Evolution of Human Monogamy and Pair-Bonding
Introduction to Human Pair-Bonding
The question of whether humans are naturally monogamous occupies a central position in evolutionary biology, physical anthropology, and sociological inquiry. Determining the ancestral mating system of Homo sapiens is heavily complicated by the species' exceptional behavioral flexibility, the divergence between social pair-bonding and genetic monogamy, and the profound influence of culture, technology, and ecological constraints on reproductive strategies. Unlike the majority of mammalian species, which display rigidly canalized and predictable mating systems, humans exhibit a vast spectrum of pair-bonding arrangements. This spectrum ranges from strict life-long monogamy to serial monogamy, polygyny, polyandry, and varying degrees of consensual non-monogamy 12.
To accurately reconstruct the ancestral human mating system, researchers must synthesize evidence across multiple independent scientific domains. Comparative primatology and human anatomy offer physiological markers of historical sexual selection, providing fossilized clues through skeletal sexual dimorphism and the soft-tissue comparative analysis of testicular volume 34. Evolutionary game theory provides mathematical models for understanding the adaptive payoffs of cooperative breeding, paternal investment, and state-dependent risk-taking under conditions of prehistoric resource scarcity 567. Furthermore, the ethnographic record, particularly extensive analyses of the Standard Cross-Cultural Sample (SCCS), details the socioecological distribution of contemporary and historical marriage practices across hundreds of pre-industrial societies 12. Finally, recent advances in paleogenomics and the high-resolution sequencing of ancient DNA (aDNA) allow researchers to map prehistoric kinship networks directly, illuminating the actual mating, inheritance, and residential practices of early human communities with unprecedented accuracy 8910.
Synthesizing this multidisciplinary data indicates that the ancestral human baseline is not strictly monogamous in the obligate manner of certain avian species, nor is it fiercely promiscuous like the social structures of chimpanzees. Instead, hominins likely evolved a system of predominant social monogamy or serial monogamy, overlaid with facultative polygyny and varying rates of extra-pair copulation. This flexible mating architecture was heavily supported by, and evolved in tandem with, deep cooperative alloparenting networks 211.
Biological and Anatomical Markers of Sexual Selection
In evolutionary biology, morphological characteristics frequently serve as a physiological archive of a species' mating history. Because behavioral patterns and soft tissues do not fossilize, anthropologists analyze contemporary human anatomy - specifically sexual dimorphism and reproductive physiology - in comparison with other great apes to infer the historical intensity of both intra-sexual competition (competition for mates) and sperm competition (post-copulatory competition) throughout human evolution 1213.
Sexual Size Dimorphism and Contest Competition
Sexual size dimorphism (SSD) - the divergence in body mass and physiological weaponry between males and females of the same species - is a highly reliable indicator of mating systems across the primate order. In mammalian species where a single male monopolizes mating access to multiple females, intra-sexual contest competition is intense. This dynamic creates heavy selective pressure for male body mass, aggressive behavior, and specialized anatomical weaponry, such as larger canines or sagittal crests 121314. Conversely, species characterized by strict monogamy exhibit minimal sexual dimorphism, as both sexes experience similar reproductive variance and relatively low levels of physical competition for exclusive mates 1213.
The extant primate record demonstrates this continuum clearly. Gorillas, which operate in single-male, multi-female harem structures, exhibit extreme dimorphism. Male silverback gorillas are often more than twice the size of females, with a male-to-female mass ratio of approximately 2.37 1215. Their skeletons feature massive sagittal and nuchal crests to support heavy jaw and neck musculature utilized in male-male combat 13. Orangutans display similar levels of evolutionary divergence, with a mass ratio of 2.23, reflecting a system of solitary, transient polygyny where dominant males defend vast territories containing multiple dispersed females 12. At the opposite end of the morphological spectrum, socially monogamous gibbons exhibit almost no sexual size dimorphism, with a male-to-female mass ratio approaching 1.0. Male and female gibbons are equally aggressive in defending their shared territories, resulting in monomorphic skeletal and muscular development 31316. Chimpanzees and bonobos, which possess multi-male, multi-female promiscuous mating systems, exhibit moderate dimorphism, with male-to-female mass ratios of 1.29 and 1.36, respectively 1217.
Modern humans display a mild to moderate degree of overall sexual size dimorphism. The average human male-to-female mass ratio is approximately 1.15, placing humans between the strictly monomorphic gibbons and the moderately dimorphic chimpanzees 312. Based purely on overall mass and skeletal height, this metric strongly suggests an evolutionary history characterized by relatively low pre-copulatory contest competition. This anatomical reality aligns more closely with a history of monogamous or serially monogamous pair-bonding than with harem-style polygyny 23.
