Grid Cells in Spatial and Abstract Cognition
The mammalian brain possesses an extraordinary capacity to continuously map its environment, track self-location, and abstract physical dimensions into complex cognitive architectures. The foundational discovery of place cells in the hippocampus established that neural activity could correlate directly with an animal's physical coordinates, providing the basis for a cognitive map 12. Decades later, the discovery of grid cells in the medial entorhinal cortex (MEC) revealed a more sophisticated phenomenon: a universal, sensory-independent spatial metric 23. Rather than firing in a single location, grid cells activate in a regular hexagonal lattice that tessellates the entire navigable environment, providing a geometric coordinate system 34.
Contemporary neurobiological analysis indicates that the grid cell network does not solely process physical space. Expanding beyond physical navigation, this hexagonal coordinate system appears to underpin various domains of abstract cognition, ranging from the mapping of auditory frequencies and social hierarchies to providing structural blueprints for artificial neural networks 567. This report investigates the properties of the grid cell network, the associated mechanistic computational models, its translation to abstract conceptual domains, methodological controversies surrounding human functional magnetic resonance imaging (fMRI) signal detection, and the architectural convergence between biological cognitive maps and artificial Transformer models.
Biological Properties and Network Topography
The geometric regularity of grid cells is internally generated by the cortex and maintained across different environments, regardless of the specific sensory landmarks present 28. This independence from sensory input distinguishes the entorhinal grid system from the hippocampal place system, which relies heavily on environmental cues to establish location-specific firing 9.
Modular Organization and Geometric Scaling
Grid cells are organized topographically along the dorsoventral axis of the MEC 10. Electrophysiological recordings have demonstrated that the spatial scale of grid fields - the distance between the nodes of the hexagonal pattern - increases progressively from the dorsal to the ventral regions of the MEC 1011. This expansion does not occur as a continuous gradient but is clustered into discrete, anatomically overlapping modules 14.
Within any given module, grid cells share the same spatial scale, orientation, and theta-frequency modulation, differing only in their spatial phase (the specific x-y offset of their grid fields) 1412. The spatial phases are uniformly distributed, allowing the population of cells within a single module to collectively tile every location in an environment 23.
Analysis of large-scale neural recordings, sometimes involving up to 186 cells from a single animal, reveals a highly precise mathematical relationship between these modules 14. The scale of the grid spacing increases from one module to the next larger module by a mean ratio of approximately 1.42, which closely aligns with the square root of 2 ($\sqrt{2}$) 11013. Theoretical biology frameworks suggest that a geometric progression of $\sqrt{2}$ represents an optimal strategy for mapping environments at the highest possible spatial resolution while utilizing a minimal number of neurons 1013. Furthermore, while grid cells can undergo spatial remapping in highly distorted environments, the rigid geometric relationship and alignment of axes between cells within the same module are strictly preserved 1213.
Cellular Architectures for Spatial Navigation
The navigation system relies on a diverse array of specialized neurons that function cooperatively across the entorhinal-hippocampal circuit. The MEC and the hippocampus process parallel streams of sensory, directional, and temporal data to generate a coherent allocentric (world-centered) representation of space.
Diversity of Spatial Cell Types
The following table summarizes the primary cell types identified within the spatial navigation network, comparing their localization and principal firing characteristics.
