Evolutionary and cultural foundations of human fairness
The human capacity and deep-seated preference for fairness represent a profound evolutionary and psychological phenomenon. Unlike classical economic models of rational self-interest - which predict that individuals will strictly maximize their own material payoffs - empirical observations across global societies consistently demonstrate a willingness to incur personal costs to uphold equitable norms and punish transgressors 12. The origins, cognitive mechanisms, and cultural expressions of this behavior involve a complex interplay of evolutionary biology, developmental psychology, neurobiology, and sociocultural conditioning. Research over recent decades has synthesized these domains to clarify how fairness evolved as a biological imperative for mutualistic cooperation, how it is hardwired into distinct neural networks, and how it is ultimately calibrated by local cultural paradigms of distributive justice.
Evolutionary Drivers of Distributive Justice
The evolutionary origins of fairness have historically been debated through the lenses of kin selection and reciprocal altruism. Kin selection explains cooperation among genetic relatives, while reciprocal altruism describes the evolution of a disposition to engage in cooperative behavior at a temporary cost, provided the cost is eventually repaid 23. While reciprocity explains ongoing dyadic relationships, it struggles to account for the impartial, systemic nature of human fairness, including the enforcement of fairness norms by third-party observers and the moral outrage triggered by unequal distributions among strangers 34.
The Partner Choice Theory Framework
To address the limitations of reciprocal altruism and partner-control models, contemporary evolutionary biology increasingly emphasizes "partner choice theory" 265. According to this model, human cooperation evolved in a biological market where individuals constantly competed to be chosen and recruited for mutually beneficial endeavors 656. In an environment characterized by fluid social structures and outside options, individuals exercised strategic vigilance, evaluating the competence, reliability, and cooperative intent of potential partners before committing to joint ventures 69.
In a competitive biological market, an individual who consistently demanded more than their equitable share of a cooperative surplus would be shunned in favor of more egalitarian partners 678. Conversely, an individual who passively accepted less than their fair share would be systematically exploited, resulting in a severe fitness disadvantage. Consequently, the optimal evolutionary stable strategy is strict impartiality: distributing the costs and benefits of collective action strictly in proportion to each individual's contribution or their available outside opportunities 678. Under partner choice theory, the human sense of fairness is not a byproduct of irrational altruism, but an adaptive psychological mechanism designed to secure long-term reproductive and survival benefits by maintaining a reputation as a highly desirable, non-exploitative cooperative partner 29.
The drive to maintain this reputation shapes the heuristics used to evaluate social interactions. Individuals evolved to project genuine cooperative intent, as superficial or highly calculated generosity can be detected by strategically vigilant peers 69. This dynamic creates an evolutionary pressure for the internalization of moral norms, rendering fairness an intrinsic motivation rather than a purely calculated response to the immediate threat of punishment 27.
Reputational Management and the Perception of Charitability
The partner choice framework also explains the phenomenon of strategic generosity and the psychological aversion to "hyper-fairness" or overt displays of altruism that appear calculated. Reputational management is deeply embedded in human social cognition; individuals value the perceived motivations behind fair behavior as much as the behavior itself 910. If prosocial behavior is viewed as a mere transaction to secure future benefits, the actor is judged as less morally reliable 10.
Recent experiments mapping attributions of charitability onto Maimonides' Ladder of Charity reveal the nuanced ways humans evaluate cooperative partners. These studies demonstrate that anonymous, double-blind giving is consistently perceived as more genuinely charitable than public giving 10. When a donor reveals their identity, observers frequently ascribe ulterior reputational motives or a desire to establish a superior social hierarchy, leading to a decrease in the donor's perceived charitability 10. In experimental settings, even a hundredfold increase in the size of a donation cannot fully compensate for the social disapproval elicited when a donor forcefully reveals their identity to a beneficiary 10. Furthermore, varying states of mutual knowledge - such as shared knowledge versus common knowledge - subtly alter how fairness and generosity are evaluated, confirming that humans constantly monitor the reputational signals of potential cooperative partners 10.
The Phylogeny of Fairness in Non-Human Animals
To determine whether the human preference for fairness is a unique evolutionary adaptation or a homologous trait shared with other species, researchers have extensively studied non-human animals. Early experiments, notably the 2003 studies on capuchin monkeys, suggested that non-human primates exhibit "inequity aversion" by actively protesting and rejecting a lesser reward (e.g., a slice of cucumber) when a partner receives a highly preferred reward (e.g., a grape) for completing the same task 11121314.
