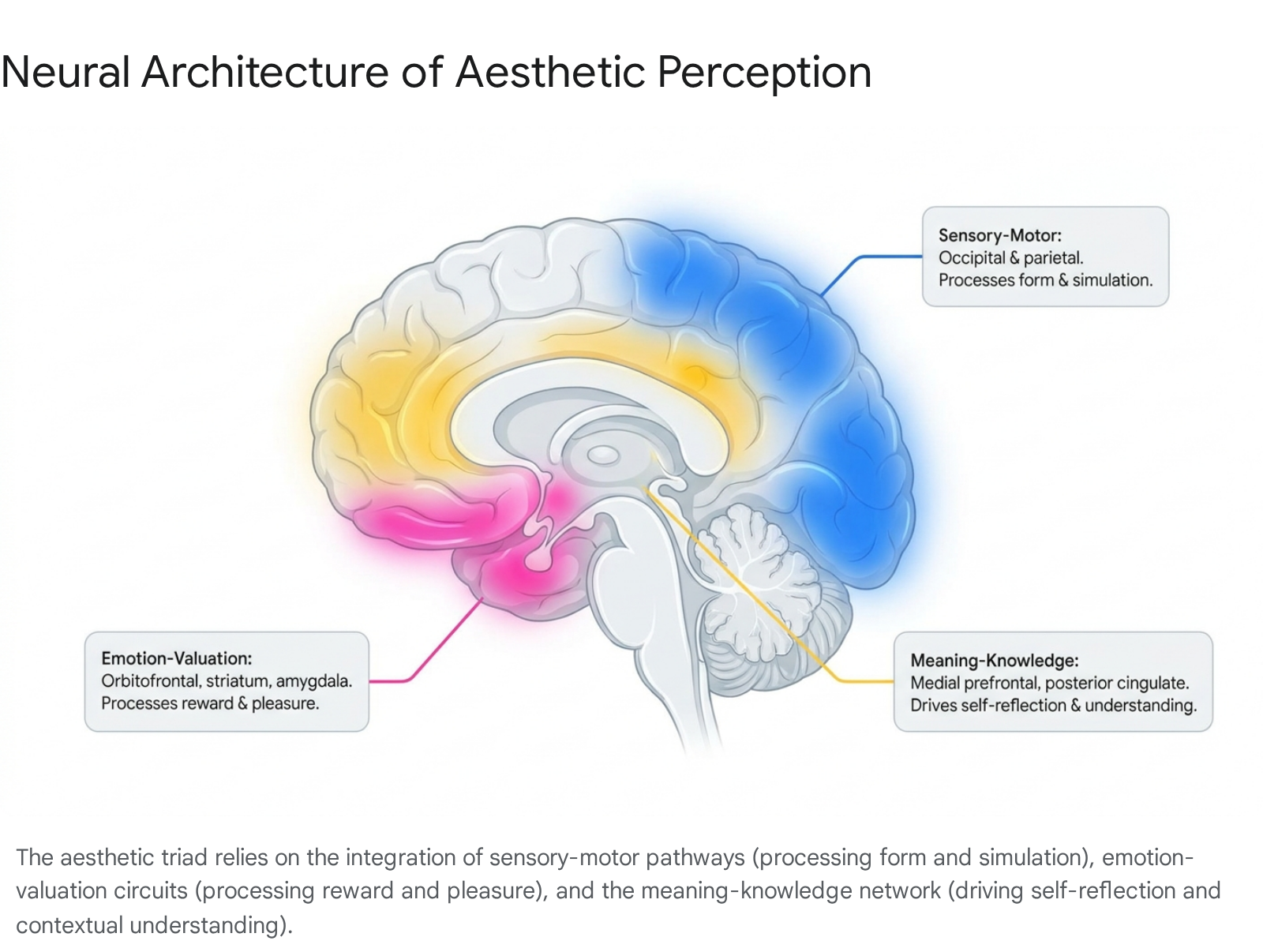
Neuroaesthetics posits that the desire to create, experience, and respond to beauty is biologically rooted and profoundly influences cognitive states, emotional regulation, and social behavior 2. Current research indicates that the experience of beauty is not localized to a singular "beauty center" in the brain. Instead, aesthetic appreciation emerges from the dynamic interplay of complex, large-scale neural networks 96. This systems-theoretical approach defines the aesthetic experience through an "aesthetic triad" comprising three interacting neural systems: the sensory-motor system, the emotion-valuation system, and the meaning-knowledge system 37.
