Biological foundations of human social hierarchy
The architecture of human social organization represents a profound evolutionary puzzle. Unlike many species characterized by rigid, species-typical social structures, humans exhibit extraordinary behavioral plasticity. The anthropological and historical records demonstrate that humans are capable of sustaining strictly stratified, despotic autocracies as well as fiercely egalitarian, stateless communities. Determining whether human beings are biologically "hardwired" for hierarchy requires a comprehensive synthesis of comparative primatology, evolutionary anthropology, cognitive neuroscience, and behavioral endocrinology. The accumulated empirical evidence indicates that humans do not possess a singular, inescapable drive for coercive dominance. Rather, human neurobiology and endocrinology provide a highly specialized, adaptable toolkit optimized for navigating complex, shifting social ranks. These biological systems support parallel strategies for status acquisition - ranging from ancient mammalian dominance mechanisms to uniquely human prestige pathways - while simultaneously possessing deeply evolved neurological mechanisms for suppressing hierarchy when ecological or social contexts demand egalitarian cooperation.
Evolutionary Primatology and the Pan Clade
To understand the biological foundations of human hierarchy, researchers rely heavily on the Pan-Homo clade, which offers the most direct comparative framework for tracking the evolutionary origins of human social behavior. Humans share approximately 98 to 99 percent of their genomic sequence with the two extant species of the Pan genus: the chimpanzee (Pan troglodytes) and the bonobo (Pan paniscus) 12. These two species diverged from a common ancestor with humans approximately six to eight million years ago, and from each other merely one to two million years ago 22. Despite their extremely close phylogenetic relationship and morphological similarities, chimpanzees and bonobos exhibit drastically divergent social systems that represent the two extreme hierarchical poles that the human psyche is capable of navigating.
Chimpanzee and Bonobo Social Structures
Chimpanzee societies are characterized by strict, despotic hierarchies that are overwhelmingly male-dominated. Within chimpanzee communities, robust competition exists among males for dominance and priority of access to mating opportunities and resources 1. Male chimpanzees frequently employ physical coercion, aggressive displays, and the formation of strategic, agonistic alliances to achieve and maintain alpha status 123. The hierarchy is actively enforced through intimidation and, occasionally, lethal violence. Furthermore, this aggressive disposition extends beyond the internal group dynamics; male chimpanzees are highly territorial and engage in cooperative border patrolling and lethal intergroup encounters that researchers frequently compare to primitive forms of human warfare 34. Because female dispersal is the standard pattern in chimpanzee communities, the core social and political bonds are forged almost entirely among related and allied males 14.
In stark contrast, bonobos reside in highly egalitarian, female-centered communities. Although male bonobos are physically larger than females, the females hold the highest social status. They achieve this not through individual physical dominance, but by forming strong, cooperative female-female coalitions that actively and effectively suppress male aggression 125. The social atmosphere of a bonobo community is notably peaceful and lacks the rigid, coercive stratification seen in chimpanzees. Lethal intergroup encounters are entirely absent in the bonobo behavioral repertoire; groups from different communities frequently mingle without conflict, and individuals from separate groups may interact peacefully or even engage in sexual behavior 34. Instead of utilizing violence to resolve disputes, bonobos employ frequent socio-sexual behavior and adult play to diffuse tension, negotiate the sharing of resources, and strengthen community bonds 12.
The reproductive biology of bonobos structurally undermines the utility of coercive male dominance. Bonobo females exhibit an extended estrus period that spans nearly the entire month, and they actively disguise ovulation 4. Because male bonobos lack the cues necessary to identify when a female is ovulating, aggressive males cannot effectively monopolize fertile females. This reproductive dynamic fundamentally alters the evolutionary calculus, drastically reducing the fitness benefits of maintaining a despotic male hierarchy and promoting an egalitarian social structure 4.
Neurobiological Divergence in Apes
The divergent hierarchical behaviors of chimpanzees and bonobos are not merely cultural or ecological artifacts; they are reflected deeply in the distinct neuroanatomies of the two species. Advancements in neuroimaging, specifically diffusion tensor imaging (DTI) and voxel-wise analysis of T1-weighted MRI images, have elucidated the neurobiological substrates underlying these behavioral differences. Research indicates that bonobos possess significantly more gray matter in brain regions critically associated with perceiving distress in both oneself and others, specifically the right dorsal amygdala and the right anterior insula 2.