These initial findings led to the hypothesis that disadvantageous inequity aversion - the primary reaction to receiving less than a partner - was widespread among species that cooperate outside of kinship bonds, including chimpanzees, bonobos, and certain macaque species 1415. However, the interpretation of these behaviors as evidence of a biological precursor to the human sense of distributive justice has been subjected to rigorous methodological scrutiny and extensive re-evaluation.
| Primate Species | Level of Social Cooperation | Documented Response in Inequity Paradigms | Theoretical Interpretation |
|---|---|---|---|
| Chimpanzee (Pan troglodytes) | High (alliances, non-kin cooperation) | Negative reaction to unequal rewards; high refusal rates when disadvantaged. | Initially viewed as inequity aversion; increasingly interpreted as a contrast effect/disappointment 111516. |
| Bonobo (Pan paniscus) | High (strong social tolerance) | Ambiguous; elevated refusal rates in some controlled studies when receiving lesser rewards. | Possible sensitivity to unequal outcomes, though advantageous inequity aversion remains undocumented 1315. |
| Capuchin Monkey (Cebus apella) | High (coalitions, food sharing) | Pronounced agitation and refusal of low-value rewards when a partner receives high-value rewards. | Heavily debated; recent large-scale meta-analyses attribute behavior to reward salience and individual expectation rather than social comparison 111216. |
| Orangutan (Pongo) | Low (solitary, minimal non-kin cooperation) | Minimal to no reaction to unequal reward distributions. | Lack of cooperative market pressures results in no evolutionary need for reward comparison 15. |
| Squirrel Monkey (Saimiri) | Low (sympatric with capuchins, but less cooperative) | Reactions driven by prior personal offers rather than partner outcomes. | Behavior governed entirely by individual contrast effects rather than social inequity 15. |
A comprehensive 2024 meta-analysis encompassing 23 studies, 18 species, and over 60,000 observations of accept/reject paradigms found no conclusive empirical evidence for true inequity aversion in non-human animals 1617. The rejection of lower-value rewards by primates in unequal settings is increasingly attributed to the "disappointment hypothesis." In these scenarios, the mere visual presence of a high-value reward creates an expectation; when the animal is subsequently given a low-value reward, it rejects the food out of individual frustration rather than a social comparison regarding the partner's outcome 111216.
This divergence emphasizes that while non-human primates engage in cooperative behavior, the specific psychological architecture required to compute equitable distributions, anticipate a partner's resentment, and voluntarily sacrifice resources to equalize outcomes - foundational to human advantageous inequity aversion - appears to be a uniquely human adaptation shaped by the intense pressures of human biological markets 714.
Ontogenetic Development of Fairness Norms
If human fairness is an embedded cognitive architecture shaped by natural selection, its foundational elements should emerge early in human development, prior to extensive cultural socialization. Developmental psychology provides compelling evidence that a rudimentary understanding of distributive justice is present in pre-verbal infants, though the behavioral execution of fairness undergoes a prolonged maturation process lasting into late childhood.
Emergence of Fairness Expectations in Infancy
Research utilizing violation-of-expectation paradigms demonstrates that infants possess expectations regarding resource distribution and social parity long before they can articulate moral rules 181920. When presented with visual scenarios of resources being distributed by a third party, infants as young as 12 to 15 months look significantly longer at unequal distributions (e.g., a 3:1 allocation ratio) than equal distributions (e.g., a 2:2 allocation ratio) 1920. This heightened visual attention indicates surprise and a violation of their baseline expectations of egalitarian fairness 19.
Furthermore, sociomoral evaluations emerge before the end of the first year; infants demonstrate a clear preference to interact with agents who previously distributed resources equally rather than those who distributed them unfairly 1819. The onset of this fairness expectation is closely linked to the infant's own prosocial behavior. Individual differences in a 9-to-15-month-old infant's willingness to naturally share preferred toys reliably predict their sensitivity to fairness norm violations 1920. Infants who readily share highly valued resources show a much sharper detection of inequitable distributions than those who do not yet share, suggesting that the experiential onset of prosocial behavior helps scaffold the cognitive detection of distributive justice 1920.