The sensory-motor system processes the physical features of stimuli, extracting data regarding symmetry, color, spatial frequency, and motion from the occipital and parietal cortices, while also facilitating embodied motor resonance 27. Concurrently, the emotion-valuation system, encompassing the mesocorticolimbic reward circuitry such as the orbitofrontal cortex, ventral striatum, and amygdala, assigns hedonic value and emotional weight to the perceived object 77. Finally, the meaning-knowledge system, heavily reliant on the Default Mode Network and executive control regions, integrates cultural context, personal identity, and semantic understanding to formulate higher-order judgments 237. Together, these mechanisms transform raw sensory input into an emotionally charged, meaningful aesthetic experience.
Evolutionary Psychology of Facial Beauty
The human brain demonstrates a highly specialized and remarkably rapid capacity for processing facial beauty, a mechanism deeply rooted in evolutionary biology and survival mechanics. The neurological evaluation of an attractive face occurs in approximately 13 milliseconds, operating faster than conscious awareness 12. This speed underscores the evolutionary imperative of rapid social evaluation, wherein physical beauty historically served as a proxy for genetic fitness, robust health, and reproductive viability 1128.
Biological Markers of Facial Attractiveness
Cross-cultural and evolutionary psychology research reveals a universal baseline for facial attractiveness driven by specific geometric and biological markers 12. Mathematical averageness and bilateral facial symmetry are consistently rated as highly attractive across diverse populations 1412. Asymmetry in nature is frequently an indicator of developmental disorders or parasitic infections; thus, symmetric faces signal robustness against environmental and genetic stressors 1. Furthermore, composite faces - images generated by digitally blending multiple individual faces - are routinely evaluated as more attractive than singular individual faces because their structural averageness implies greater genetic diversity and heterozygosity 1.
Sexual dimorphism also plays a critical role in aesthetic appraisal. Traits associated with heightened fertility and hormonal health, such as large eyes and high cheekbones in females, or low brows and strong jawlines in males, elicit strong aesthetic responses 1. Beyond structural anatomy, recent fMRI research indicates that skin quality operates as an independent modulator of aesthetic reward. Changes in skin luminosity and clarity, even without any alterations to underlying facial geometry, significantly influence perceived attractiveness and activate the brain's reward centers 8. Radiant skin signals vitality and systemic health, providing a direct neurological bridge between dermatological aesthetics and the observer's emotional processing 8.
Reward Circuitry and Neural Processing
Attractive faces trigger the dopamine-driven reward system, mirroring the neural mechanics observed in responses to primary survival rewards such as food. When individuals view beautiful faces, the medial orbitofrontal cortex (mOFC) exhibits heightened activation 1512. This activation occurs automatically and continuously; even when subjects are instructed to focus exclusively on identifying the face rather than judging its beauty, the OFC still illuminates in the presence of attractive features 1.
The aesthetic evaluation of faces involves specific regions that separate generalized reward from face-specific beauty. The lateral OFC is selectively activated for facial beauty over landscape beauty, indicating domain-specific processing pathways for evaluating conspecifics 5. Correspondingly, activation in the nucleus accumbens, ventral striatum, and anterior cingulate cortex scales linearly with the perceived attractiveness of faces and bodies, providing a quantifiable neural index of aesthetic valuation 75.
The Intersection of Aesthetics and Morality
The neural preference for facial beauty extends deeply into complex social and moral judgments, manifesting in cognitive biases such as the "halo effect." Faces deemed structurally attractive often elicit automatic attributions of positive personality traits, such as honesty, trustworthiness, and kindness, as the brain assumes those who are aesthetically pleasing are also beneficial to social survival 8.
Conversely, the aesthetic processing of facial anomalies - such as scars, disfigurement, or paralysis - triggers implicit biases and distinct neural responses. Observers evaluating faces with anomalies display neural activation patterns indicative of diminished empathy, evaluate the character of the individual more negatively, and demonstrate stronger "just-world" beliefs (the psychological assumption that people get what they deserve) 19. These problematic stereotypes highlight how deeply hardwired aesthetic algorithms can inappropriately influence higher-order social valuation, moral reasoning, and even lead to implicit dehumanization 9.
