Mechanisms of Human Cooperation
The existence of widespread, large-scale cooperation among genetically unrelated individuals constitutes a significant theoretical puzzle in evolutionary biology, anthropology, and the social sciences. In a strictly Darwinian framework where natural selection optimizes individual fitness, behaviors that incur a personal survival or reproductive cost to bestow a benefit upon a competitor should theoretically be eliminated. However, cooperation acts as a fundamental architect of biological complexity - driving the evolution of genomes, multicellular organisms, and human civilization. The contemporary understanding of human prosociality synthesizes mathematical models from evolutionary game theory, functional neural mapping from social neuroscience, and demographic data from evolutionary anthropology.
Research demonstrates that human cooperation is not an evolutionary anomaly. Instead, it relies on specific spatial and temporal interaction structures, neurobiological reward and threat-detection pathways, and the long-term mechanics of gene-culture coevolution. The following analysis systematically examines the variables that permit cooperative behaviors to emerge, stabilize, and occasionally collapse in human populations.
Evolutionary Game Theory and Interaction Structures
Mathematical models of natural selection inherently favor defection in a well-mixed population where any individual is equally likely to interact with any other. In the standard framework of cooperative dilemmas, such as the Prisoner's Dilemma, a cooperator pays a fitness cost ($c$) to dispense a fitness benefit ($b$) to a recipient. A defector pays no cost and distributes no benefit 1. Because defectors reap the rewards generated by cooperators without bearing the energetic or reproductive costs, their relative fitness is higher. In the absence of an organizing mechanism, selection continuously reduces the abundance of cooperators until they face extinction 123.
Mechanisms of Assortment and Reciprocity
For cooperation to survive, natural selection requires specific interaction structures that bias the distribution of benefits toward other cooperators. Evolutionary biologists have formalized five primary mechanisms that alter population dynamics to allow cooperation to outcompete defection 1456.
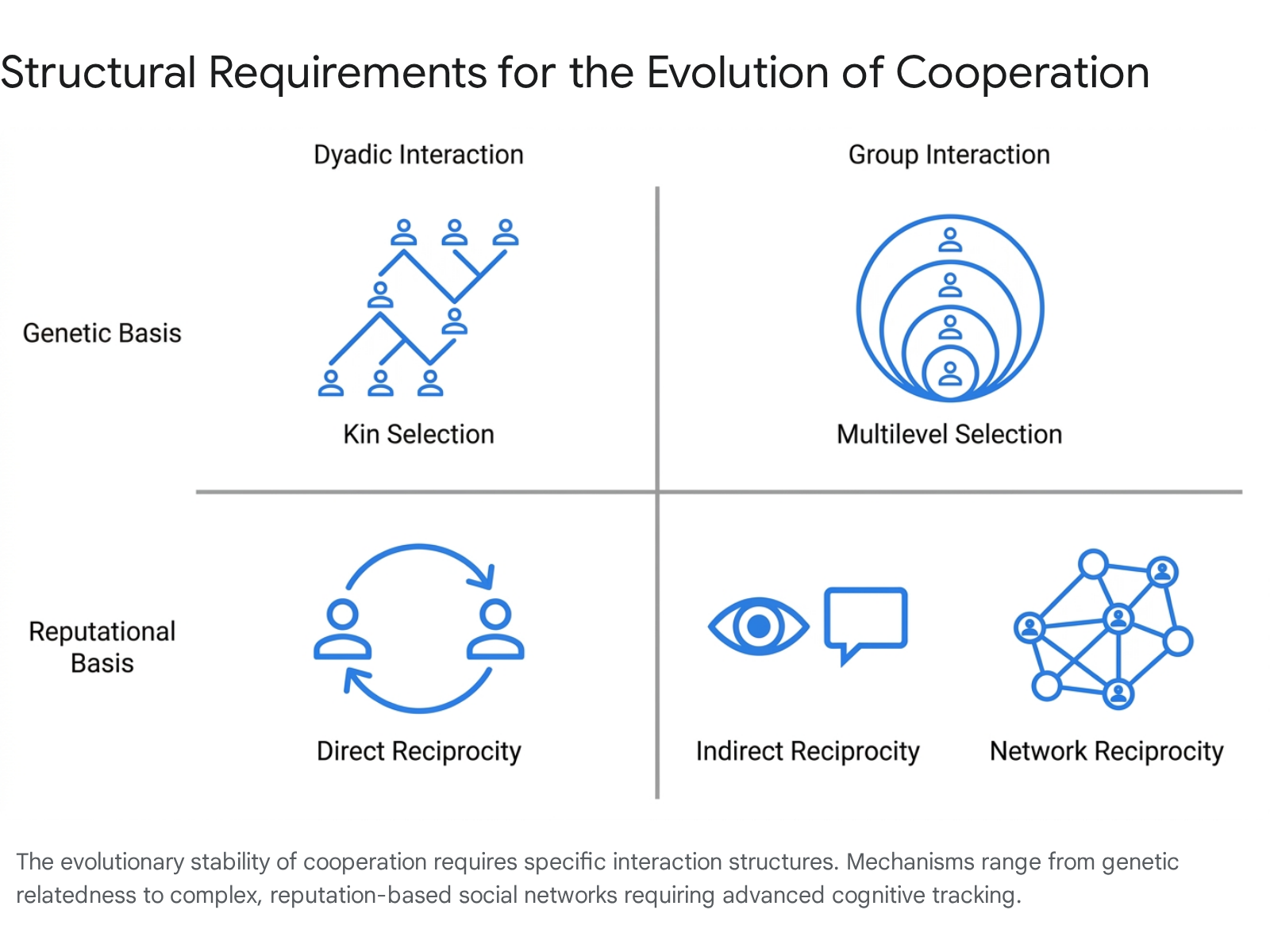
Each mechanism modifies the structure of interactions and yields a specific mathematical threshold that must be surpassed for cooperation to be evolutionarily stable.
| Mechanism | Structural Requirement | Mathematical Threshold | Evolutionary Dynamics |
|---|---|---|---|
| Kin Selection | Genetic relatedness | $r > c/b$ | Cooperation is favored if the coefficient of genetic relatedness ($r$) exceeds the cost-to-benefit ratio, explaining prosociality within biological families 17. |
