Evidence for mirror neurons in empathy and social brain science
The discovery of the mirror neuron system during the 1990s introduced a profound neurobiological framework for understanding how primates and humans perceive, interpret, and internalize the actions of others 12. Initially identified through single-cell recordings in the ventral premotor cortex of macaque monkeys, these highly specialized neurons exhibited a unique physiological property: they fired both when the animal executed a specific, goal-directed motor action and when the animal passively observed another agent performing the identical action 13. This bidirectional mapping mechanism suggested a direct physiological link between perception and motor execution, rapidly elevating mirror neurons to the center of theoretical debates regarding the evolution of social cognition, imitation, and language 45.
Over the subsequent decades, the concept of a human mirror neuron system was extrapolated to explain highly complex psychological phenomena, most notably empathy 67. Popular scientific discourse frequently labeled these cells as "empathy neurons," positing that they provided an automatic, innate, and unmediated simulation of other minds 68. However, as the fields of cognitive neuroscience, cultural psychology, and computational modeling have matured, the empirical evidence has forced a rigorous reevaluation of these early claims. Contemporary neuroscience indicates that the mirror neuron system does not operate as an isolated engine for empathy or mindreading 8910. Rather, social cognition relies on highly distributed, dynamic, and culturally modulated cortical networks, within which basic sensorimotor mirroring provides only lower-level, context-dependent computational inputs 1011.
Foundational Discovery and Anatomical Homology
The foundational evidence for mirror mechanisms derives from highly invasive electrophysiological research in non-human primates. In macaques, the mirror neuron system is localized primarily within two interconnected cortical hubs: area F5 of the ventral premotor cortex and areas PF and PFG of the anterior inferior parietal lobule 31213. These regions are deeply involved in the planning and execution of goal-directed movements, such as grasping, holding, and tearing 19.
Establishing a homologous network in the human brain has been a central endeavor for comparative neuroanatomy. Because single-neuron recordings in humans are exceedingly rare - limited primarily to opportunistic intracranial measurements during pre-surgical evaluations for epilepsy, which have nonetheless identified neurons with mirroring properties in the supplementary motor area and medial temporal lobe 14 - researchers have relied heavily on cytoarchitectonic comparisons, structural connectivity analyses, and functional neuroimaging to map the human mirror neuron system 1516.
Anatomical mapping and diffusion magnetic resonance imaging (MRI) demonstrate that macaque area F5 shares substantial structural and functional homology with human Brodmann areas (BA) 44 and 45 121317. These areas constitute the opercular and triangular parts of the inferior frontal gyrus (IFG), regions classically associated with Broca's area and language production 1317. Tractography reveals that human BA 44 and 45 exhibit connectivity profiles strikingly similar to macaque F5, particularly regarding robust white-matter tracts connecting to the inferior parietal lobule, including the angular and supramarginal gyri 131718. Furthermore, visual input regarding biological motion is fed into this frontoparietal circuit from the superior temporal sulcus in both species 1215.
| Macaque Cortical Region | Putative Human Homologue | Primary Functional Role in Mirroring |
|---|---|---|
| Area F5 (Ventral Premotor Cortex) | Brodmann Areas 44 / 45 (Inferior Frontal Gyrus) | Encoding the motor goals and execution parameters of goal-directed actions 31317. |
| Area PF / PFG (Inferior Parietal Lobule) | Supramarginal Gyrus / Angular Gyrus | Integrating kinematic and somatosensory information; classifying action typologies 13. |
| Superior Temporal Sulcus (STS) | Posterior Superior Temporal Sulcus | Visual processing of biological motion; feeding perceptual data to the frontoparietal mirror network 1215. |
Functional Bifurcation of Social and Non-Social Pathways
Early functional MRI studies conceptualized the human mirror neuron system as a unitary frontoparietal circuit that responded uniformly to observed actions. However, aggregate data analyses have revealed that the system dynamically reconfigures its topography based on the intrinsic sociality of the observed stimulus. A comprehensive meta-analysis encompassing 174 fMRI studies established that the human mirror neuron system bifurcates into distinct functional pathways when processing social versus non-social actions 192021.