Cultural Mediation of Facial Aesthetics
While symmetry and averageness are largely universal drivers of attractiveness, cultural context heavily modulates the weighting of specific aesthetic variables. Studies evaluating facial beauty across non-WEIRD (Western, Educated, Industrialized, Rich, and Democratic) and Western populations reveal divergent secondary preferences. For instance, in cross-cultural evaluations involving participants from Mozambique, Brazil, and France, researchers found broad agreement on the salience of proportionality, but distinct cultural differences emerged regarding preferred facial breadth and profile thinness, with French participants exhibiting a unique preference for thinner facial profiles 10.
Furthermore, cultural ideals often intersect with physiological markers to shape conscious preference. In some sub-Saharan contexts, skin tone has been shown to impact attractiveness judgments independently of symmetry, although structural symmetry generally accounts for a vastly larger percentage of the variance in attractiveness ratings (e.g., 85% of total variance attributed to symmetry compared to 5% for skin tone in specific experimental parameters) 10. This underscores a dual-process model of facial aesthetics: an evolutionary, innate neural mechanism prioritizing symmetry and health markers, heavily overlaid by culturally acquired and socially reinforced preferences 312.
Visual Art and Default Mode Network Dynamics
While human responses to facial beauty and natural landscapes exhibit high cross-cultural consistency, the aesthetic appreciation of cultural artifacts - such as visual art and complex architecture - is marked by extreme individual variability 111213. Artifact-based culture has existed for a relatively short evolutionary timeframe (approximately 45,000 years), resulting in visual art preferences that are highly individualized and inextricably tied to personal identity, semantic interpretation, and cultural exposure 1114. The core neural substrate that mediates this highly subjective aesthetic experience is the Default Mode Network (DMN) 121516.
Redefining Network Dynamics
The DMN comprises a set of heavily interconnected brain regions, including the medial prefrontal cortex (mPFC), posterior cingulate cortex (PCC), precuneus, and temporoparietal junction 1718. Originally characterized as a "task-negative" network, the DMN is typically highly active during periods of rest, daydreaming, autobiographical memory retrieval, and self-referential thought 111214. Standard cognitive models dictate that the DMN must be suppressed when an individual focuses attention on external stimuli, allowing the executive control network (ECN) and sensory cortices to process the outside world effectively 1113.
However, neuroaesthetics has revealed a paradoxical mechanism during profound aesthetic experiences. When viewers observe artworks they rate as highly "moving" or exceptionally beautiful, DMN deactivation is alleviated. Instead of suppressing the DMN to process the external painting, the brain actively engages the DMN simultaneously with the visual cortices 111214. This engagement suggests that moving artworks gain access to neural substrates concerned with the self, effectively signaling "self-relevance" 121419. The viewer processes the external artifact through an internal lens, deriving "pleasure from understanding" and integrating the artwork into their personal narrative and emotional history 1116.
Domain-General vs. Domain-Specific Coding
A critical debate in neuroaesthetics is whether the brain houses a universal module that evaluates all aesthetic domains equally, or if different types of beauty are processed in isolated, domain-specific networks 915. Multivoxel pattern analysis (MVPA) of fMRI data has elucidated this architecture by separating the processing of stimulus content from the processing of aesthetic appeal 1315.
When humans view different categories of images - such as natural landscapes, architecture, or abstract paintings - the ventral occipitotemporal cortex (VOT) exhibits highly domain-specific coding. The VOT accurately represents the physical content of what is seen but does not generalize aesthetic appeal across categories 1315. In direct contrast, the DMN contains a domain-general representation of visual aesthetic appeal. Activity patterns within the DMN can accurately predict whether an observer finds an image beautiful, regardless of whether the image is a landscape, a portrait, or a cultural artifact 1315.