However, critiques of pure mass ratios suggest that total body weight obscures specific tissue dimorphism. When assessing muscle mass independently of fat, human sexual dimorphism becomes significantly more pronounced. Men possess roughly 40% more fat-free mass and 60% more total lean muscle mass than women 17. Furthermore, human males feature 80% greater upper-body muscle mass and 50% greater lower-body muscle mass compared to females 17. This significant divergence in musculature indicates that while historical male-male physical contest competition was less extreme than in gorillas, it remained a distinct selective pressure throughout human evolution. This muscular dimorphism reflects a mating system that likely accommodated some degree of polygyny or episodic, intense male competition for mates, preventing the hominin lineage from evolving the complete physical parity seen in gibbons 1417.
Craniofacial and Skeletal Dimorphism
Skeletal structure provides additional context regarding the evolutionary trajectory of human sexual dimorphism. In general, skeletal dimorphism in primates is primarily a byproduct of body mass dimorphism, but specific cranial and dental features offer distinct evolutionary clues 13.
The breadth, length, and height of the neurocranium in adult male gorillas and orangutans are roughly 9% larger than in adult females of the same species. In contrast, gibbons exhibit a neurocranial sex difference of only 4% to 5% 13. Human cranial dimorphism is relatively low, supporting the hypothesis of decreased contest competition over time. More critically, the fossil record of early hominins reveals a distinct trajectory regarding canine tooth size. While species like Australopithecus afarensis exhibited marked dimorphism in overall body size, their canine size was already significantly smaller than that seen in modern chimpanzees 12. The continuous reduction in canine size from early hominins to modern Homo sapiens indicates a steady evolutionary shift away from utilizing anatomical weaponry for male-male mating competition, further supporting a deep historical trend toward pair-bonding and cooperative social structures 12.
Testicular Anatomy and Sperm Competition
While overall body mass, muscular distribution, and skeletal weaponry reflect pre-copulatory contest competition, testicular volume serves as the primary biological marker for post-copulatory sperm competition. In species where females frequently mate with multiple males during a single estrus cycle (multi-male promiscuity), males face intense selective pressure to produce massive quantities of sperm. This biological imperative allows them to outcompete the genetic material of rival males directly within the female reproductive tract 319.
Comparative analysis of relative testis mass - defined as testis weight as a percentage of overall body weight - reveals profound evolutionary divergence among hominoids 420.
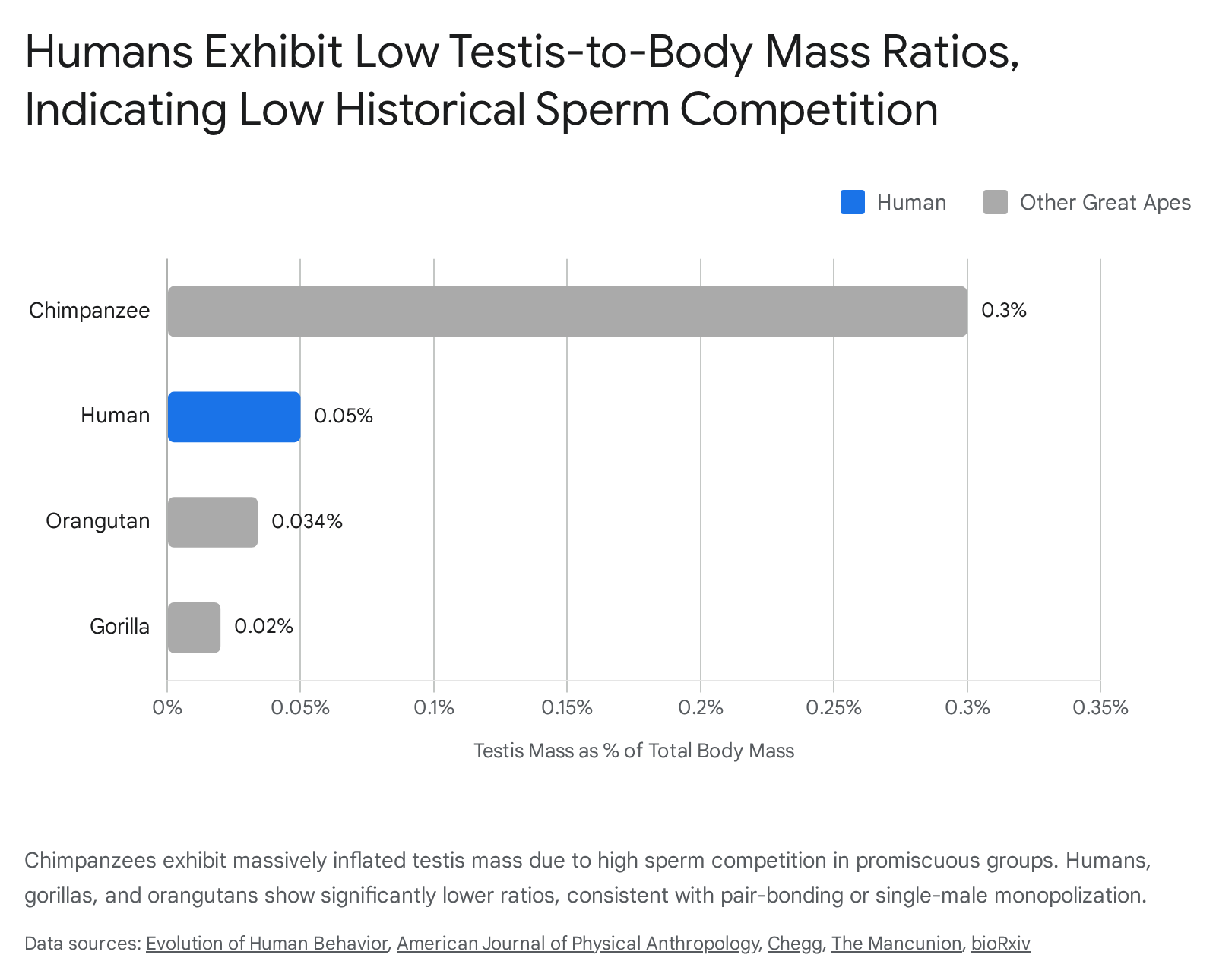