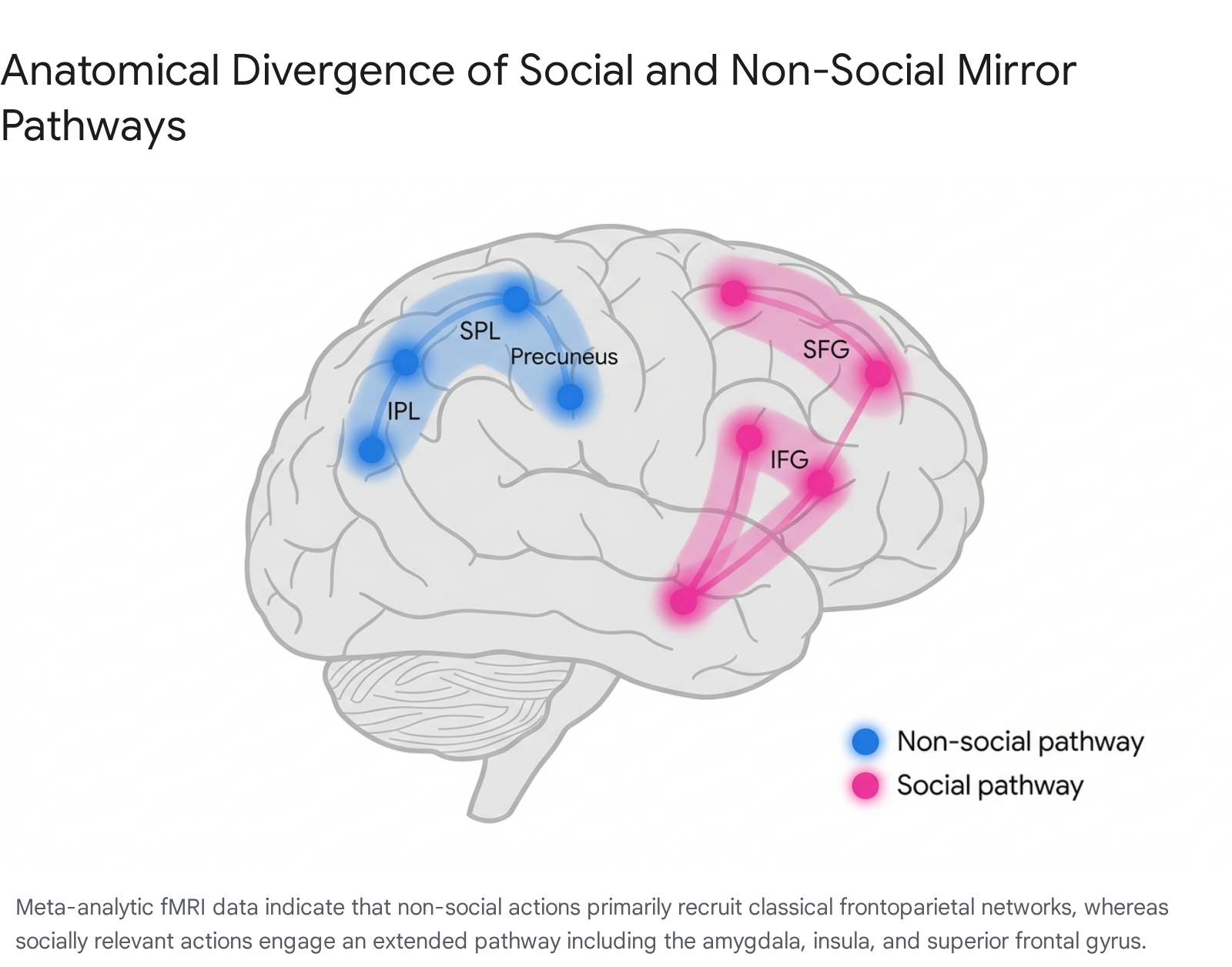
When humans observe non-social, object-directed actions - such as grasping a tool or manipulating an inanimate object - the resulting neural activation adheres to the classical frontoparietal architecture. This non-social mirror pathway is heavily concentrated in the bilateral inferior parietal lobe, the left superior parietal lobule, the bilateral postcentral gyrus, and the bilateral precuneus 1921. The dense parietal involvement reflects a high demand for spatial kinematics, object affordance processing, and exact motoric mapping.