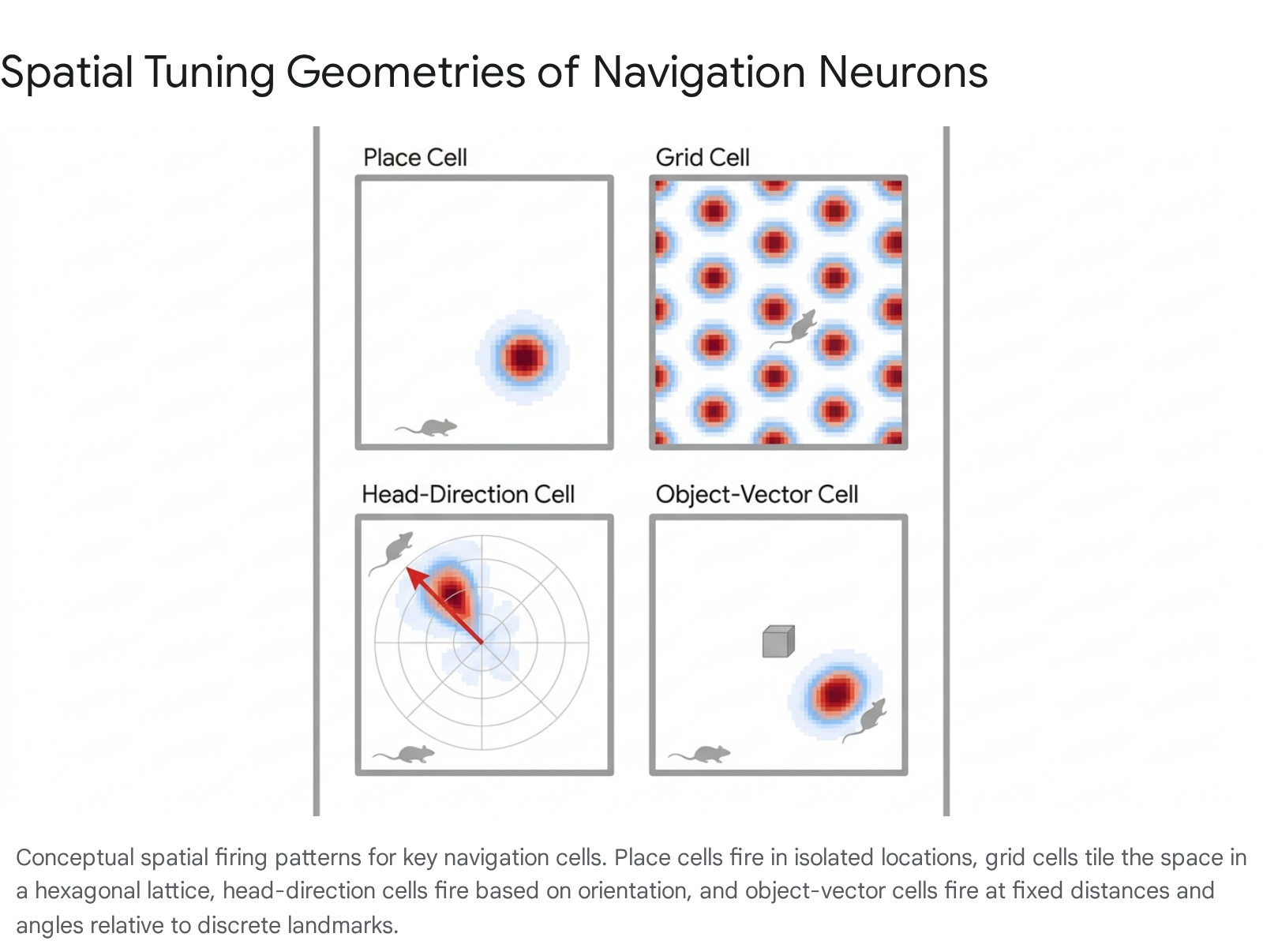
| Cell Type | Primary Brain Region | Firing Characteristic | Reference |
|---|---|---|---|
| Place Cells | Hippocampus (CA1, CA3) | Fire at specific, circumscribed locations within a given environment (place fields). | 11114 |
| Grid Cells | Medial Entorhinal Cortex (MEC) | Fire in multiple locations forming a periodic hexagonal lattice. | 3411 |
| Head-Direction Cells | Presubiculum, MEC | Fire when the animal's head is oriented in a specific azimuth direction, independent of location. | 1115 |
| Border Cells | MEC | Fire predominantly when the animal is near environmental boundaries or edges. | 1115 |
| Conjunctive Cells | MEC, Presubiculum | Fire in a hexagonal grid pattern but only when the animal is facing a specific direction. | 1112 |
| Object-Vector Cells | MEC | Fire at a specific distance and direction relative to a discrete object, regardless of absolute location. | 161718 |
| Predictive Grid Cells | MEC (Layer 3) | Grid fields represent projected future locations, shifting against the direction of travel. | 161920 |
Object-Vector Integration
Recent electrophysiological advances have expanded the known repertoire of MEC neurons beyond pure metric computations. While standard grid cells provide a sensory-independent spatial map, animals must also anchor these coordinates to the external world 121. Object-vector cells in the MEC facilitate this by firing exclusively when the animal is situated at a specific distance and allocentric direction from a discrete environmental object 1617.
Unlike cells in the lateral entorhinal cortex (LEC) that respond to the identity or presence of the object itself, object-vector cells are active in the space surrounding the object 18. These neurons maintain their vector relationships even when the object is moved to a novel location within the environment, establishing a highly flexible mechanism for mapping navigable space between obstacles and landmarks 1822.