The Concept-Action Gap in Early Childhood
Despite early conceptual awareness, young children exhibit a well-documented "concept-action gap" regarding fairness 2122. By age three, children can explicitly articulate the principle of equality and correctly identify proportional merit - recognizing, for instance, that an individual who exerted more effort or completed more work on a task is entitled to a larger reward when judging third-party scenarios 2123. However, when their own resources are at stake, preschool-aged children frequently behave selfishly, prioritizing personal material advantage over the fairness principles they explicitly endorse 2122.
This behavioral gap highlights the dual nature of fairness enactment: it requires both cognitive evaluation of the rule and the maturation of executive control networks to override immediate self-interest 21. Children in this developmental stage are highly sensitive to relative advantage; the drive to maintain superiority over peers often supersedes their conceptual knowledge of equity 2122. It is not until executive functioning improves and the reputational costs of selfishness are internalized through peer interaction that children begin to consistently align their actions with their stated fairness beliefs.
Maturation of Inequity Aversion Across Cultures
The alignment of fairness knowledge and actual fair behavior typically solidifies in middle childhood, though the exact trajectory is heavily moderated by cultural context. Cross-cultural developmental studies highlight that "disadvantageous inequity aversion" - the rejection of scenarios where a peer receives a larger reward - emerges universally across diverse global populations between the ages of four and ten 24. This universal onset aligns with the evolutionary mandate to prevent exploitation and signal an unwillingness to accept subordinate status in a biological market 1424.
Conversely, "advantageous inequity aversion" - the willingness to reject a scenario where the child themselves receives a disproportionately larger reward than a peer - emerges much later, typically around ages nine to ten 1424. Crucially, this second-order aversion is not a human universal. In global comparative studies, advantageous inequity aversion reliably emerges only in a subset of predominantly Western societies (such as the United States and Canada) and select other populations (such as Uganda) 24. This indicates that while rejecting exploitation is an innate developmental milestone, the active sacrifice of a personal material advantage to maintain strict egalitarianism is dependent on specific cultural socialization and the prevailing societal emphasis on formal equality 24.
Neurobiology of Fairness and Norm Enforcement
Advances in functional magnetic resonance imaging (fMRI), coordinate-based meta-analyses, and intracerebral electroencephalography (iEEG) have elucidated the specific neural architectures that calculate fairness, evaluate social norms, and execute complex moral decisions. Current neurobiological models conceptualize fairness-related decision-making as the interaction between a reflexive, intuitive neural network (System 1) and a reflective, deliberate executive network (System 2) 25.
The dual-system architecture recruits the anterior insula (aINS) for the intuitive detection of norm violations, the ventromedial prefrontal cortex (vmPFC) for subjective valuation, and the dorsolateral prefrontal cortex (dlPFC) for executive control. Additionally, regions such as the temporoparietal junction (TPJ) and the anterior cingulate cortex (ACC) play essential roles in mentalizing and conflict monitoring, respectively 425262728.
Processing Reward and Punishment Prediction Errors
The Ultimatum Game (UG) - in which a proposer offers a split of resources and a responder can accept or reject the offer, destroying the resources for both parties - serves as the standard neuroeconomic paradigm for investigating fairness 129. Rejections of unequal, financially beneficial offers in the UG represent "costly norm enforcement" or negative reciprocity 42530.
The initial evaluation of a fairness norm violation is processed by the brain's intuitive system, which relies heavily on the anterior insula (AI) and the amygdala 42531. The anterior insula is consistently activated when participants receive unfair offers; the magnitude of AI activation scales directly with the degree of unfairness and serves as a strong predictor of whether the offer will be rejected 42631. The AI acts as a primary hub for the brain's salience network (SN), coding for "punishment prediction errors" (PPE). It represents the negative emotional arousal, subjective frustration, and aversive internal states triggered by social inequity 27313233.
Conversely, the subjective value of fair offers and the inherently rewarding nature of equitable social interactions are processed by the ventromedial prefrontal cortex (vmPFC) and the ventral striatum 42526. The vmPFC is functionally specialized for tracking reward prediction error (RPE) signals and encodes the positive hedonic response associated with adherence to social norms and equality 427. Intracranial recordings reveal that the rapid encoding of these prediction errors relies on distinct, localized interactions: RPEs are heavily trafficked between the lateral orbitofrontal cortex and the vmPFC, whereas PPEs drive interactions between the anterior insula and the prefrontal cortex 27.