| Direct Reciprocity | Repeated dyadic encounters | $w > c/b$ | Cooperation evolves if the probability ($w$) of subsequent interactions between the same two individuals is sufficiently high to allow conditional strategies 17. |
| Indirect Reciprocity | Reputation tracking | $q > c/b$ | Cooperation is sustained when the probability ($q$) of knowing a stranger's past reputation exceeds the cost-to-benefit ratio 178. |
| Network Reciprocity | Spatial/network structure | $k < b/c$ | Clustering allows cooperators to form localized support networks. The benefit-to-cost ratio must exceed the average number of neighbors ($k$) 17. |
| Multilevel Selection | Inter-group competition | $1 + n/m < b/c$ | Competition between populations favors groups with cooperators. Evolution relies on maximum group size ($n$) and the total number of competing groups ($m$) 17. |
While kin selection effectively explains altruism among close relatives, it remains insufficient to explain the scale of human cooperation across vast networks of genetically unrelated strangers 1. Direct reciprocity - operationalized by conditional strategies such as "tit-for-tat" or "win-stay, lose-shift" - expands cooperation to unrelated individuals, provided they interact repeatedly and can conditionalize their behavior based on previous encounters 2789.
Humans, however, frequently cooperate in one-shot, anonymous interactions where future encounters are highly unlikely. Evolutionary game theory posits that indirect reciprocity serves as the primary mechanism bridging this gap. Indirect reciprocity relies on the monitoring and broadcasting of reputation: an individual assists another to establish a social standing that will elicit future cooperation from third parties 47. This mechanism requires sophisticated cognitive architecture, specifically social intelligence and the capacity for complex language, to accurately track, negotiate, and communicate the moral standing of numerous network members 589.
Spatial Selection and Dynamic Network Modifications
Static game theory models often assume a fixed population structure, but empirical human interactions occur within highly fluid social networks. Theoretical models and behavioral experiments demonstrate that dynamic network restructuring serves as a powerful exogenous mechanism to stabilize cooperation 10.
When individuals are embedded in social networks and given the agency to update their connections, they consistently engage in strategic link formation and dissolution. In experimental public goods games, if networks are shuffled randomly or remain rigidly fixed, cooperation inevitably decays over time 10. Conversely, when participants can frequently update their network ties, cooperation is maintained at exceptionally high levels. Individuals preferentially sever ties with defectors and forge new ties with cooperators 1011. This endogenous network rewiring creates a structural incentive for prosociality without requiring individuals to alter their internal baseline disposition toward cooperation.