Conversely, the observation of socially relevant actions - such as communicative gestures or interpersonal touch - recruits a markedly different circuitry. The social mirror pathway exhibits attenuated activation in the parietal lobe but significantly heightened engagement of the frontal lobe, the limbic system, and sublobar regions 1920. Key regions recruited during social mirroring include the bilateral inferior frontal gyrus, the right superior frontal gyrus, the right precentral gyrus, the bilateral amygdala, and the right insula 1920.
This bifurcation provides critical neurobiological evidence that the mirror neuron system is not a blind motor-resonance engine, but rather a system exquisitely sensitive to social valence. Furthermore, these pathways are heavily modulated by the specific physical effector performing the action. Social actions predominantly map onto neural representations of facial movements, automatically recruiting affect-related regions like the left amygdala and right IFG independent of whether the observed action is performed by a face or a hand 19. Non-social actions, meanwhile, map more closely onto robust hand-movement representations in the somatosensory and precentral gyri 19.
Methodological Critiques and the Problem of Reverse Inference
The assumption that mirror neurons provide humans with direct, unmediated experiential access to the mental states of others has been heavily critiqued as an unwarranted "inferential leap" 92223. Critics within the cognitive sciences argue that attributing high-level intentional state recognition (such as understanding why someone is acting) to low-level sensorimotor neurons conflates subpersonal neural activity with personal-level psychological competences, an error often classified as the mereological fallacy 9.
This debate exposes the methodological vulnerability of "reverse inference" in functional neuroimaging research. Reverse inference occurs when researchers observe activation in a specific brain region during a task and deduce that a particular cognitive process must be occurring, based solely on prior literature associating that region with that process 924. Because the brain regions housing the putative human mirror neuron system - such as the inferior frontal gyrus and the inferior parietal lobule - are highly heterogeneous and routinely involved in domain-general cognitive functions like working memory, attention, and executive control, equating their activation directly with "empathic simulation" or "mindreading" is deductively invalid 2324.
Challenges to Mu Suppression as a Valid Biomarker
Because functional MRI provides excellent spatial resolution but poor temporal resolution, many researchers have turned to electroencephalography (EEG) to measure mirror neuron activity in real-time. The primary EEG metric used to infer human mirror neuron engagement is the suppression, or desynchronization, of the mu rhythm 252627. The mu rhythm is an electrical oscillation in the 8 - 13 Hz frequency band detected over the sensorimotor cortex during states of physical rest 2628. When an individual executes a movement, or observes someone else executing a movement, the amplitude of these waves decreases - a phenomenon termed mu suppression 2628.
For years, mu suppression was treated as a direct, non-invasive biomarker for mirror neuron activation, and was utilized extensively to measure social cognition and empathy deficits in clinical populations 2627. However, recent methodological reviews and registered reports have demonstrated that mu suppression is highly sensitive to general attentional and executive control demands 2729.
When tasks require sustained visual attention, working memory allocation, or spatial reasoning, the mu rhythm desynchronizes even in the absence of biological motion or social observation. The failure of many historical mu suppression studies to adequately control for these broad attentional factors severely confounds the interpretation of their results 2729. While mu suppression remains a useful index of general sensorimotor engagement, its direct one-to-one mapping onto specific mirror neuron populations or its use as definitive proof of higher-order social cognitive states is now viewed with substantial skepticism by the neuroscientific community 2729.
Theoretical Evolution in Social Cognition
For nearly two decades, the dominant theoretical framework posited that mirror neurons were an innate, hardwired evolutionary adaptation explicitly selected to facilitate action understanding, imitation, and social bonding 1930. This classical view suggested that humans automatically comprehend another's intentions by simulating their motor plans via an unmediated, bottom-up neural resonance, providing a distinct evolutionary advantage for group cohesion 9.