Executive Control and the Resolution of Conflicting Motives
Translating these conflicting signals - the pure financial self-interest of accepting money versus the emotional drive to punish a transgressor - into an actionable decision requires the executive control network (ECN). This network is primarily anchored in the dorsolateral prefrontal cortex (dlPFC) and the anterior cingulate cortex (ACC) 2526323334. The dlPFC integrates the competing global learning signals emanating from the AI and the vmPFC. It exercises top-down cognitive control to resolve the conflict, either by suppressing the emotional urge to punish (allowing the acceptance of an unfair but financially beneficial offer) or by overriding pure economic self-interest to enforce the fairness norm via rejection 252734.
Fairness decisions in real-world environments rarely occur in a vacuum; they frequently require the brain to adjudicate between multiple, sometimes mutually exclusive, prosocial motives. Recent neuroimaging studies have modeled complex scenarios where inequality aversion directly conflicts with harm aversion (the desire not to reduce others' absolute payoffs) and rank-reversal aversion (the reluctance to overturn an existing social hierarchy) 28.
Under these multidimensional conditions, the brain does not rely on a monolithic "fairness module." Instead, distinct motives compete for representation. The dorsomedial prefrontal cortex (DMPFC), an area associated with mentalizing and conflict resolution, processes harm-related information and communicates directly with the striatum 28. The striatum acts as a crucial neural hub, receiving equality-seeking signals, harm-aversion signals from the DMPFC, and contextual environmental data, integrating them into a final behavioral output 28. If the functional connectivity between the DMPFC and the striatum is robust, the individual is more likely to prioritize harm aversion over strict equality, resulting in less egalitarian redistribution decisions. This complex neural choreography explains why human fairness is highly sensitive to context, mitigating the strict pursuit of equality when it violates other established moral parameters 28.
Large-Scale Brain Networks and the Effects of Aging
The orchestration of fairness relies on dynamic communication between large-scale functional networks, primarily the default mode network (DMN), the executive control network (ECN), and the salience network (SN) 323335. The interaction between emotional processing (SN) and cognitive regulation (ECN) is underpinned by synchronized neural oscillations, particularly in the alpha and gamma bands, which facilitate rapid shifts in attention and conflict resolution during social tasks 3233.
Lifespan development and aging significantly alter the internal organization of these networks and their connectivity with task-relevant brain regions 35. While older adults generally maintain the capacity for norm enforcement and display similar behavioral sensitivity to fairness norm violations as younger adults, neuroimaging reveals that the neural strategies employed shift with age. Older adults often rely on altered connectivity patterns between the DMN and ECN to process social context and partner similarity during economic games, indicating that the brain dynamically reorganizes to preserve normative social behavior and protect against exploitation as cognitive architectures change over time 35.
Third-Party Punishment and Social Uncertainty
A critical feature of human fairness is third-party punishment (TPP) - the willingness of an unaffected observer to incur a cost to penalize a norm violator. TPP is essential for maintaining large-scale social cooperation equilibrium 3637. Neural activation during TPP paradigms strongly implicates the temporoparietal junction (TPJ) and the medial prefrontal cortex, regions central to the brain's mentalizing system, suggesting that intent attribution is a core component of fairness enforcement 3638.
However, TPP is highly vulnerable to uncertainty. Real-world social contexts frequently obscure whether a norm violation was intentional (intent uncertainty) or whether the unequal outcome was accidental (outcome uncertainty) 37. Emerging research indicates that uncertainty induces negative affective states, such as anxiety, which can disrupt the typical neural pathways governing prosocial behavior and norm enforcement. In contexts of high uncertainty, individuals must recalibrate their punishment thresholds, relying heavily on the dlPFC to weigh the risk of unjustly punishing a cooperative partner against the risk of allowing a genuine transgressor to exploit the system 37. Furthermore, individual differences in Social Value Orientation (SVO) - whether an individual is inherently "prosocial" or "proself" - modulate the brain's electrophysiological responses to unfairness. Prosocial individuals exhibit a stronger fairness effect on the medial frontal negativity (MFN) component, making them significantly more likely to engage in costly punishment when faced with norm violations 4.