Predictive Grid Coding and Rapid Spatial Sweeps
High-resolution temporal recordings have identified a subpopulation of neurons known as "predictive grid cells," located primarily in Layer 3 of the MEC 1920. Rather than encoding an animal's current position, these neurons project future locations by shifting their grid fields against the current direction of travel during goal-directed behavior 1923. These predictive cells discharge synchronously at the trough phases (approximately 180°) of hippocampal CA1 theta oscillations 1920. By organizing sequences of trajectories from present to future positions across each theta cycle, the MEC provides a predictive map essential for forward planning and path simulation 1921.
The dynamics of this predictive capacity extend to rapid, rhythmic scanning functions. Advanced neural probing demonstrates that grid cells coordinate to perform highly regular "sweeps" into the space ahead of the navigating animal 22. These populations alternate between tracking real-time position and scanning up to 30 degrees to the left and right of the current trajectory 22. These forward sweeps occur at approximately ten times per second (a duration of 125 milliseconds per cycle, aligning directly with the theta wave) and follow a geometric "herringbone pattern" 22. Computational modeling indicates this pattern is mathematically optimal for probing future trajectories without generating overlapping representations in adjacent map sectors 22.
Mechanistic Models of Grid Pattern Generation
The emergence of perfectly hexagonal firing patterns from biological neural networks has driven extensive theoretical modeling. Three primary mechanistic classes dominate the computational neuroscience literature: Continuous Attractor Networks, Oscillatory Interference, and Self-Organizing Maps.
Continuous Attractor Networks (CAN)
Continuous Attractor Network (CAN) models posit that grid-like activity arises from the collective, recurrent behavior of a tightly interconnected neural network 2324. In these models, the state of the network is restricted to a low-dimensional continuous manifold formed by local excitatory and inhibitory synaptic connections 2324. The network translates velocity inputs into movements of an "activity bump" across this manifold, effectively performing biological path integration 2324.
Depending on the specific connectivity matrix, the manifold can be conceptualized as a torus, where the activity wraps around the boundaries to produce an infinitely repeating periodic spatial pattern 2324. CAN models are highly robust to intrinsic phase noise and can accurately integrate velocity inputs over substantial distances 23. Furthermore, in vivo whole-cell patch-clamp recordings from moving rodents demonstrate slow depolarizing membrane potential ramps that are highly characteristic of attractor dynamics, validating the biological plausibility of CAN models 25.
Oscillatory Interference (OI) Models
Conversely, Oscillatory Interference (OI) models suggest that the grid pattern is generated independently within single cells rather than relying on a highly recurrent network architecture 232627. OI models depend on velocity-controlled oscillators (VCOs) located in the medial septum or within the dendrites of the grid cells themselves 2627. The frequencies of these oscillators vary based on the animal's movement speed and direction 26.
The grid pattern emerges intrinsically from the constructive and destructive interference between these multiple somatic or dendritic oscillations 2627. While OI models elegantly account for the theta-phase precession consistently observed in grid cells, they face significant physical limitations. They are highly sensitive to phase noise; minor biological desynchronizations rapidly degrade the spatial stability of the grid, necessitating continuous, highly precise phase-reset mechanisms to maintain the lattice structure 2326.
Self-Organizing Maps (SOM)
An alternative computational approach applies the principles of Self-Organizing Maps (SOMs), initially developed as unsupervised machine learning techniques for dimensionality reduction and clustering 313233. SOM models demonstrate how entorhinal cells can develop grid properties developmentally through competitive learning, without relying on pre-configured continuous attractors or perfectly tuned oscillators 2829.
In a biological SOM model, inputs representing integrated linear velocity (often modeled as directionally-selective stripe cells) map onto a discrete grid of cortical neurons 2829. As the animal navigates, the network utilizes competitive learning to identify a "Best Matching Unit" (BMU) for a given state 3236. The BMU and its spatial neighbors adjust their synaptic weights to align more closely with the input distribution 3236. Over repeated exposures, this topology-preserving learning process naturally categorizes the spatial environment into multiple scales. The SOM framework effectively explains how anatomically overlapping modules with distinct spacings and orientations can self-organize along the dorsoventral axis of the MEC based purely on the temporal integration rates of the neurons 2829.