Chimpanzees and bonobos possess highly promiscuous mating structures, leading to extreme sperm competition. To accommodate the necessary continuous sperm production, male chimpanzees have evolved massive testicles, weighing between 150 and 170 grams, which can account for roughly 0.30% of their total body weight 192122. These testicles are highly specialized at the microscopic level, possessing a high ratio of seminiferous tubules to interstitial tissue to maximize spermatogenesis rates 22. In stark contrast, male gorillas, despite weighing upwards of 200 kilograms, have extremely small testicles weighing approximately 10 grams, which is roughly 0.02% of their body weight 1921. Because a silverback gorilla has exclusive sexual access to his harem, he faces almost zero sperm competition and thus requires minimal caloric and biological investment in testicular tissue 3423.
Human testicles range from 16 to 50 grams, representing roughly 0.04% to 0.08% of total body weight 212218. This relative mass occupies a distinct intermediate position. It is significantly larger proportionally than that of a gorilla, but drastically smaller than that of a chimpanzee 323. The intermediate size of human testes implies that while ancestral human females occasionally mated with multiple males - preventing the evolution of gorilla-like micro-testes - sperm competition was relatively low, firmly precluding the hyper-promiscuity seen in chimpanzee societies 319.
Genomic and cellular studies corroborate this morphological data. Rates of sequence evolution linked to sperm competition place humans far below chimpanzees. Research analyzing the genetic sequences of 55 primate species demonstrates that primates with larger testes experience higher rates of genome evolution, likely as a byproduct of greater sperm production and subsequent mutation 19. Human sequence evolution aligns with low-competition species. Furthermore, human sperm exhibits physiological traits, such as lower sperm concentration, slower swimming speed, and higher rates of morphological abnormality, that more closely align with the low-competition environment of the gorilla than the high-competition environment of the chimpanzee 19.
| Primate Species | Dominant Mating System | Sexual Dimorphism (Male:Female Mass Ratio) | Approximate Absolute Testes Weight | Testes as % of Total Body Weight |
|---|---|---|---|---|
| Gorilla | Single-male, multi-female (Polygyny) | Extreme (2.37) | ~10 g | ~0.02% |
| Orangutan | Solitary, transient polygyny | High (2.23) | ~35 g | ~0.03% |
| Chimpanzee | Multi-male, multi-female (Promiscuity) | Moderate (1.29) | 150 - 170 g | ~0.30% |
| Human | Social Monogamy / Mild Polygyny | Low-Moderate (1.15) | 16 - 50 g | ~0.04 - 0.08% |
| Gibbon | Strict Territorial Monogamy | Minimal (~1.00) | < 5 g | < 0.10% |
| Table 1: Comparison of physiological markers of sexual selection across primate species, illustrating the intermediate traits of modern humans. 31215192118 |
Evolutionary Game Theory and Cooperative Breeding
Beyond physical anatomy, the evolution of human pair-bonding has been extensively modeled using evolutionary game theory and behavioral ecology. The central paradox in human evolution is the development of a highly altricial (helpless) infant with an extraordinarily long maturation period. Human offspring require an immense caloric investment, estimated by anthropologists to demand roughly 13 million calories from birth to nutritional independence around age eighteen 20. No single female hominin could reliably provision such an offspring alone while securing her own survival, leading to intense selective pressures for social structures that secured reliable caloric subsidies and protective care 112721.