Breaking links functions as a costless alternative to traditional punishment. In classical models, costly punishment - where an individual pays a material fee to inflict a larger penalty on a defector - can force compliance but results in a net loss of resources for the broader group 410. Link dissolution isolates defectors from the collective benefits of the cooperative cluster without draining resources from the cooperators, preventing retaliatory vendettas and enhancing overall network fitness 10.
Multilevel Selection Dynamics
Recent syntheses in evolutionary game theory have extended these models to describe multilevel dynamics utilizing nested birth - death processes and partial differential equations (PDEs) 12. These models track natural selection acting simultaneously within groups (favoring defectors) and among groups (favoring cooperators). Mathematical analysis reveals a threshold selection strength ($\lambda^*$) required for the long-term survival of cooperation, determined by the ratio of the individual incentive to defect against the collective incentive to cooperate 12.
However, mathematical limits indicate the presence of the "shadow of lower-level selection." Because no amount of inter-group competition can entirely erase the local individual advantage a defector gains within a cluster of cooperators, multilevel selection often yields "second-best" solutions rather than absolute social optimums 12.
Social Norms and the Economics of Punishment
The enforcement of cooperation in multi-agent environments relies heavily on social norms and the willingness of third parties to sanction norm violators. Altruistic punishment - the sanctioning of those contributing little to a public good - has historically been proposed as the principal mechanism underpinning the extraordinary extent of human cooperative culture 13. However, empirical investigations across diverse global societies reveal that punishment does not universally promote cooperation; its efficacy is strictly contingent upon prevailing cultural frameworks.
Cross-Cultural Variation in Antisocial Punishment
Research analyzing cross-cultural public goods games has identified the widespread phenomenon of "antisocial punishment" - the costly sanctioning of individuals who contribute the same as, or more than, the punisher 13141516. Rather than targeting free-riders, some individuals weaponize punishment against extreme cooperators to prevent them from establishing dominance or elevating group contribution expectations. Finite mixture models applied to global datasets reveal significant heterogeneity in punisher profiles. Approximately 64.7% of punishers act as standard "Type F" individuals who exclusively target free-riders, while roughly 35.3% manifest as "Type AF" individuals who indiscriminately punish both free-riders and high-contributing cooperators 1416.
The prevalence of Type AF punishers varies dramatically across cultural boundaries and has profound implications for economic efficiency.
| Cultural and Economic Context | Punishment Dynamics | Impact on Public Goods Provision |
|---|---|---|
| High Civic Norms / Strong Rule of Law | Low antisocial punishment; sanctions are reliably directed at free-riders (Type F punishers dominate) 131417. | Initiates a "virtuous circle" where social punishment is accepted, driving populations toward near-total cooperation and maximum payoff 17. |
| Weak Civic Norms / Low Rule of Law | High antisocial punishment; cooperators are frequently sanctioned by low-contributors (Type AF punishers prevalent) 131417. | Triggers a "vicious circle" of retaliation. Antisocial sanctions neutralize the benefits of prosocial punishment, preventing full public goods provision 1617. |
In cultural contexts where antisocial punishment is frequent (such as specific regional cohorts observed in Southern Europe and the Middle East), the introduction of punishment mechanisms actively depresses overall public goods provision 17. The existence of antisocial punishment negatively correlates with survey-based macro-indicators of civic cooperation and gross domestic product (GDP) 1317. This dynamic underscores that introducing punitive institutions will only stabilize cooperation if the underlying social norms already condemn free-riding and respect high contributors 181919.
Neurobiological Substrates of Prosociality
The behavioral algorithms predicted by evolutionary game theory necessitate specific neurobiological hardware to execute them. Evolutionary pressures have instantiated neural architectures and biochemical pathways designed to facilitate, evaluate, and reward cooperative behavior. The neurobiology of cooperation is largely governed by the interplay between the dopaminergic reward system, oxytocin-mediated social boundaries, and prefrontal networks responsible for inequity aversion.
Dopaminergic Modulation of Social Learning
Cooperation frequently requires individuals to forgo immediate, tangible benefits in anticipation of delayed, abstract, or socially mediated rewards. This behavioral flexibility is heavily regulated by dopamine (DA) signaling within the striatum 202122.