However, as empirical data accumulated, this adaptationist account struggled to explain the extreme plasticity of mirror neurons, prompting a paradigm shift toward developmental and computational models of social cognition.
The Learned Matching Hypothesis
In response to the limitations of the innate adaptation model, an alternative framework known as the "Learned Matching" hypothesis has gained substantial empirical traction 831. Advanced by theorists such as Cecilia Heyes, this model posits that mirror neurons are not specialized genetic adaptations for social cognition, but rather "cognitive gadgets" - byproducts of domain-general associative learning mechanisms operating within a culturally rich social environment 830.
According to the associative learning account, mirror neurons acquire their matching properties through correlated sensorimotor experience during early development 832. When an infant performs a random, spontaneous motor act - such as smiling, vocalizing, or reaching - and a caregiver immediately imitates that behavior, the infant simultaneously experiences the descending motor command in their frontal cortex and the ascending visual feedback of the action from their temporal lobe 8. Through classical Hebbian learning principles ("neurons that fire together, wire together"), bidirectional synaptic links are forged between visual and motor representations over thousands of daily interactions 833.
This framework successfully explains why mirroring responses are highly plastic and experience-dependent. Laboratory training paradigms have demonstrated that researchers can easily generate "counter-mirror" neurons - cells that fire upon observing an action completely different from the one they execute - or establish robust mirror-like neural responses to entirely non-biological stimuli, such as arbitrary geometric shapes or abstract auditory cues, simply by manipulating sensorimotor contingencies 313234. If mirror neurons were a hardwired evolutionary adaptation explicitly for biological social cognition, such rapid functional reassignment to arbitrary stimuli would not be possible 832.
Predictive Coding in the Action Observation Network
Integrating the learned matching hypothesis with modern computational neuroscience, the framework of predictive coding offers a highly sophisticated model for how the action observation network functions in real-time 2533. Predictive processing shifts the paradigm away from a passive, feedforward motor simulation toward an active, hierarchical inference model of brain function 2433.
Within the action observation network, higher-order cortical regions (such as the premotor cortex) act as generative models, continuously producing top-down predictions regarding the expected sensory consequences of an observed action based on contextual cues, learned contingencies, and prior experience 33. These predictions are sent backward down the cortical hierarchy to sensory regions, such as the superior temporal sulcus. Simultaneously, incoming visual data about the observed action flows forward up the hierarchy.
If the incoming visual data deviates from the top-down prediction, a "prediction error" is generated. This prediction error propagates upward to update and refine the internal model 3334. In this framework, mirror neuron activity is not a passive reflection of another's movement, but rather the dynamic minimization of prediction error during social interaction 2433. When an individual successfully anticipates another's action, the top-down predictions successfully suppress the incoming sensory signal, leading to efficient, synchronized neural processing.
Deconstructing Empathy and Affective Resonance
The popular characterization of mirror neurons as the fundamental source of human empathy conflates distinct psychological constructs and oversimplifies complex neurobiological architecture. Empathy is a multidimensional phenomenon comprising at least three interacting components: automated motor mimicry, affective resonance, and explicit cognitive perspective-taking 101135. The human brain segregates these functions into distinct, though highly networked, neural systems 91136.
| Empathy Component | Functional Description | Primary Neural Substrates | Relationship to the Mirror Neuron System |
|---|---|---|---|
| Motor Empathy / Mimicry | Automatic imitation of observed gestures, facial expressions, and postures. | Inferior Frontal Gyrus (IFG), Inferior Parietal Lobule (IPL), Premotor Cortex. | Directly relies on the classical frontoparietal Action Observation Network / MNS 1125. |
| Affective Resonance | "Emotional contagion"; sharing the aversive or positive emotional state of another person. | Anterior Insula (AI), Anterior Cingulate Cortex (ACC), Amygdala, Temporal Pole. | Interacts with the MNS, but the actual visceral experience of emotion relies on the distinct "pain matrix" and limbic system 1011. |
| Cognitive Empathy | "Theory of Mind" (ToM) / Mentalizing; explicitly inferring the thoughts, beliefs, or intentions of another. | Temporoparietal Junction (TPJ), Medial Prefrontal Cortex (mPFC), Precuneus. | Functionally distinct from the MNS; relies on abstract representations rather than sensorimotor simulation 91137. |
Affective resonance - frequently referred to in the literature as emotional contagion - describes the process whereby perceiving another's emotional state triggers a homologous, visceral affective response in the observer 11. When individuals witness others experiencing physical pain (e.g., a needle piercing a hand) or social pain (e.g., social exclusion or emotional distress), functional neuroimaging consistently reveals robust activation in the anterior insula and the anterior cingulate cortex 1038. While the classical mirror neuron system may facilitate the initial motor simulation of an emotional expression - such as automatically mirroring a grimace or a smile - the actual internal, somatic experience of that emotion relies predominantly on these deeper visceromotor networks 19.