Abstraction of Physical Space into Cognitive Maps
The rigid relational structure of grid cells led to the hypothesis that the entorhinal-hippocampal system provides a universal, domain-general framework for organizing continuous variables 5143031. This implies that the algorithms evolved for physical path integration are recycled by the brain to constitute a "cognitive map" of abstract, non-spatial dimensions 1431.
Mapping Sensory and Temporal Dimensions
Empirical data reveals that grid-like coding mechanisms readily map purely sensory dimensions. When monkeys and humans visually explore static environments while their heads are immobilized, entorhinal neurons and functional imaging signatures demonstrate a six-fold rotational symmetry mapping visual space, utilizing identical path integration mechanisms to physical locomotion 3032.
In the auditory domain, recordings from rodent entorhinal and hippocampal neurons during tasks where animals use a joystick to manipulate sound frequency reveal that spatial cells (including both grid and border cells) form discrete firing fields tuned to specific sound frequencies 530. Because sound frequency operates on a continuous, linear dimension, the neural mechanisms tracking it overlap almost entirely with the spatial cell populations, suggesting a domain-general mechanism for encoding continuous task variables 5.
Mental time travel and temporal processing invoke similar geometric architectures. When human subjects are tasked with mentally simulating trajectories across a conjugated two-dimensional "age-day" space (manipulating conceptual combinations of age and time-of-day), fMRI readings detect hexagonally symmetric, grid-like representations in the entorhinal cortex actively supporting the cognitive trajectory 40.
Navigation of Social Hierarchies
Abstract mapping extends into highly complex relational logic, such as the navigation of discrete social hierarchies. In heavily controlled behavioral paradigms, participants learned the social ranks of individuals across two independent, abstract dimensions: competence and popularity 313334. Crucially, participants never viewed a complete map of these entities; they only learned pairwise relationships on single dimensions via feedback-based training 34.
When subsequently asked to identify the best business partner by combining these dimensions (evaluating the maximum "growth potential" via a calculated vector), participants inferred novel, direct trajectories through the unseen 2D social space 734. Neuroimaging during this decision-making process revealed classic hexadirectional grid-like modulation in both the entorhinal cortex and the medial prefrontal cortex (mPFC), strictly aligned with an internally consistent grid orientation 734. These findings indicate that the human brain constructs a unitary 2D cognitive map of abstract social relationships, deploying the ancient grid cell network to calculate optimal vectors and inferences between abstract social entities 734.
Methodological Controversies in Human fMRI Detection
The extension of grid cell analysis to humans relies heavily on functional magnetic resonance imaging (fMRI) 35. Because fMRI lacks the spatial resolution to measure individual neurons, researchers rely on a macroscopic proxy known as the hexadirectional signal, or the Grid-Cell-Like Representation (GCLR) 353637. However, the validity and mathematical foundation of this proxy have recently become the subject of intense scientific debate.
Assumptions of Macroscopic Grid Signal Detection
The fMRI approach is predicated on the observation that the angular orientation of grid axes is shared among neighboring cells within an MEC module 1237. Due to this coherent orientation, researchers posited that a subject running in virtual reality aligned with the internal grid axes would produce systematic variations in population-level activity compared to running misaligned with the axes 1237.
Historically, this macroscopic variation was attributed to two possible biological mechanisms 3738. The first hypothesis posits directional modulation via conjunctive grid-by-head-direction cells, where preferred firing directions naturally align with the grid axes, boosting the signal at 60-degree intervals 1237. The second hypothesis suggests repetition suppression: moving along aligned axes triggers repeated traversals of dense grid nodes, causing rapid differential adaptation and thereby lowering the localized BOLD signal at specific angles 333747. By modeling a 60-degree (6-fold) sinusoidal modulation of the BOLD signal against the participant's movement trajectory, researchers have widely claimed to detect population-level grid activity in human subjects 1237.
Mathematical Critiques of Mean Signal Analysis
Extensive theoretical analyses and large-scale rodent dataset evaluations published between 2024 and 2026 have critically challenged the statistical assumptions underlying fMRI hexadirectional analysis 353940. The core mathematical critique demonstrates that, from a first-principles perspective, the mean firing rate of a spatially tuned neural population is strictly not modulated by the direction of the trajectory 35. If an environment is sliced into parallel trajectories at any angle, the average activity across those slices remains constant because the total spatial activity across the environment is conserved 35.
Instead, the grid cell architecture predicts a dominant sixfold modulation of cell-level variance, not the population-level mean 3539.