Human neuroimaging and positron emission tomography (PET) studies demonstrate that the ventral striatum is critically involved in signaling the value of reward outcomes relative to their expectation, a metric known as the reward prediction error (RPE) 2223. During cooperative exchanges, such as repeated Trust Games, anticipated rewards generated from a participant's belief in their partner's reciprocity activate the bilateral ventral striatum 21.
However, recent multimodal studies combining task-based functional magnetic resonance imaging (fMRI) with $[^{11}\text{C}]\text{Raclopride}$ PET imaging reveal that dopamine release extends beyond mere reward processing. Greater striatal dopamine release is uniquely associated with the adoption of task-optimal learning strategies and the generation of higher-order learning heuristics 2224. Dopaminergic signaling interacts with a broader cortical network - including the anterior cingulate cortex, medial prefrontal cortex, and posterior parietal cortex - primarily during the expectation phase of an interaction rather than the outcome phase 2224. This indicates that the striatal dopamine system actively guides the strategic learning process required to navigate complex social dilemmas. Furthermore, dopamine receptor availability (specifically $D_2/D_3$ receptors) in the fronto-striatal circuit is strongly negatively correlated with stop-signal reaction times, implicating this network in response inhibition - a necessary cognitive function for suppressing selfish impulses 25.
Oxytocin and Parochial Altruism
The hypothalamic neuropeptide oxytocin (OT) is a fundamental mediator of mammalian social attachment and trust. Popular neuroscientific literature frequently characterizes oxytocin as a biological promoter of general benevolence; however, advanced neuroendocrinological studies reframe its role specifically within the context of intergroup competition and "parochial altruism" 262728293031.
Parochial altruism is the behavioral tendency to incur personal costs to benefit one's in-group while simultaneously displaying hostility or withholding resources from competing out-groups 272832. Double-blind, placebo-controlled experiments indicate that intranasal administration of oxytocin triggers a localized "tend and defend" response.
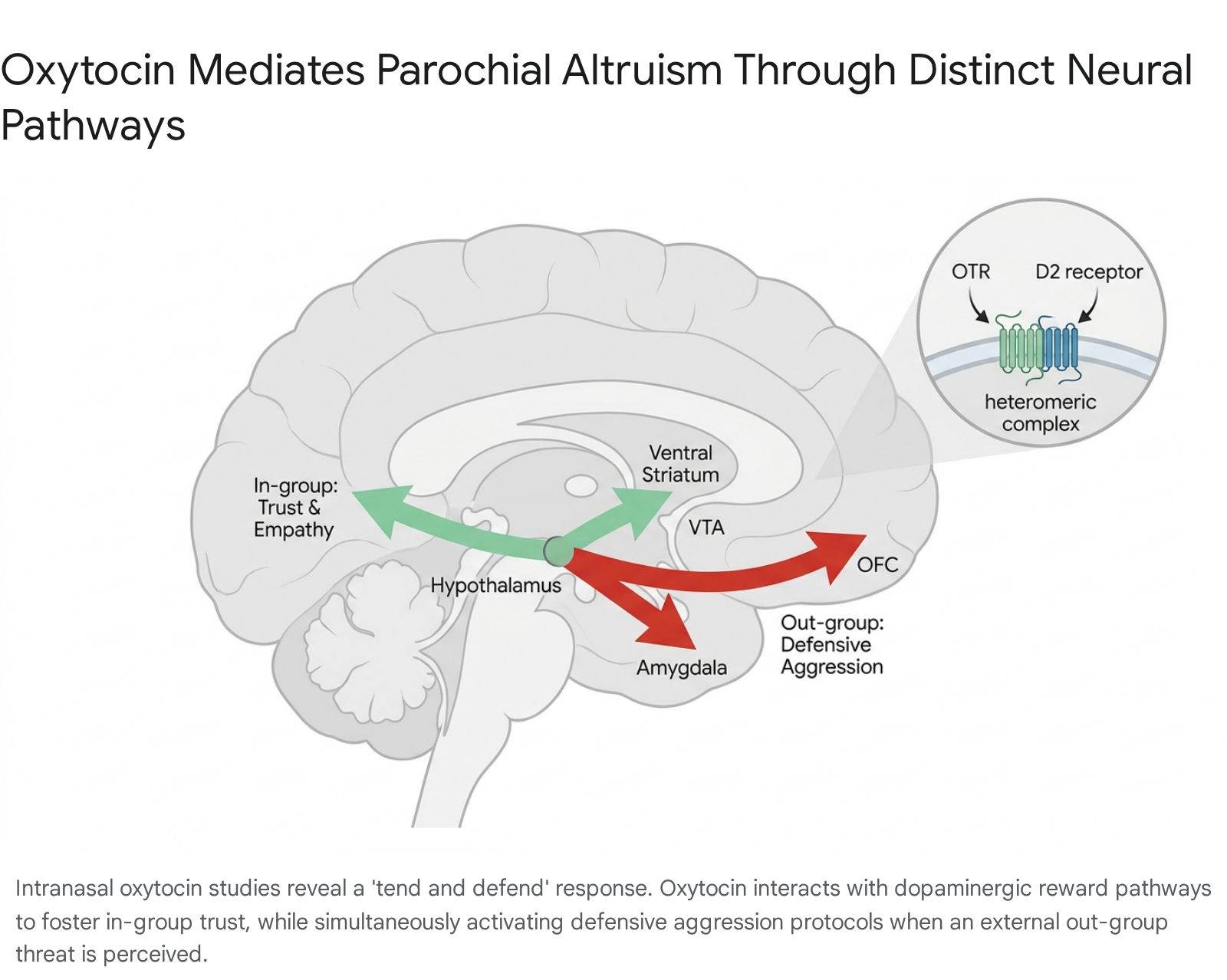
Oxytocin administration significantly upregulates in-group trust, empathy, and conformity 272829. However, when subjects are faced with a threatening out-group, oxytocin selectively promotes defensive aggression - actions aimed at neutralizing the threat to protect the in-group - rather than unprovoked, offensive hate 26272829.