| Neural Network / Region | Aesthetic Processing Function | Specificity | Primary Associated Cognitive Processes |
|---|---|---|---|
| Ventral Occipitotemporal Cortex (VOT) | Object and Scene Recognition | Domain-Specific | Encodes the physical content (e.g., distinguishing architecture from a human face). Does not generalize beauty across categories. 1315 |
| Default Mode Network (DMN) | Self-Relevance & Meaning | Domain-General | Predicts overall aesthetic appeal across diverse visual domains. Active during intense, "moving" aesthetic immersion. 121315 |
| Orbitofrontal Cortex (OFC) | Reward & Hedonic Valuation | Stimulus-Dependent | Tracks the magnitude of pleasure and reward. The lateral OFC shows distinct selectivity for faces over landscapes. 15 |
| Lateral Frontopolar Cortex (FPl) | Complex Environmental Evaluation | Domain-Specific | Decomposes and parallel-processes complex environmental features during spatial and landscape selection. 20 |
This architecture supports a feed-forward model of aesthetic appreciation: domain-specific sensory representations in high-level visual areas feed into a core, domain-general system located in the DMN and subcortical reward circuitry, where the final computation of aesthetic value, pleasure, and self-relevance is executed 1315.
Temporal Architecture of Art Appreciation
Aesthetic experiences are not instantaneous; they unfold dynamically over time. Unlike the rapid 13-millisecond processing of facial attractiveness, the appreciation of complex art demands slow temporal dynamics 122122. Time-series fMRI analyses reveal that within the first few seconds of viewing a highly pleasing artwork, activity spikes simultaneously in the DMN, high-level visual areas, and reward regions, suggesting a partially automatic initial valuation 2122.
As the viewing duration extends, a suppressive effect occurs across the network. However, a defining signature of a profound aesthetic experience emerges at the exact moment the stimulus is removed. For highly pleasing art, the DMN response exhibits a synchronized return to baseline that is strictly time-locked to the image offset 2122. Conversely, when viewing non-pleasing art, the network dynamics are inconsistent, fragmented, and disengaged from the visual stimulation 2122. This temporal trajectory indicates that the DMN actively tracks the viewer's internal state of aesthetic immersion, continuously updating the balance between externally directed attention and internally directed meaning-making 2122.
Visual Ambiguity and Negative Aesthetics
Modern art frequently employs visual ambiguity to challenge the viewer, requiring complex cognitive resolution. The evaluation of ambiguous artistic photographic stimuli is specifically associated with stronger fixation-related neural activity in the left medial prefrontal cortex, alongside increased right fronto-parietal and occipito-temporal activity 23. This pathway supports the resolution of visual ambiguity through attentional reorienting, navigating between the global and local aspects of the artwork, indicating that the cognitive challenge itself forms part of the aesthetic reward 23.
Conversely, neuroaesthetics has historically neglected negative aesthetic evaluations, focusing overwhelmingly on beauty. Recent meta-analyses evaluating neural responses to displeasing art and negative imagery from the International Affective Picture System (N-IAPS) highlight the role of the right fusiform gyrus 24. While viewing explicitly negative non-artistic imagery activates widespread frontal, temporal, and subcortical threat areas, negative aesthetic evaluations of art (i.e., finding a painting ugly rather than frightening) predominantly isolate activity to the right fusiform gyrus encroaching on the anterior cerebellum 24. This implies that domain-general affective systems process visual displeasure, but the specific context of stimulus engagement modulates the severity and spread of the neural response 24.
Cultural Enculturation in Art Perception
While the neural mechanics of the DMN and reward systems remain consistent across human populations, the specific aesthetic content that triggers these networks is heavily mediated by cultural enculturation. Art appreciation relies significantly on processing fluency - the ease with which the brain decodes information - which is shaped by predictive models based on cultural background 16.
When analyzing brain activation in native Chinese and European participants viewing traditional Western and Chinese landscape paintings, fMRI data reveals a distinct intracultural bias 25. Participants exhibited significantly stronger brain activation to artistic expressions originating from their own culture 25. Western and Eastern artists historically employ divergent perspectives to represent the visual world - such as linear perspective in Western art versus reversed-geometric, multi-point perspective in traditional Chinese landscapes 25. The brain responds more robustly to familiar aesthetic vocabularies because repeated cultural exposure shapes the neural templates used to decode meaning and formulate concepts of beauty 625.
Embodied Cognition and Motor Simulation
The aesthetic experience is fundamentally rooted in the body. The framework of embodied cognition and neuroaesthetics suggests that perceiving visual art is not merely an act of passive visual observation but involves active, albeit covert, motor simulation 726. The brain perceives the actions, emotions, and implied movements captured in a static artwork by simulating them within the observer's own motor and visceromotor networks 72728.