Critiques of the Paternal Provisioning Hypothesis
Historically, the dominant explanation for the evolution of human monogamy was the "Man the Hunter" or paternal provisioning hypothesis, popularized in the mid-twentieth century. This model posited a clean, evolutionarily driven division of labor: males hunted and provided highly caloric meat to a specific, sexually exclusive female. In exchange for this continuous provisioning, the female provided paternity certainty, thereby forming the nuclear pair-bond and establishing the foundation of human social structure 202122.
However, feminist anthropologists and behavioral ecologists, notably Sarah Blaffer Hrdy and Barbara Smuts, have systematically dismantled the strict assumptions of this model. Cross-cultural studies of contemporary foraging societies, such as the Hadza of Tanzania, reveal that large game hunted by men is usually distributed widely and egalitarianly across the entire camp, rather than being funneled exclusively to a man's biological offspring 2021. Because male provisioning of specific households is frequently unreliable due to the inherent high variance and failure rate of hunting large game, female hominins could not rely solely on male mates for survival 2123.
Consequently, the assumption that monogamy evolved purely as a direct economic trade of female fidelity for male meat is insufficient 2123. Furthermore, the traditional theory fails to account for the fact that strict, exclusive paternal care is exceedingly rare in mammalian species, particularly among closely related primates, making its sudden, isolated emergence in hominins as the sole driver of pair-bonding highly unlikely 23. Studies indicate that female gathering activities consistently provided the vast majority of reliable caloric intake in ancestral environments, rendering females far less economically dependent on male hunters than the traditional model assumed 2122.
The Cooperative Breeding Hypothesis
The modern consensus in evolutionary anthropology emphasizes the Cooperative Breeding Hypothesis as the primary driver of human social evolution. Rather than relying solely on a monogamous pair-bond, human child-rearing relies heavily on "alloparents" - individuals other than the biological parents who assist in care, protection, and provisioning 1124. This extended network includes grandmothers, aunts, older siblings, and non-kin community members 1123.
The "Grandmother Hypothesis," specifically, posits that post-reproductive females played a vital evolutionary role. By foraging for reliable staple foods, such as underground storage organs (tubers), grandmothers subsidized the extreme caloric demands of weaned children 2021. This alloparental support allowed hominin mothers to maintain shorter inter-birth intervals compared to other great apes, driving population expansion despite the extreme dependency of human infants 2024. Cooperative breeding also necessitated advanced social cognition; human infants evolved to be hyper-aware of the intentions and moods of caregivers to elicit necessary attention, fostering the deep intersubjectivity and empathy characteristic of the species 2724.
State-Dependent Risk-Taking and Male Investment
In the context of cooperative breeding, male pair-bonding and paternal investment did evolve, but they functioned within a broader ecological matrix rather than as a solitary adaptation. Game-theoretic models suggest that as cooperative networks expanded, the evolutionary payoff for a male remaining with a single female and ensuring the survival of his specific offspring began to outweigh the potential, but highly variable, payoffs of seeking multiple mates 721.
The application of state-dependent risk-taking models, as explored by evolutionary biologists like Bergstrom and Meacham, mathematically explains how environmental variance influences mating strategies 5. These models distinguish between need-based and ability-based risk-taking. In environments where infant mortality is exceedingly high without both biparental and alloparental care, the high-risk gamble of a male abandoning a mate to pursue polygyny yields an evolutionary payoff of zero if none of the neglected offspring survive to reproductive age 5. Thus, monogamy functions as a low-risk, steady-payoff evolutionary strategy under conditions of severe resource constraint and offspring dependency 6.
Furthermore, game theory highlights the role of male-male contest competition and infanticide in the origins of pair-bonding. In primate societies, incoming dominant males frequently commit infanticide to return nursing females to estrus 732. Mate guarding may have initially evolved as a defensive strategy to protect infants from rival males. This necessity led to prolonged, continuous proximity between males and females, which subsequently exapted into emotional pair-bonding and direct paternal provisioning 73233.