This bounded prosociality highlights the evolutionary origin of oxytocin's function: it evolved to increase the cohesion, efficiency, and survivability of the immediate biological or social unit in a resource-competitive environment 282931. At the molecular level, oxytocin and dopamine systems are deeply interconnected. In the neurons of the nucleus accumbens and the caudate putamen, oxytocin receptors ($OTR$) and dopamine $D_2$ receptors form heteroreceptor complexes 20. Oxytocin-induced $OTR$ activation enhances $D_2$ receptor recognition and signaling, meaning social recognition and bonding are directly integrated into the brain's baseline reward architecture 2033.
Empathy, Inequity Aversion, and Mentalizing Networks
Cooperation is behaviorally constrained by social expectations of fairness and an innate neurological aversion to inequity. Meta-analyses of fMRI studies involving economic exchange games elucidate distinct neural signatures corresponding to cooperation enforcement (fairness and reciprocity) versus cooperation violation (unfairness and betrayal) 343537.
When individuals engage in successful mutual cooperation or enforce fairness, brain regions associated with reward - specifically the orbitofrontal cortex (OFC) and the right caudate nucleus within the striatum - are robustly activated 3436. In contrast, experiencing unreciprocated cooperation or betrayal selectively activates the anterior insula (aINS), a region deeply implicated in the processing of negative emotions, disgust, and physical pain 3435. Quantitative meta-analyses identify the dorsal anterior cingulate cortex (dACC), anterior midcingulate cortex (aMCC), supplementary motor area (SMA), and bilateral anterior insula as forming the core neurocognitive network for empathy 394037.
Neuroimaging further distinguishes between reactions to disadvantageous inequity (receiving less than a partner) and advantageous inequity (receiving more). Disadvantageous inequity triggers the aINS and the amygdala, reflecting an immediate, aversive emotional reaction akin to physical pain 3537. Advantageous inequity, however, engages the dorsomedial prefrontal cortex (DMPFC) and the right dorsolateral prefrontal cortex (rDLPFC) 35. These regions are central to the brain's "mentalizing" network - the system responsible for Theory of Mind and perspective-taking. This neurological divergence suggests that while receiving less than others triggers an immediate emotional rejection, receiving an unfairly high reward triggers complex social calculations. The individual must mentally model the frustration of their partner and calculate the long-term reputational damage of violating fairness norms, which poses a threat to future cooperative benefits 35373937.