The Mirror Neuron System in Aesthetics
The neurological foundation of embodied neuroaesthetics lies in the Mirror Neuron System (MNS), primarily encompassing the ventral premotor cortex (vPMC), posterior parietal cortices (PPC), and the superior temporal sulcus (STS) 729. When an observer views an artwork depicting dynamic human actions - such as a sculpture of a figure in mid-stride or a painting of a complex dance - the MNS is automatically activated 729. The observer's brain essentially simulates the implied movement physically. Transcranial magnetic stimulation (TMS) studies corroborate this effect, demonstrating increased corticospinal excitability in observers viewing dynamic paintings, which directly correlates with the perceived aesthetic preference and dynamism of the work 29.
This embodied simulation extends beyond explicitly represented human figures to the traces of the artist's physical gestures left upon the canvas. Known as "Embodied Calligraphic Resonance," this phenomenon is highly evident in the aesthetic appraisal of mediums like traditional Chinese calligraphy or abstract expressionist painting 626. Viewers do not simply process the static stroke as a geometric shape; their motor cortices implicitly reconstruct the dynamic movement, speed, and physical vitality of the brushstroke originally generated by the artist 626. This motor familiarity is particularly strong when professional artists observe works within their own domain of expertise, relying heavily on embodied pathways rather than merely perceptual learning 2636.
Liberated Simulation and Empathy
In the specific context of engaging with art, embodied simulation is often described in neuroaesthetic literature as "liberated" 30. Because the viewer's higher-order cognitive systems recognize the artwork as a representation or fiction rather than an immediate physical reality, the simulation is freed from the burden of executing actual survival-oriented actions 30. This aesthetic distance allows the viewer to safely experience intense affective states and bodily sensations without defensive withdrawal.
This liberated mechanism is crucial for the generation of deep empathy. Witnessing pain, struggle, or intense emotion depicted in an artwork recruits the anterior insula and the anterior cingulate cortex - the identical neural circuitry involved in experiencing first-hand emotion and physical pain 7. Therefore, aesthetic appreciation serves as an epistemic affordance, allowing individuals to bridge the gap between their own somatic experiences and the subjective experiences expressed by the artist, expanding the boundaries of the viewer's peripersonal space 7.
Environmental Aesthetics and Neurobiophilia
The human affinity for natural environments is ubiquitous and was historically framed by the "biophilia hypothesis," which posits an innate, evolutionary drive to affiliate with nature and other living systems 313233. Recent developments in environmental neuroscience have translated this psychological theory into the empirical framework of "Neurobiophilia" 3233. Brain imaging studies consistently demonstrate that exposure to nature, as opposed to urban or artificial environments, yields distinct neurophysiological benefits ranging from enhanced top-down attentional control to accelerated stress recovery 174142.
Mechanisms of Fractal Fluency
A primary driver of nature's restorative capacity is its underlying geometric structure, specifically the ubiquitous presence of fractal patterns. Fractals are self-similar structures that repeat at varying scales of magnification, found heavily in nature in the form of tree branches, cloud formations, coastlines, and leaf veins 314334.
The "fractal fluency" theory suggests that over millions of years of evolution in natural environments, the human visual system adapted to process these specific geometries with maximum efficiency and minimal metabolic cost 314345. Nature primarily consists of mid-range fractals, mathematically defined by a fractal dimension ($D$) falling between 1.3 and 1.5 433536.
Neurophysiological data indicates that viewing mid-$D$ fractals triggers a distinct, optimized biological response: - EEG Oscillations: Exposure to mid-range fractals induces a significant increase in alpha wave activity (8 - 12 Hz) in the brain, an electrophysiological marker associated with a relaxed yet highly alert state conducive to creative insight 414837. - fMRI Activation: Mid-$D$ fractals selectively activate attention and reward-related circuits while demanding measurably less metabolic effort from early visual processing areas, facilitating fluid pattern recognition without cognitive fatigue 4334. - Autonomic Regulation: Autonomic nervous system monitoring reveals that visual exposure to fractal patterns leads to a rapid, measurable reduction in cortisol levels, lowered blood pressure, and balanced heart rate variability (HRV) 413435.
When humans are placed in artificial environments devoid of these scale-invariant structures, the brain struggles to process the visual field, leading to increased cognitive load and subsequent physiological stress 34.