Cognitive empathy, or mentalizing, operates on an entirely different level of abstraction. It requires the individual to explicitly construct a model of another's internal world while maintaining a rigorous self-other distinction, ensuring that the observer does not confuse their own beliefs with those of the agent 11. The Theory of Mind network, anchored by the temporoparietal junction, the precuneus, and the medial prefrontal cortex, governs this capacity 937. Neuroscientific consensus now indicates that while the mirror neuron system provides crucial low-level, embodied input regarding the physical kinematics of an observed action, it lacks the computational architecture to independently extract complex intentionality, process abstract beliefs, or maintain the self-other boundary required for mature, regulated empathy 911.
Clinical Manifestations of Atypical Mirroring and Empathy
Investigations into clinical and neurodivergent populations provide critical insights into how the sub-components of empathy, action observation, and social cognition can be uncoupled when specific neural hubs or their regulatory connections are disrupted.
Rethinking Autism Spectrum Conditions
For many years, the "Broken Mirror Hypothesis" (BMH) served as the dominant neurobiological framework for explaining the social phenotype of autism. The BMH postulated that a global, structural dysfunction within the core mirror neuron system directly caused the deficits in social communication, empathy, and imitation characteristic of autism spectrum conditions (ASC) 394041.
However, as experimental designs became more rigorous, large-scale fMRI and EEG studies failed to find consistent, global mirror neuron impairments in autistic individuals. Research demonstrated that basic visuo-motor matching mechanisms are often entirely intact in ASC; autistic children can successfully execute and understand goal-directed actions, and can imitate precisely when explicitly instructed to do so 293940. Consequently, the BMH has been largely superseded by models emphasizing selective pathway dysfunction and aberrant top-down cognitive regulation.
| Theoretical Model of Autism | Core Proposed Mechanism | Predictions Regarding Behavioral Phenotype | Current Empirical Status |
|---|---|---|---|
| Broken Mirror Hypothesis (BMH) | Global dysfunction within the core MNS (IFG/IPL). | Global failure in action observation, execution, and imitation tasks regardless of context. | Largely unsupported; fails to account for intact basic goal-directed imitation in ASC 394041. |
| EP-M Model | Divergence in Emulation/Planning versus Mimicry pathways. | Goal-directed emulation (P-pathway) is intact. Non-goal-directed, automatic mimicry (M-route) is specifically impaired. | Supported by evidence showing ASC individuals struggle selectively with non-meaningful action imitation 3940. |
| STORM Model | Dysfunction in Social Top-Down Response Modulation (mPFC). | The MNS is structurally intact but lacks top-down regulatory input regarding the social context of an action. | Supported; explains why ASC individuals can imitate when instructed, but lack spontaneous, socially modulated mimicry 3940. |
The EP-M model refines the anatomy of imitation by proposing distinct routes for processing actions. It posits that the pathway serving goal-directed emulation - running from the middle temporal gyrus (MTG) to the inferior parietal lobule to the inferior frontal gyrus - remains intact in ASC. Conversely, the direct "mimicry route" connecting the MTG straight to the IFG, which supports the automatic imitation of non-goal-directed, socially affiliative behaviors, is selectively impaired 3940.