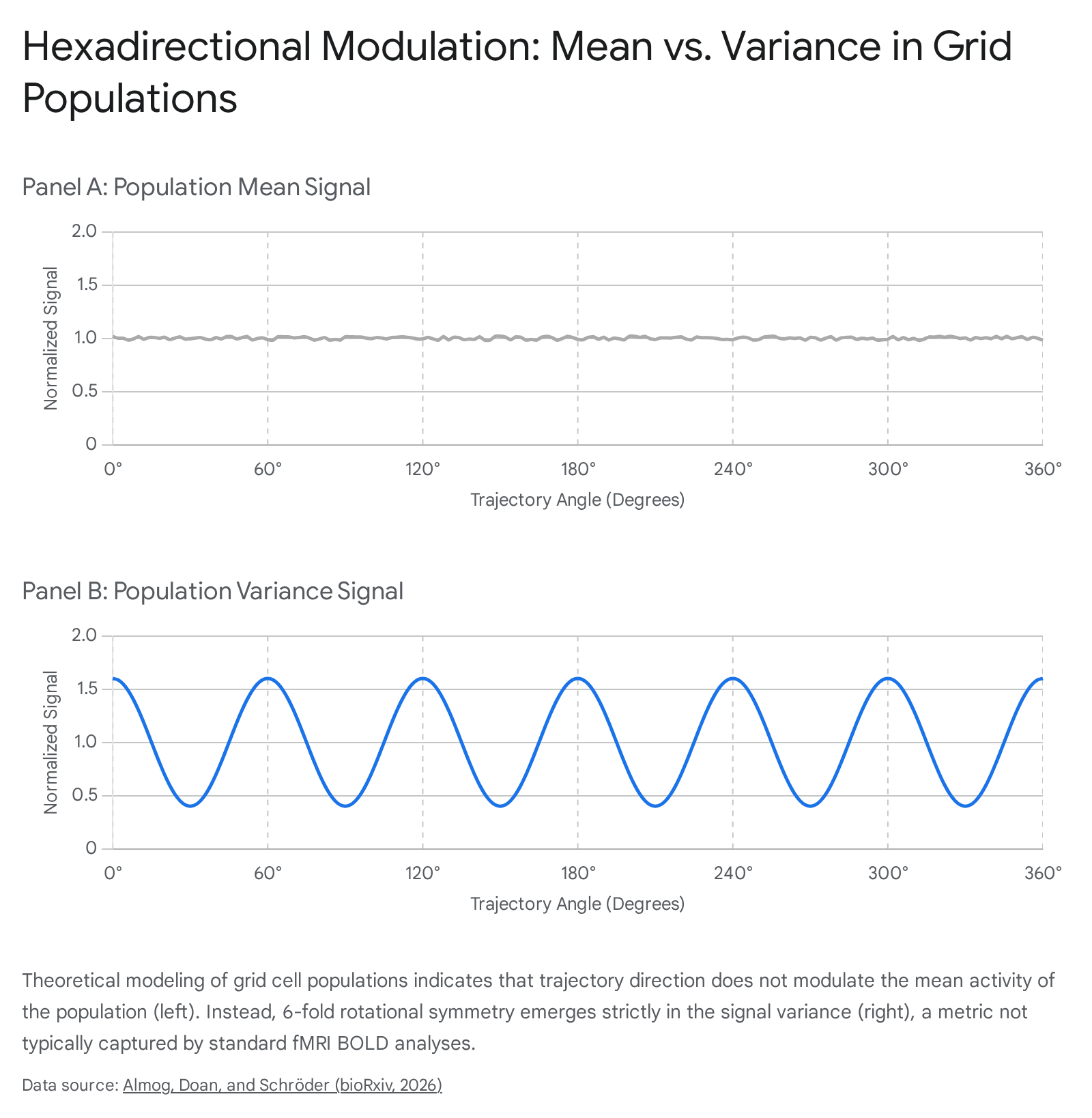
Because the spatial phases of grid cells within a module are distributed uniformly to tile the space, the population-level contrast smooths out, virtually eliminating macroscopic variance 35. Consequently, the standard hexadirectional mean approach used in fMRI studies is largely insensitive to grid firing per se 39.