Neurobiophilia and Environmental Enrichment
The Neurobiophilia framework categorizes the brain's interaction with nature into seven hierarchical "neuro-needs" (7NNs): neuroprotection, neurophysiological regulation, neurotrophic factor stimulation, and neurogenesis, among others 3233. Extended exposure to immersive natural environments - and specifically environments combining dense vegetation with expansive skies - activates unique neural pathways. fMRI studies utilizing prolonged exposure to sky compositions reveal specific activations in regions associated with spatial cognition, circadian rhythm regulation, and the perception of expansive space, pathways completely untriggered by generalized positive artificial stimuli or traditional interior ceilings 38.
Evidence from translational neuroscience highlights nature's vital role in upregulating Brain-Derived Neurotrophic Factor (BDNF), a critical protein supporting adult hippocampal neurogenesis 323339. The absence of nature access due to rapid modern urbanization creates an environmental deprivation that impairs these neurogenic mechanisms, increasing population-level vulnerability to depressive symptoms, stress reactivity, and cognitive decline 3940.
Comparative Immersive Environments
The cognitive benefits of specific environmental aesthetics have been tested directly through fully immersive behavioral studies. A prominent 2025 study (Holzman et al.) tested human aesthetic appreciation, executive functioning, and creativity across three distinct settings: a real natural forest, an indoor room designed with biophilic elements (e.g., moss walls, fir wood, fractal rugs, diffuse light), and a standard control laboratory room 414243.
Aesthetic appreciation - measured across dimensions of coherence, fascination, and hominess - was highest in the natural forest, followed by the biophilic room, and lowest in the control room 414243. However, the study found null results regarding differences in standard attention, working memory, or mood across the three environments, suggesting that brief exposures may not sufficiently alter baseline mood or rote memory 4243. Crucially, the study did reveal that participants were significantly more creative in the natural environment than in either indoor space, demonstrating heightened cognitive flexibility and superior divergent thinking capabilities when immersed in actual nature 4243.
| Experimental Environment | Aesthetic Appreciation Score | Impact on Divergent Creativity | Impact on Rote Attention / Mood |
|---|---|---|---|
| Natural Forest | Highest (Maximized fascination and coherence) | Significant Improvement (High cognitive flexibility) | Null (No measurable deviation from baseline) |
| Biophilic Indoor Room | Moderate (Superior to standard controls) | Marginal / Baseline | Null (No measurable deviation from baseline) |
| Control Laboratory | Lowest (Lacking natural geometric patterns) | Baseline | Null (No measurable deviation from baseline) |
| Data derived from Holzman et al. (2025) comparative environmental study 414243. |
Spatial Cognition and Neuroarchitecture
Neuroarchitecture applies the principles of neuroaesthetics and biophilia to the built environment, examining how structural parameters physically shape brain function 4445. Modern urban living frequently subjects the human brain to "allostatic overload" - a chronic state of overtaxed physiological stress systems resulting from maladaptive environmental design 45.
The Biological Cost of Poor Design
Prolonged exposure to cramped spaces, harsh lighting, and predominantly angular designs triggers physiological cascades analogous to genuine physical threats. Chronic architectural stress is hypothesized to induce amygdala hyperactivity and potentially contribute to long-term hippocampal shrinkage, actively deteriorating memory formation, emotional regulation, and decision-making capacities 45. Conversely, neuro-informed architecture seeks to mitigate this neurological tax by integrating biomorphic forms and optimized spatial geometry 3145. Wearable functional near-infrared spectroscopy (fNIRS) studies assessing college students show that exposure to highly unnatural, high-load artificial environments (like prolonged social media consumption in sterile settings) reduces dorsolateral prefrontal cortex (dlPFC) activation, reflecting acute impairments in working memory and cognitive inhibition 4.
Biomorphic Forms and Contextual Amygdala Activation
Biomorphic design mimics the organic, fluid aesthetics of the natural world through architectural forms 31. Experimental aesthetics frequently investigates the dichotomy between curved and angular architectural contours. In general aesthetic evaluations, humans exhibit a strong innate preference for curved shapes, which are perceived as safer and more pleasant, while sharp angles often activate threat-perception mechanisms in the amygdala 46.
However, neuroimaging has revealed that context heavily modulates this baseline aesthetic preference. In specific functional environments, such as complex hospital layouts where rapid wayfinding is paramount, fMRI data unexpectedly shows greater amygdala activation in response to highly curved designs 46. In these high-stress contexts, excessive curvature induces spatial ambiguity and navigational anxiety, overriding the baseline preference for organic shapes 46. This highlights a crucial principle in neuroarchitecture: aesthetic interventions cannot be universally applied without actively considering the cognitive demands and emotional state of the occupant 46.