Cross-Cultural Variances in Fairness Conceptualization
While the cognitive capacity to track investments, compute predictions, and evaluate social partners is a universal evolutionary endowment, the specific parameters defining "what is fair" are heavily dictated by cultural frameworks. Cross-cultural research systematically reveals that the heuristics utilized to allocate resources - and the severity of punishment directed at those who violate them - vary dramatically depending on a society's economic structure, historical institutions, and ontological worldview 3940.
Defining Equality, Equity, and Need
In the psychological, sociological, and anthropological literature, fairness is typically operationalized through the lens of distributive justice, which dictates how assets, opportunities, and privileges ought to be allocated within a community 41. Distributive justice generally relies on three distinct and often competing principles: 1. Equality (Sameness): Allocating identical resources to all individuals, irrespective of input, competence, or baseline condition 4243. 2. Equity (Proportionality/Merit): Allocating resources in strict proportion to individual effort, tangible investment, or societal contribution 214041. 3. Need: Allocating resources based on the varying baseline requirements of individuals, providing disparate levels of support to achieve equivalent life outcomes and systemic equity 214243.
A persistent historical bias in psychological and behavioral economic research is the profound overrepresentation of WEIRD (Western, Educated, Industrialized, Rich, and Democratic) populations 8444546. In Western, market-driven societies, fairness is predominantly framed through an individualistic lens that prioritizes equity and meritocratic proportionality 39404147. Social systems and legal structures in WEIRD populations are largely designed to reward individual accountability and personal contribution, aligning closely with transactional conceptualizations of justice where outcomes are strictly tied to inputs 404748.
Distributive and Procedural Justice in Non-WEIRD Contexts
When research expands beyond WEIRD demographics, fundamental differences in the ontological perception of fairness emerge. In many non-Western contexts - such as East Asian Confucian societies, traditional African communitarian societies, and Indigenous populations in Latin America - distributive justice is inextricably linked to social harmony, relational obligations, and collective well-being rather than isolated individual merit 49505155.
In African bioethics and traditional jurisprudence, for instance, ethical decision-making is grounded in relational personhood and communitarianism. Distributive justice is achieved not by rewarding the most productive individual, but by ensuring collective survival, mitigating distress within the extended community, and allocating resources based on familial or communal membership 505253. The Western prioritization of individual autonomy is often viewed as socially disruptive in these contexts; instead, collective health and harmony supersede individual accumulation 5052.
Similarly, in societies strongly influenced by Confucian traditions, maintaining interpersonal harmony and respecting hierarchical social roles are paramount 4955. Research on procedural justice (the perceived fairness of the processes used to allocate resources) in East Asian organizations reveals that employees may tolerate unequal distributive outcomes if the allocation process preserves social cohesion and respects institutional hierarchies 4955. In cultures with a high Power Distance Index, asymmetric resource allocation is often perceived not as an unfair violation of equity, but as an appropriate and stabilizing reflection of necessary societal structure 5055. Furthermore, ecological distribution conflicts involving Indigenous communities highlight that fairness often extends beyond human resource allocation to encompass "planetary justice" - integrating considerations of multi-generational time, sacred space, and reciprocal obligations to non-human entities, a framework entirely absent from standard Western economic models 54.
Economic Game Outcomes Across Global Societies
The profound impact of cultural paradigms on fairness is most starkly illustrated in cross-cultural behavioral economic experiments, specifically the Ultimatum Game (UG) and the Dictator Game (DG). In a classic UG played among WEIRD university students, proposers typically offer between 40% and 50% of the total endowment to the responder, and responders routinely reject offers below 20% or 30% 129555657. The rejection of "free money" in these contexts demonstrates a powerful aversion to inequity and a willingness to pay a premium to punish norm violators 25556. In the DG, where responders cannot reject the offer, offers drop significantly but rarely to zero, indicating an internalized normative pressure to appear fair even without the threat of punishment 5658.
However, seminal field research by anthropologists administering these games in small-scale, non-WEIRD societies disrupted the assumption that the 50/50 split and aggressive negative reciprocity are human universals 4659. Both the proposed allocations and the rejection thresholds diverge significantly from Western baselines depending on the society's specific ecological niche and social structure.