These theoretical critiques establish that a hexadirectional signal could only emerge in the mean BOLD response if the integrated neuronal firing undergoes a severe second-order nonlinear (superlinear) transformation 35. Even under optimal, constrained conditions, the expected effect size of this mean modulation is remarkably weak, yielding approximately a 2% modulation 35. Furthermore, standard fMRI analyses frequently suffer from harmonic distortion and mis-specified null distributions; traditional sinusoidal resonance tests identify false-positive 6-fold symmetries when evaluating non-sinusoidal, singular directional peaks 35. Concordant with these theoretical critiques, several recent high-powered fMRI studies attempting to utilize grid signals as early biomarkers for Alzheimer's disease have failed to replicate the detection of grid-like representations during passive navigation, highlighting the extreme fragility of the hexadirectional proxy 374142.
Convergence with Artificial Intelligence and Transformers
As computational neuroscience abstracts the grid cell from a physical navigator to an algorithmic processor, striking architectural parallels have emerged between biological spatial circuitry and the networks powering advanced artificial intelligence, particularly Transformer networks and Large Language Models (LLMs).
The Tolman-Eichenbaum Machine (TEM)
The Tolman-Eichenbaum Machine (TEM) represents a unifying theoretical framework detailing how the hippocampal-entorhinal system solves both spatial and relational memory tasks through generalization 43444555. TEM proposes that the MEC operates as a structural basis, calculating the abstract rules and transition matrices of a space, while the hippocampus binds this universal structure to specific sensory representations 4344.
Crucially, the computational mechanisms underpinning TEM demonstrate a direct mathematical equivalence with Transformer neural networks 6. The associative memory component of TEM functions analogously to the self-attention mechanism in Transformers, retrieving memories weighted by their similarity to a query state 6. Furthermore, when a standard artificial Transformer is modified to incorporate recurrent positional encodings instead of static embeddings, the network spontaneously learns to generate internal representations identical to biological entorhinal grid cells and hippocampal place cells to solve simulated spatial tasks 6. This isomorphism suggests that biological grid cells and artificial self-attention layers execute fundamentally similar algorithms: learning the factorized, structural relationships of sequential data streams 643.
Grid-Inspired AI and the Geometry of Thought
Insights from grid cell biology are actively informing next-generation AI design. Modern Transformer architectures excel at sequencing language but natively lack an understanding of multi-dimensional spatial topologies 4657. To resolve this, researchers introduced Grid-cell inspired Positional Encoding (GridPE) techniques 4657. Relying on the premise that biological grid cells represent Euclidean space through a summation of multi-dimensional Fourier basis functions, GridPE applies Random Fourier Features to mimic the hexagonal periodicity of the MEC 4657. Integrating positive spatial frequencies corresponding to multiple biological scales allows Vision Transformers to encode locations in arbitrarily high-dimensional spaces with robust translational invariance 4657.
Similarly, neuro-symbolic computing frameworks utilizing Grid Cell Vector Symbolic Architectures (GC-VSA) deploy structured 3D neuronal modules that explicitly replicate the discrete scales and phases of biological grids to execute complex hierarchical logic and symbolic reasoning tasks 58.
These advances coincide with the emerging "geometry of thought" framework, which evaluates the manifold topology of LLMs during reasoning tasks 476048. Analysis of LLM hidden states reveals that during complex inference, logical structures undergo geometric phase transitions. For instance, during rigorous legal or mathematical computations, the representational dimensionality of the neural activations collapses and forms highly structured, discrete lattices - a process termed "crystallization" 4748. This phenomenon mirrors the brain's reliance on low-dimensional, highly regular geometric manifolds (like the grid cell torus) to enforce coherent cognitive pathways, suggesting that the geometric structuring of information may represent a universal prerequisite for complex reasoning 4748.
Conclusion
The characterization of the grid cell network has precipitated a paradigm shift in cognitive neuroscience, expanding the definition of spatial navigation to encompass the generalized mapping of relationships. From the strict geometric scaling of modules in the medial entorhinal cortex to the predictive sweeps projecting future states, the system provides a robust, sensory-independent coordinate framework. While methodological limitations challenge the interpretation of macroscopic human fMRI data, the underlying neurobiology confirms that the brain processes auditory frequencies, temporal sequences, and discrete social hierarchies using the exact topological algorithms deployed for physical path integration. Ultimately, the mathematical convergence between entorhinal continuous attractor networks and the self-attention mechanisms of Transformer architectures implies that the geometric structuring of information - the hexadirectional parsing of relationships - operates as a universal algorithm for generalized intelligence across both biological and artificial systems.