The hormonal infrastructure of modern human males supports this evolutionary pathway. Unique among great apes, interactions with infants consistently trigger distinct neuroendocrinological shifts in human fathers: testosterone levels measurably decrease, while oxytocin and prolactin levels elevate 32. This hormonal transition downregulates male aggression and facilitates deep emotional attachment to both the infant and the mother, providing a biological mechanism that cements the pair-bond beyond mere economic exchange 32.
Cross-Cultural Ethnography and Marriage Systems
While the biology of humans leans toward intermediate pair-bonding shaped by cooperative breeding, analyzing how this is reflected in the structural organization of global human societies requires extensive ethnographic data. Anthropological datasets, specifically the Standard Cross-Cultural Sample (SCCS) comprising 186 culturally distinct, predominantly pre-industrial societies, provide a comprehensive map of human mating variation across diverse socioecological environments 12.
The Prevalence of Polygyny Versus Monogamy
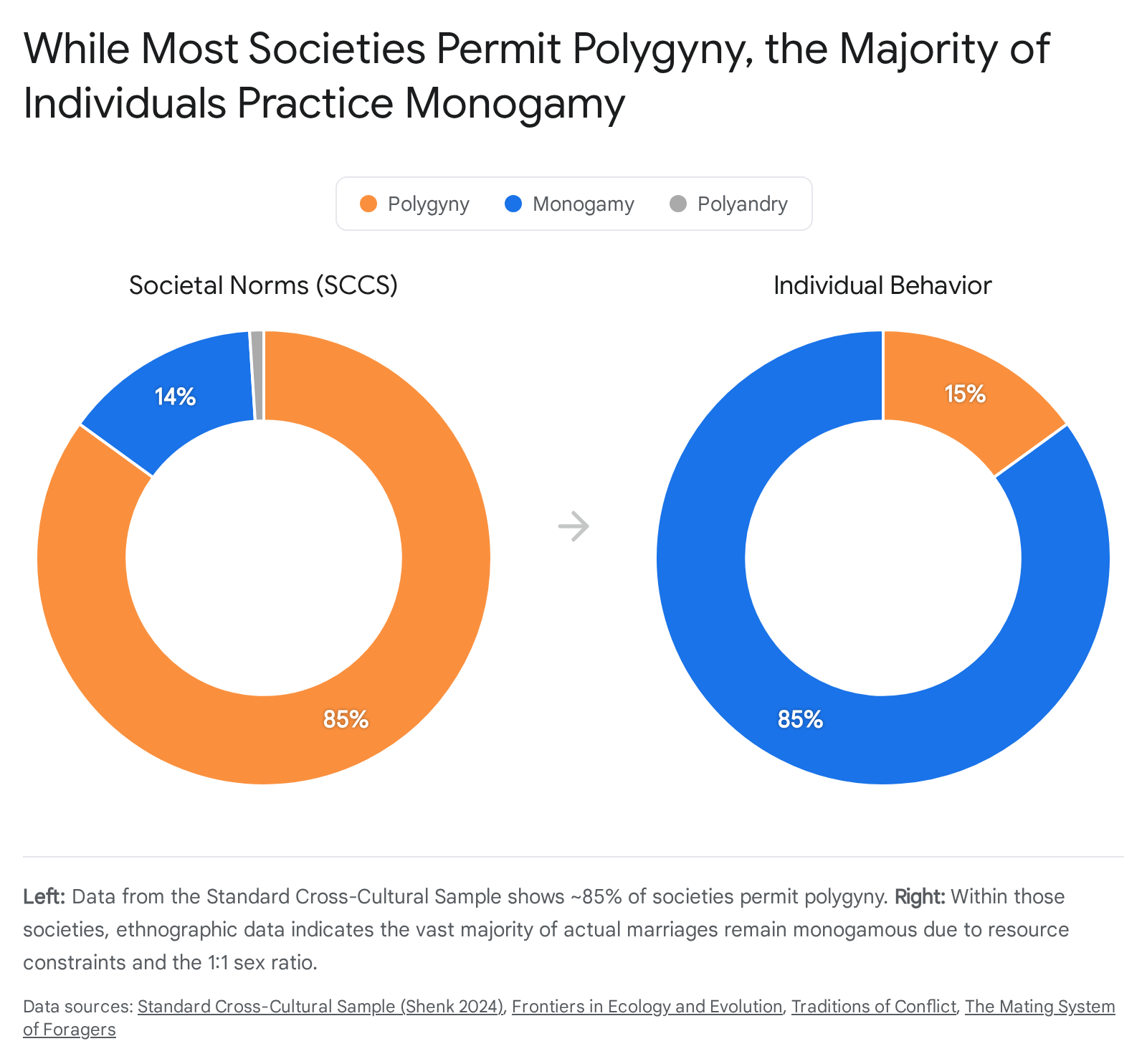
A superficial reading of cross-cultural anthropological data often leads to the erroneous conclusion that humans are inherently and overwhelmingly polygynous. According to the SCCS, approximately 83% to 85% of documented human societies allow or socially sanction polygyny (specifically, one man marrying multiple women) 122526. Roughly 15% to 16% of societies prescribe strict normative monogamy, explicitly forbidding plural marriage. Polyandry (one woman marrying multiple men) is exceedingly rare, documented in fewer than 1% of societies. Where polyandry does exist, it is almost exclusively found in specific ecological niches, such as the Tibetan highlands, where extreme land scarcity demands that brothers pool resources and share a single wife to prevent the division of inheritances 12526.