Multisensory Integration and Frontopolar Processing
To achieve optimal cognitive outcomes, architectural biophilia must be multisensory. Research indicates that single-variable interventions (e.g., visual biophilia alone, such as a localized plant wall) are substantially less effective at reducing neuroinflammatory markers and enhancing task performance than multi-sensory integrations that combine visual, acoustic (e.g., water sounds), and tactile (e.g., wood grain) elements 41.
The evaluation of complex spatial environments relies heavily on the lateral frontopolar cortex (FPl). While traditional decision-making (e.g., choosing a specific item or task) recruits the ventromedial prefrontal cortex (vmPFC), the FPl is uniquely equipped to process spatial environments 20. The FPl decomposes complex environmental features via parallel processing, evaluating the hierarchy, complexity, and quality of a space before navigational decisions are made 20. By designing environments rich in coherent complexity, fractal scaling, and optimal light penetration, architects reduce the computational friction required by the FPl, thereby preserving cognitive bandwidth for high-order creative and executive tasks 203741.
Emotional Regulation and Clinical Applications
Beyond theoretical models of beauty and architectural preferences, neuroaesthetics provides actionable, empirical insights for psychological health and clinical neurorehabilitation. The capacity to engage deeply with aesthetic stimuli fundamentally relies on, and actively trains, the brain's emotional regulation networks.
Prefrontal Control and Cognitive Reappraisal
Aesthetic experiences often require the viewer to process complex, sometimes negative or challenging, emotions within a safe cognitive frame 3047. This mirrors the psychological process of cognitive reappraisal - the ability to cognitively transform the meaning of an emotional experience to lessen distress 47.
fMRI studies examining emotion regulation reveal a distinct neural signature for cognitive reappraisal. Passive viewing of highly negative or distressing stimuli reliably activates limbic regions, primarily the amygdala and anterior insula 48. However, when individuals actively reappraise the stimulus (as one might when interpreting a dark or challenging artwork), there is a pronounced increase in the activation of the dorsolateral (dlPFC) and ventrolateral prefrontal cortices (vlPFC), alongside the anterior cingulate cortex 474861. This prefrontal activation effectively exerts top-down inhibitory control over the amygdala and medial orbitofrontal cortex, significantly reducing the subjective experience of negative affect without the physiological cost of simply suppressing behavioral expression 47.
The efficacy of this top-down control is highly vulnerable to environmental context. Acute situational stress attenuates prefrontal activation and enhances limbic responses, disrupting the critical functional connectivity between the prefrontal cortex and the amygdala 48. Interventions that utilize aesthetic environments (such as immersive nature or highly coherent architecture) serve to lower baseline allostatic load, thereby preserving the integrity of these prefrontal-limbic communication channels, allowing individuals to regulate emotion more effectively 4248.
Neurorehabilitation and Art Therapy
The anatomical overlap between aesthetic processing networks and cognitive control networks provides a rigorous neurological rationale for clinical art therapy and neurorehabilitation. In patients recovering from stroke or suffering from neurodegenerative diseases (e.g., Alzheimer's disease, Parkinson's disease, or frontotemporal dementia), exposure to preferred aesthetic stimuli - a phenomenon termed the "Michelangelo Effect" - has been shown to stimulate brain plasticity and enhance functional recovery 23.
Art-based interventions do not simply act as passive mood enhancers; they actively reconfigure large-scale brain networks 329. By engaging the Mirror Neuron System through the physical act of artistic creation or the embodied simulation of viewing, and by simultaneously recruiting the Default Mode Network for self-referential processing, art therapy facilitates a neurological shift away from rigid, maladaptive neural states 2729.
Furthermore, during the creative generation stage, aesthetic processing synergistically engages the Salience Network (SN) to monitor novel emotional features, while temporarily suppressing the Executive Control Network (ECN) to allow for free associative thinking 18. Multi-level interpersonal synchronization during shared aesthetic experiences between a patient and therapist amplifies oxytocin release, deepening therapeutic connection and providing a neurobiological scaffold for corrective emotional learning and cognitive rehabilitation 727.