Simultaneously, the Social Top-Down Response Modulation (STORM) model posits that the mirror neuron system in ASC is fundamentally healthy, but it does not receive appropriate top-down regulatory signals from the medial prefrontal cortex. In neurotypical individuals, the mPFC continually modulates the mirror neuron system based on the social context of a situation, facilitating spontaneous mimicry during social bonding. In ASC, atypical connectivity from the mPFC leaves the mirror system unguided by social relevance, resulting in reduced spontaneous imitation without a fundamental inability to perform the motor actions 3940.
Therapeutic interventions are beginning to leverage these refined network models. Experimental protocols utilizing intermittent theta-burst stimulation (iTBS) - a non-invasive form of transcranial magnetic stimulation - over the bilateral inferior frontal gyrus and inferior parietal lobule in adolescents with ASC have demonstrated transient alterations in gamma oscillatory patterns. These neurophysiological shifts correlate with modest but significant improvements in clinical measures of social reciprocity, suggesting that targeted neuromodulation of the mirror and regulatory networks may enhance adaptive plasticity 4242.
Borderline Personality Disorder and the Empathy Paradox
Borderline Personality Disorder (BPD) presents an "empathy paradox" that beautifully illustrates the anatomical and functional independence of affective resonance from cognitive regulation. Individuals diagnosed with BPD frequently exhibit intense emotional reactivity and rapid emotional contagion, indicating that their bottom-up affective resonance mechanisms are not only intact but hyper-responsive 11. Functional neuroimaging of BPD cohorts observing emotional stimuli consistently reveals hyperactivation of the amygdala and the anterior insula, the core structures of the visceral pain matrix 11.
However, this heightened bottom-up resonance is coupled with profoundly impaired top-down cognitive regulation. Inefficient interactions between the affective limbic networks and the cognitive control networks result in a diminished capacity for explicit mentalizing and self-other distinction, processes governed primarily by the temporoparietal junction 11. Consequently, when observing another person in distress, an individual with BPD is overwhelmed by the shared somatic emotion (personal distress) but lacks the cognitive architecture to appropriately contextualize the emotion as belonging to the other person, hindering the deployment of regulated, prosocial empathic concern 11.
Cross-Cultural Modulators of Empathy and Social Neuroscience
While foundational neuroscience frequently treats empathy as a universal, hardwired biological mechanism, emerging research in cultural neuroscience has demonstrated that the neural correlates of affective resonance are highly malleable and subject to profound top-down contextual modulation 4344. Empathy is rarely deployed indiscriminately; its neural execution is gated by social identity, perceived similarity, and prevailing cultural norms.
In-Group Bias and Intergroup Neural Resonance
Neuroimaging studies consistently demonstrate powerful racial and cultural in-group biases in the recruitment of empathy-related neural networks 104345. When viewing the physical or social pain of an in-group member, observers typically exhibit significantly heightened activation in the anterior cingulate cortex, bilateral insula, and specifically the medial prefrontal cortex compared to when viewing out-group members experiencing identical distress 104346.
The medial prefrontal cortex is critical for self-referential processing and identity representation. Its selective activation in response to in-group pain suggests that perceived similarity facilitates a deeper, more personalized neural simulation of suffering 4346. Furthermore, observing out-group distress can sometimes elicit neural activation associated with disparate moral and self-conscious emotions rather than pure empathy. In a study of post-apartheid South African cohorts, participants viewed videos from the Truth and Reconciliation Commission depicting victims of political violence experiencing genuine emotional distress. While all participants showed basic amygdala and mentalizing network responses, White participants exposed to Black out-group distress exhibited heightened neural signatures associated with shame and guilt. Crucially, these self-conscious moral emotions were statistically associated with blunted activation in the core empathic networks, suggesting that profound intergroup conflict reshapes the automaticity of shared affective resonance 1047.
These neural biases are, however, deeply neuroplastic. Longitudinal fMRI studies indicate that repeated, meaningful interactions with out-group cultures can effectively reverse racial in-group biases in empathic brain activity. Individuals who spent extended periods immersed in a foreign culture (e.g., Caucasian students living in East Asia) demonstrated strengthened neural responses to the pain of the host culture's demographic upon return, suggesting that extended intercultural exposure recruits neuroplastic mechanisms to expand the cognitive boundaries of the "in-group" 45.