However, the statistic that 85% of societies permit polygyny is frequently misinterpreted in popular science to mean that most individuals within those societies actively practice it. In reality, within almost all socially polygynous societies, the overwhelming majority of actual marriages remain firmly monogamous 236.
The Polygyny Threshold and Resource Monopolization
Ethnographic data reveals that typically only 10% to 20% of men in a permitted-polygyny society actually acquire multiple wives. Maintaining multiple spouses and their subsequent offspring requires massive wealth, political power, and resource monopolization that is inaccessible to the average male 136. This phenomenon is described by the "polygyny threshold model," a framework drawn from behavioral ecology. The model posits that females will only choose to enter a polygynous union - agreeing to share a husband's resources and attention - if that male's resources are so vast that a divided share is still functionally superior to the full wealth of an exclusive, but lower-status, monogamous male 2627.
For example, among the Savanna Pumé foragers of South America, while polygyny is permitted, over 80% of women are exclusively in monogamous relationships 2. Similarly, among the Dobe Ju/'hoansi hunter-gatherers of the Kalahari, early anthropological surveys demonstrated that 93% of married men lived monogamously 36. Because the human sex ratio at reproductive age naturally hovers near 1:1, mathematical realities dictate that high rates of polygyny inevitably leave massive swaths of the male population entirely unmarried 28.
Social Consequences of High Polygyny
When societal wealth inequality increases - such as during the transition from egalitarian foraging to intensive agriculture and pastoralism - elite males gain the capacity to hoard resources and accumulate vast numbers of wives 27. However, this extreme reproductive skew generates severe social externalities.
Cross-cultural data demonstrates that societies with high functional rates of polygyny experience heightened levels of intra-societal violence, crime, and instability. The monopolization of women by elite males creates a large, disenfranchised pool of low-status, unmarried men who engage in high-risk, aggressive behaviors - including homicide, theft, and warfare - to acquire the resources necessary to enter the marriage market 28. Furthermore, highly polygynous households exhibit lower average genetic relatedness among co-residents, which correlates statistically with elevated risks of intra-household abuse, neglect, and conflict compared to monogamous households 28. Consequently, while elite males continuously push toward polygyny to maximize their individual reproductive success, the ecological baseline for the vast majority of human beings across the globe is behavioral monogamy. This is often practiced sequentially as "serial monogamy," wherein individuals have multiple exclusive partners over a lifetime due to divorce or spousal mortality 12.
| Marriage System | Societal Permission (SCCS %) | Estimated Individual Practice Globally | Key Drivers & Ecological Context |
|---|---|---|---|
| Monogamy (Strict/Serial) | ~15-16% (Prescribed) | > 80% | Default for majority due to 1:1 sex ratio and resource constraints. |
| Polygyny | ~83-85% (Permitted) | < 20% | Wealth inequality, elite male resource monopolization, agricultural/pastoral surplus. |
| Polyandry | < 1% | < 1% | Extreme resource/land scarcity requiring male resource pooling (e.g., fraternal polyandry). |
| Table 2: Comparison of human marriage systems, contrasting macro-societal rules against micro-individual behaviors based on the Standard Cross-Cultural Sample. 12252636 |
Paleogenomics and Prehistoric Social Structures
While physiological markers and modern hunter-gatherer ethnography allow for robust inferences, they remain indirect measures of the past. However, the recent methodological revolution in paleogenomics - specifically the extraction and sequencing of ancient DNA (aDNA) from archaeological remains - provides direct, empirical evidence of the actual mating patterns, kinship networks, and social organizations of prehistoric humans 829.