Anthropological Observations of Forager-Horticulturalists
While mathematical models and neuroimaging outline the mechanistic substrates of cooperation, empirical data from contemporary subsistence societies provide a window into the demographic and ecological contexts in which human prosociality evolved. Cross-cultural data from populations such as the Tsimane of the Bolivian Amazon, the Hadza of Tanzania, and the Batek of Malaysia illustrate how socioecological pressures dictate the structure and function of cooperative networks.
Caloric Production and Intergenerational Transfers
The Tsimane Health and Life History Project provides extensive biometric and behavioral data on approximately 16,000 forager-horticulturalists living in village clusters along the Maniqui River system 38. The Tsimane possess a high total fertility rate (averaging nine births per woman) and experience a relatively high pathogen burden, mirroring the demographic realities of ancestral human populations prior to the demographic transition 383940.
Tsimane society requires massive, multigenerational cooperative energy transfers to sustain its population. Because children and adolescents consume significantly more calories than they produce, the caloric burden on young parents is severe 38. Anthropological modeling reveals that Tsimane adults do not reach their peak net caloric production efficiency until their 40s 38. Consequently, survival relies on an extended kin-based sharing network. Older adults (grandparents) continue to produce a net surplus of calories and transfer resources to their descendants well into their 70s, subsiding only when physical disability or sensory decline limits agricultural labor 3840.
Social Networks and Labor Divisions
When analyzing collaborative labor networks, Tsimane cooperation is highly structured and compartmentalized. Network analyses of over 1,500 interviews indicate that Tsimane women share horticultural labor almost exclusively with primary kin (55%) and spouses, with fewer than 1% of ties occurring with unrelated individuals 41.
This modular, kin-intensive network structure sharply contrasts with the labor networks of nomadic foragers like the Batek of Malaysia. Among the Batek, daily nomadic foraging yields high variation in labor partnerships, resulting in diffuse cooperative networks heavily inclusive of non-kin 41. The Tsimane reliance on geographically fixed, extended-family agricultural fields incentivizes restricted, kin-directed labor exchanges, supporting the game theoretic concept that spatial structures heavily dictate the breadth of cooperative ties 741.
Childhood Ontogeny and Skill Acquisition
The complex skills required for subsistence cooperation are not innate; they must be culturally transmitted and practiced. Observational scan samples of Tsimane children demonstrate that cooperative divisions of labor are instilled early in ontogeny through unstructured play 4243.
Both general and object-oriented play peak between the ages of 3.5 and 7.5 years 43. During this developmental window, play heavily mirrors the sex-specific division of labor observed in adult Tsimane society. Boys predominantly engage with objects related to hunting and male-associated agricultural labor, whereas girls disproportionately engage with objects associated with food processing and household maintenance 4243. This pattern of object play acts as an adaptive, culturally embedded mechanism, providing a low-risk environment for children to develop the specific biomechanical and cognitive tool-use skills they will rely upon as adults 4243. When acquiring essential survival skills, Tsimane learners rely heavily on vertical transmission from older, same-sex relatives (primarily parents), while utilizing older generations (grandparents) for skills requiring extensive cultural knowledge but lower physical strength, such as storytelling 44.
Market Integration and Changing Partner Preferences
The criteria for selecting cooperative partners among non-kin shift dynamically in response to ecological and cultural changes. Studies of the Hadza hunter-gatherers demonstrate that the valuation of a cooperative partner is sensitive to external institutional exposure 49.
In 2016, Hadza individuals exhibited a strong preference for campmates who were highly skilled hunters, valuing gross caloric return over strict generosity. However, as the population experienced greater exposure to outside institutions, tourism, and market economies, this preference shifted. By 2019, the social preference for generous individuals superseded the preference for pure foraging ability 49. Furthermore, field experiments challenge the assumption that human food sharing is exclusively driven by intrinsic fairness. Among the Hadza, redistribution is frequently characterized by "demand-sharing" - a system where resources are relinquished due to the persistent requests and social pressure of campmates rather than unsolicited altruism 49. In controlled "give-or-take" economic games, the majority of Hadza participants tolerated inequality when it benefited themselves but rejected it when it benefited others 49. This indicates that high levels of observed cooperation in subsistence societies are often maintained by extrinsic social norms and peer pressure rather than innate egalitarian preferences.