Cultural Dimensions, Collectivism, and the Global South
A significant limitation in current social cognitive neuroscience is the overrepresentation of data derived from Western, Educated, Industrialized, Rich, and Democratic (WEIRD) populations 484950. Extrapolating neurobiological models of empathy globally presents severe validity issues, as behavioral and neurobiological expressions of empathy vary distinctly along the individualism-collectivism spectrum 435152.
Cultures prioritizing collectivism, familial interdependence, and social harmony foster environments where adapting to the needs of others is a paramount developmental goal 4352. Behavioral and neuroimaging data suggest that individuals from highly collectivistic societies engage the cognitive components of empathy - such as mentalizing and perspective-taking - more automatically, spontaneously, and with less cognitive effort than individuals from individualistic societies, who tend to prioritize ego-centric frameworks 43.
In studies comparing socio-emotional development in the Global South, such as among adolescents in Guatemala, researchers observed that specific economic and cultural contexts deeply influence empathy. Guatemalan adolescents attending schools in environments of deep economic scarcity, yet immersed in a culture prioritizing familism, respectfulness, and spirituality, reported significantly higher levels of daily cognitive and affective empathy than peers in higher-resource individualistic settings 5253.
The lack of culturally validated social cognition assessments for populations in South America, Africa, and parts of Asia remains a critical bottleneck for the field. Standardized tests measuring the Theory of Mind or emotion recognition (e.g., the Hinting Task or the 'Reading the Mind in the Eyes' test) rely heavily on Western linguistic idioms, social norms, and Caucasian facial morphotypes. When applied without rigorous cultural adaptation, these tools compromise diagnostic validity, risking the mischaracterization of social cognitive deficits in neurodegenerative conditions like frontotemporal dementia or schizophrenia in non-Western patients 4954.
Metabolic and Physiological Constraints on Prosocial Behavior
Finally, the translation of affective resonance into actionable prosocial behavior is strictly constrained by fundamental physiological states, underscoring the severe biological and metabolic costs of empathy. The generation of a robust empathic response, followed by a decision to assist another, requires significant cognitive and metabolic resource allocation 55.
Recent neurobiological research highlights the interface between homeostatic energy balance and complex prosocial behavior. In mammalian models, negative energy balance - specifically starvation or acute hunger - actively suppresses helping behaviors. This suppression is mediated by agouti-related peptide (AgRP) neurons in the hypothalamus, which serve to prioritize self-preservation and feeding behaviors over the highly energy-demanding cognitive processes required for empathy and altruistic rescue 55. When an animal is well-fed, forebrain ATP/ADP ratios decline sharply during active helping tasks, indicating that prosociality requires a substantial, quantifiable caloric investment 55. Consequently, environmental stressors, including severe resource scarcity and nutritional deficits, directly modulate the neural execution of empathetic behavior at the cellular level, anchoring the lofty concepts of social cognitive neuroscience firmly within the constraints of systemic evolutionary biology.
Conclusion
The proposition that mirror neurons serve as a singular, localized "empathy engine" is a theoretical oversimplification that has been largely retired by contemporary cognitive neuroscience. While the mirror neuron system - comprising the inferior frontal gyrus and inferior parietal lobule - undeniably provides a foundational sensorimotor mechanism for mapping the physical kinematics of observed actions onto the self, it does not operate in isolation to generate complex social understanding.
Current evidence favors the integration of mirror neurons within broader, highly distributed cortical networks governed by hierarchical predictive coding and shaped extensively by lifelong associative learning. Empathy itself emerges not from simple motor mirroring, but from the highly orchestrated interplay between the visuo-motor mapping of the mirror neuron system, the visceral affective resonance pathways of the limbic system, and the sophisticated cognitive regulatory mechanisms of the Theory of Mind network. Furthermore, the robust modulation of these neural networks by cultural context, in-group identity, and even fundamental metabolic states underscores that human social cognition is a dynamic, highly plastic phenomenon, fundamentally dependent on continuous environmental and cultural interaction rather than strict genetic determinism.