Matrilineal Organizations in Early Agrarian Societies
Traditional anthropological and archaeological theory frequently assumed that the transition to agriculture inevitably and universally ushered in strict patriarchal and patrilineal systems, as men sought to secure the inheritance of accumulated property and land. However, genomic analysis of Late Neolithic sites demonstrates a vastly more complex, flexible, and varied social landscape.
A landmark 2025 study examining aDNA from 131 skeletons at Çatalhöyük, a 9,000-year-old proto-city in modern-day Turkey, revealed unexpected kinship patterns 930. Researchers tracking intergenerational biological relationships within domestic burial clusters discovered that familial continuity was predominantly matrilineal 931. DNA sequences mapping mitochondrial genomes showed that women were the genetic anchors of the household, while males frequently originated from different households or outside communities 3032. This genetic signature strongly indicates matrilocal residence patterns, where the groom moved in with the bride's family upon pair-bonding 30. Furthermore, biological analysis of grave goods revealed that female infants and children were interred with up to five times more offerings than males. This material disparity underscores a female-centered social structure, decisively refuting the assumption that early agrarian societies originating in Anatolia were universally male-dominated 93133.
Similar findings emerged from the 5,000-year-old Fujia site in Shandong, China. Published in 2025, paleogenomic analyses of 60 individuals across two cemeteries revealed strong matrilineal organization 10. Through a combination of isotopic analysis and genome sequencing, researchers confirmed this community practiced high endogamy (mating strictly within the local population) with remarkable social stability over a period of 250 years 10. The Fujia community was completely organized around female descent lines, providing a rare and pristine example of a matrilineal and matrilocal kinship system operating successfully in prehistoric East Asia without patriarchal dominance 10.
Consanguinity and Patrilineal Shifts
Conversely, ancient DNA also records distinct regional transitions toward intensive patrilineal systems and extended genetic family networks. Analyses of the Late Neolithic Longshan culture in central China (c. 2275 - 1844 BCE) utilizing runs of homozygosity (ROH) - a genomic metric used for identifying inbreeding - revealed evidence of consanguineous mating 834. The genetic data from these sites indicates deep paternal kinship ties that extended well beyond the nuclear family, showing that as social complexity intensified during the Longshan period, specific communities transformed into extended, genetically cohesive patrilineal family units 834.
Social Kinship Versus Biological Relatedness
Moreover, aDNA analysis demonstrates that prehistoric humans frequently operated with conceptions of "social kinship" that superseded strict biological relatedness. The definition of family in antiquity was highly adaptable. At the post-Roman site of Worth Matravers in the United Kingdom, researchers analyzed multiple burials containing individuals interred together in double or triple graves 35. Genomic sequencing revealed that these individuals were genetically unrelated, indicating that household or community identity frequently transcended strict genetic lineage 35. As modern anthropological geneticists advocate, biological relatedness does not automatically equate to cultural kinship; paleogenomic data proves humans flexibly utilized both biological pair-bonds and fictive kinship networks to adapt to ecological pressures and construct stable societies 36.
Genomic Evidence of Admixture Mating with Neanderthals
Ancient DNA also provides highly specific insights into human mating behaviors prior to the extinction of other hominin species. A 2026 genomic study analyzed the introgression of Neanderthal DNA into modern Homo sapiens. By analyzing the severe, unexpected depletion of Neanderthal DNA specifically on the human X chromosome, researchers inferred the precise directional dynamics of ancient interbreeding events occurring roughly 45,000 years ago in Eurasia 3748.
Because genetic females inherit two X chromosomes and genetic males inherit one, the specific genomic signature observed in modern non-African populations indicates a strong historical preference for mating between male Neanderthals and female modern humans, rather than the reverse 3748. While the exact behavioral or demographic mechanisms driving this asymmetry remain obscured - whether due to social preference, physical dominance, or the lower survival rates of offspring from human males and Neanderthal females - this data provides unparalleled resolution regarding the deep-time sexual encounters and hybrid pair-bonding of our species 3748.
Modern Behavioral Flexibility and Consensual Non-Monogamy
The culmination of millions of years of hominin evolution has not resulted in a biologically deterministic mandate for strict, lifelong monogamy. Instead, it has produced a profound behavioral flexibility capable of navigating vast cultural and ecological shifts. This adaptability is clearly reflected in contemporary sociological trends surrounding marriage, mating, and the conscious re-engineering of the pair-bond.