| Subsistence Population | Primary Ecological Mode | Cooperative Network Structure | Key Cooperative Dynamic |
|---|---|---|---|
| Tsimane (Bolivia) | Slash-and-burn horticulture, hunting, fishing 3840. | Modular, kin-intensive. Low absolute frequency of non-kin labor sharing 41. | Multi-generational, top-down caloric transfers (grandparents to youth) 3840. |
| Batek (Malaysia) | Nomadic foraging, gathering, hunting 41. | Fluid, highly variable. Frequent daily cooperation with diverse non-kin 41. | Daily pooling of foraging returns within the active, fluid camp 41. |
| Hadza (Tanzania) | Hunting, gathering, increasing market integration 49. | Fluid camp mixing, robust assortment of cooperators clustering together 49. | Extrinsically motivated "demand-sharing" driven by peer pressure 49. |
Gene-Culture Coevolution and Cultural Group Selection
The biological and anthropological data converge on a broader macro-evolutionary framework: human cooperation cannot be entirely explained by genetic evolution acting on individuals. Instead, human prosociality is the product of gene-culture coevolution - a dynamic where cultural innovations rapidly create novel environments that expose the human genome to new selective pressures 5045524647. Well-documented examples of this dynamic include the evolution of lactase persistence in response to dairy farming and the proliferation of the sickle-cell trait in response to agriculture-induced mosquito habitats 5246.
The Norm Psychology Hypothesis
Unlike other species, humans possess a cumulative cultural inheritance system that operates in tandem with genetic inheritance. Theoretical biology suggests that as our ancestors developed complex cultural practices, a specific "norm psychology" evolved 50. This encompasses a suite of psychological adaptations explicitly designed to infer, encode, adhere to, and enforce the shared behavioral standards of a local community 50.
Because individual survival in early human societies became heavily dependent on inclusion within a cooperative group, natural selection favored individuals possessing the cognitive architecture to rapidly absorb local norms and the neurobiological aversion to violating them 3550. This culture-driven genetic adaptation explains the human propensity for prestige-biased learning - preferentially copying the behaviors of successful or highly respected individuals rather than simply relying on individual trial-and-error 4849.
Competition and Equilibrium Selection Between Cultural Groups
Cultural Group Selection (CGS) posits that social learning mechanisms, paired with rewards and punishments, can stabilize diverse behavioral equilibria across different human groups. When groups compete, those possessing norms that foster high internal cooperation and cohesion outcompete and displace less cooperative groups, thereby spreading prosocial norms across the broader population 4850515253.
Because cultural evolution occurs much faster than genetic evolution, and because cultural F-statistics (the measure of between-group variation) are significantly higher than genetic variation among human populations, selection acting on cultural variation is mathematically highly plausible 465455. Over time, this dynamic means that human survival and reproduction depend less on individual genetic traits and more on the health and cultural infrastructure of the society to which they belong 55.
Critiques of Cultural Group Selection and the Selfish Gene
Despite its explanatory power, the specific mechanics of Cultural Group Selection remain fiercely debated. Traditional models of CGS, sometimes termed "maladaptive CGS," assume that humans rely on content-blind social learning strategies (such as strict conformism), which can theoretically stabilize any arbitrary behavior, even those detrimental to the individual 4850. Critics of this view argue that empirical evidence indicates social learning is highly strategic. Individuals actively evaluate their socioecological environment and rely on payoff-biased transmission, abandoning norms if they prove personally disadvantageous 4850.
Furthermore, critics argue that large-scale human cooperation does not exclusively require brutal inter-group competition or random cultural drift. Instead, human cooperation frequently arises endogenously through the conscious negotiation and design of institutions. Rational actors actively construct and optimize rules, monitoring systems, and punishment mechanisms to align self-interest with collective goals 4850.
These debates reflect a broader movement challenging gene-centric paradigms, most famously popularized by Richard Dawkins' "Selfish Gene" metaphor 56645766. Mathematical models analyzing complex systems demonstrate that the effective evolutionary fitness of an allele depends heavily on the distribution of alleles in the population; complete mixing of alleles rarely occurs 56. Consequently, modeling evolution strictly at the level of isolated, competing genes fails to account for the deeply intertwined genetic and cultural networks that define human populations 56576667. Modern evolutionary theory is shifting to acknowledge that through niche construction and institutional design, human culture frequently preempts genetic adaptation, providing rapid solutions to environmental challenges that biology alone could not solve 455557.