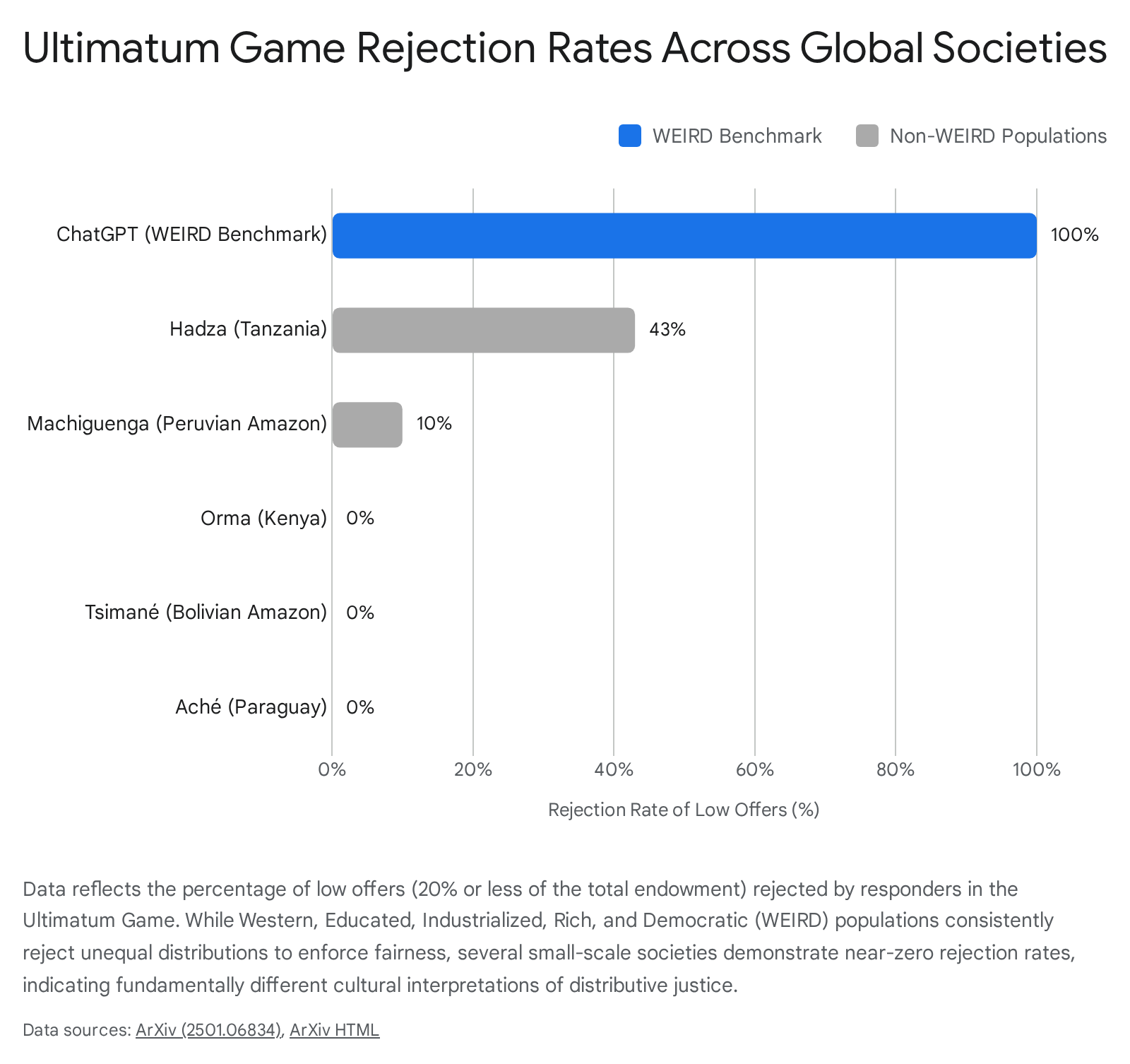
| Society (Region/Type) | Mean UG Offer (% of total) | Rejection Rate for Offers ≤ 20% | Cultural Context & Implications |
|---|---|---|---|
| WEIRD Populations (e.g., US/Europe) | 40% - 45% | ~ 40% - 50% | High market integration relies on abstract rules of fair transaction and generalized trust among strangers. Unfairness is heavily punished to maintain systemic integrity 15559. |
| Hadza (Tanzania, Hunter-Gatherers) | 26% - 33% | 43% | High rejection rates for low offers reflect a culture of rigorous, enforced egalitarianism and immediate food sharing designed to prevent any individual from achieving dominance 65. |
| Machiguenga (Peruvian Amazon, Horticulturalists) | 26% | 10% | Society comprises highly independent family units with little community-level economic reliance. Low offers are expected, and any positive amount is accepted without moral outrage 5965. |
| Orma (Kenya, Pastoral-Nomadic) | 31% | 0% | Behavior is influenced by moderate market integration, but low offers are rarely punished through costly rejection, reflecting different norms of wealth distribution 65. |
| Tsimané (Bolivian Amazon, Forager-Horticulturalists) | 30% | 0% | No rejections observed. Daily social life does not feature the kind of market-based bargaining that necessitates strict equity enforcement or punishment 5965. |
| Aché (Paraguay, Hunter-Gatherers) | 40% - 51% | 0% | Extremely high baseline offers reflecting a deeply ingrained cultural norm of pervasive, unquestioned resource sharing, rendering punitive rejection entirely unnecessary 5965. |
The variability across these societies correlates strongly with their degree of market integration and the local payoffs to cooperation 4659. In societies where daily survival requires collective action and high interdependence (e.g., cooperative hunting among the Aché), fairness norms are deeply internalized, leading to spontaneously generous offers. In societies composed of economically independent family units (e.g., the Machiguenga), fairness is not viewed through the lens of transactional equity with strangers, resulting in low offers and an absence of punitive rejections 465965. Furthermore, altering the stakes of the Dictator Game reveals that while WEIRD populations maintain their allocation proportions relatively consistently, extremely high stakes in certain contexts can erode fairness preferences, driving dictators to keep larger proportions of the endowment 565758.
Hyper-Fairness and Non-Monotonic Rejection Rates
Further complicating the universality of Western fairness models is the phenomenon of non-monotonic rejection rates. In WEIRD populations, the willingness to accept an offer generally increases monotonically with the size of the offer; it is implicitly assumed that a higher offer is intrinsically better and will always be accepted 60.