Prevalence and Demographics of Contemporary Non-Monogamy
In recent years, the practice of Consensual Non-Monogamy (CNM) - an umbrella term encompassing polyamory, open relationships, swinging, and relationship anarchy - has entered mainstream sociological analysis 3839. Recent survey data indicates that approximately 4% to 5% of the United States population currently practices some form of CNM 3839. Furthermore, lifetime prevalence is notably higher; up to 14.5% of U.S. adults report having practiced CNM, and approximately 20% of single Americans report having engaged in open relationships at some point in their lives 3839.
Data from comprehensive 2024 and 2025 surveys, encompassing thousands of practitioners worldwide, reveal that the demographics of CNM are heavily intersectional. Polyamorous individuals are significantly more likely to identify as LGBTQ+ compared to the general population; in some survey cohorts, up to 75% of non-monogamous respondents identify as something other than heterosexual 385140. This demographic overlap highlights a broader rejection of traditional, cis-heteronormative relationship structures in favor of highly customized emotional and sexual networks 5140. The data also shows a generational shift, with Gen Z respondents being twice as likely to engage in open relationships compared to the overall population, indicating evolving cultural attitudes toward pair-bonding exclusivity 53.
Psychological Outcomes and Societal Stigma
Despite practicing alternative mating strategies that deviate from the socially prescribed norm, individuals engaged in CNM report positive psychological and relational outcomes. Statistical analyses reveal exceptionally high relationship satisfaction rates (up to 85%) among polyamorous individuals 38. When compared to strictly monogamous cohorts, CNM practitioners frequently report higher scores in emotional intelligence, better communication regarding boundaries, and lower levels of neuroticism and unmanaged jealousy 3839. This data indicates that human psychological architecture - while deeply wired for attachment and capable of extreme mate-guarding jealousy - can successfully adapt to and regulate complex, multi-partner emotional landscapes when those relationships are mediated by clear communication, equity, and informed consent 3839.
However, modern society remains heavily biased toward strict monogamy, often structurally penalizing deviations. Approximately 60% to 61% of polyamorous individuals report experiencing societal stigma, discrimination, or prejudice based on their relationship status 395140. This discrimination frequently manifests in material ways, including workplace prejudice, housing insecurity, bias in child custody disputes, and familial rejection 385140. The persistence of this stigma reflects the enduring cultural legacy of intensive agricultural and state-level societies, which structurally enshrined monogamy and strict inheritance laws, embedding nuclear family exclusivity into the foundational legal and religious doctrines of the modern era 3941.
| Metric / Characteristic | Consensual Non-Monogamy (CNM) Statistics |
|---|---|
| Current Prevalence (U.S.) | 4% - 5% of the adult population 3839 |
| Lifetime Prevalence | 14.5% of adults; 20% of singles have attempted CNM 3839 |
| LGBTQ+ Overlap | Highly correlated; up to 71-76% of surveyed practitioners identify as LGBTQ+ 5140 |
| Relationship Satisfaction | 85% reported satisfaction; generally higher communication scores than monogamous baselines 3839 |
| Societal Stigma/Discrimination | 60% - 61% report experiencing prejudice (workplace, housing, familial) 5140 |
| Table 3: Sociological metrics regarding the prevalence, demographics, and outcomes of Consensual Non-Monogamy in contemporary populations. 38395140 |
Conclusion
The question of whether monogamy is definitively "natural" for human beings defies a simplistic, binary answer. Biological, anthropological, and genomic evidence overwhelmingly indicates that humans are not biologically programmed for solitary, lifelong, absolute monogamy, nor are they adapted for unchecked, chimpanzee-like promiscuity.
Instead, human nature is characterized by a highly flexible, intermediate reproductive strategy. Our anatomy - marked by moderate sexual muscular dimorphism and intermediate testicular volume - points to an evolutionary past dominated by serial pair-bonding, featuring low to moderate levels of sperm competition and physical male contest competition. Evolutionary game theory and the biological realities of human infant altriciality demonstrate that strict pair-bonding, supported by extensive cooperative alloparenting networks, was an absolute survival imperative on the Pleistocene savanna.
Yet, as the global ethnographic record of the SCCS and recent paleogenomic data confirm, humans possess the psychological and cultural plasticity to adapt these foundational bonds into polygynous, matrilineal, polyandrous, or patrilineal arrangements. These structures are continuously negotiated based on resource availability, ecological stress, wealth distribution, and cultural scaffolding. Ultimately, human pair-bonding is a dynamic synthesis of our deep-seated biological drive for secure attachment and our unparalleled cultural capacity to deliberately re-engineer social relationships.