However, in several non-WEIRD populations, researchers observe a distinct hump-shaped willingness-to-accept curve. Participants in these societies punish extreme greed by rejecting very low offers, but they also frequently reject "hyper-fair" offers that exceed 50% or approach 100% of the initial endowment 60. For example, in large-scale studies conducted across multiple ethnic groups in the Democratic Republic of the Congo (DRC) and among certain populations in Henrich's multi-society dataset, rejection rates spike significantly for offers above 50% 60.
In these cultural contexts, an excessively generous offer from a stranger is not interpreted as a mere windfall of good fortune. Instead, it is perceived as highly suspicious, potentially insulting, or a deliberate attempt to impose an unpayable social debt. Under the local norms of relational morality and gift economies, accepting a hyper-fair offer permanently alters the power dynamic, placing the recipient in an unwanted state of deep obligation or subordinate status 960. Thus, rejecting a highly advantageous offer is an assertion of autonomy and a refusal to participate in a socially disruptive exchange, providing definitive evidence that human fairness is evaluated not strictly by material outcome, but by the complex relational and ontological implications of the distribution.
Conclusion
The profound human value placed on fairness is neither an arbitrary cultural artifact nor a simple biological reflex. It is the sophisticated product of an evolutionary history where survival depended heavily on success within a competitive biological market. Through the mechanism of partner choice, natural selection favored individuals equipped with the strategic vigilance to continuously monitor distributions, demand equitable treatment to avoid exploitation, and project a reputation of reliability and impartiality to secure future cooperative opportunities.
This evolutionary scaffolding provides the foundation for the neurobiological architecture of fairness. The human brain utilizes dual computing systems - relying on the anterior insula and the salience network to generate rapid, aversive signals to inequity, the ventromedial prefrontal cortex to process the reward of social cohesion, and the dorsolateral prefrontal cortex to exert the executive control necessary to enforce norms even at a steep personal cost.
Yet, while the neural hardware for computing prediction errors and tracking fairness is a universal human trait, its operational software is entirely calibrated by culture. Developmental and cross-cultural research conclusively demonstrates that the conceptual leap from recognizing basic inequality to enacting formal equity requires intense socialization. Whether fairness is defined as strict meritocratic equity, relational harmony, or communal need depends intimately on a society's history, economic structure, and prevailing moral philosophy. Understanding human fairness, therefore, requires recognizing it as a dynamic construct: an evolved biological capacity to navigate interdependence, optimized through the unique, localized lens of cultural justice.