Crucially, bonobos exhibit a substantially larger and more robust neural pathway linking the amygdala to the ventral anterior cingulate cortex (vACC) and the ventromedial prefrontal cortex (vmPFC) 2. In hominids, this specific neural circuit is heavily implicated in the top-down cognitive control of aggressive impulses and the bottom-up relay of empathetic distress signals. The hypertrophy of this network in bonobos suggests a profound neurobiological foundation for their heightened empathic sensitivity, social tolerance, and aversion to causing harm. This neural architecture limits distress and anxiety to levels that are conducive to prosocial behavior, thereby facilitating the maintenance of an egalitarian social structure 2.
Conversely, the chimpanzee brain exhibits adaptations suited to their specific social and ecological demands. Brain imaging reveals that chimpanzees possess greater development in visual system pathways - including the right optic tract and splenium of the corpus callosum - and have a larger cerebellum relative to overall brain size 2. These adaptations likely support the intense motor and cognitive demands required for hunting, rapid navigation during territorial conflicts, and the continuous monitoring of shifting male alliances within a highly volatile dominance hierarchy 2.
| Biological and Social Trait | Chimpanzee (Pan troglodytes) | Bonobo (Pan paniscus) | Human (Homo sapiens) |
|---|---|---|---|
| Primary Social Hierarchy | Male-dominated, strict, despotic | Female-centered, egalitarian | Highly flexible (Egalitarian to Despotic) |
| Intergroup Dynamics | Highly territorial, lethal warfare | Peaceful mingling, low territoriality | Highly variable (Warfare to cooperative trade) |
| Conflict Resolution Mechanism | Coercive aggression, male alliances | Socio-sexual behavior, grooming, play | Complex negotiation, legal systems, warfare |
| Primary Social Bonding | Strong male-male alliances | Strong female-female coalitions | Multi-sex bonding, monogamy and polygyny |
| Reproductive Strategy | Male monopolization of ovulation | Concealed ovulation, extended estrus | Concealed ovulation, long-term pair bonding |
| Key Neural Adaptations | Enhanced visual pathways, larger cerebellum | Hypertrophied amygdala-vmPFC pathway | Highly developed prefrontal cortex, extreme plasticity |
The Deconstruction of Canid Dominance Models
For decades, the public and scientific understanding of mammalian social hierarchy was heavily influenced and distorted by the concept of the "alpha male" wolf. This paradigm incorrectly framed animal social structures as inherently reliant on relentless, aggressive, physical dominance, and it was frequently extrapolated to human behavior to justify rigid social stratification.
Wild versus Captive Canid Dynamics
The concept of the aggressive "alpha wolf" originated in the 1940s from observational studies conducted by biologist Rudolf Schenkel. Schenkel studied wolves in captivity at a European zoo 78. The subjects of his study were unrelated adult wolves that had been captured from various locations and forced into an artificial, confined enclosure. Under these unnatural, high-stress conditions - characterized by constrained resources, lack of kin relationships, and an inability for subordinate animals to disperse - the wolves engaged in frequent, violent dominance displays to establish a strict pecking order 7610. The researchers mistakenly identified this stress-induced, captive behavior as the natural state of the species, concluding that wolves operate in a constant, violent struggle for supremacy 71011.
This paradigm remained largely unchallenged until the late twentieth century, when wildlife biologists, most notably L. David Mech, began tracking and observing wild wolves in their natural habitats. Mech's extensive field research in locations such as the Canadian Arctic and Minnesota demonstrated that the captive observations were fundamentally flawed and biologically misrepresentative 7811. In the wild, wolf packs do not consist of random, unrelated adults fighting for rank. Rather, wild wolf packs are nuclear families. A pack is typically built around a single breeding pair - the parents - and their offspring from the current and previous years 611. The individuals previously and incorrectly labeled as "alpha males" or "alpha females" are simply the parents guiding, feeding, and protecting their young. Leadership within the wild pack is achieved naturally through parental care, age, cooperation, and survival experience, rather than through violent, agonistic dominance displays 611.

Cultural Persistence and Biological Determinism
Despite the ethological literature correcting the alpha myth decades ago, the flawed concept deeply permeated popular culture and adjacent scientific disciplines. It was widely co-opted to justify aggressive, patriarchal human hierarchies, toxic corporate leadership models, and specific socio-economic frameworks as inescapable biological imperatives 7811.
The misapplication of captive wolf dynamics to human behavior serves as a critical case study in the dangers of biological determinism. The extrapolation of stress-induced animal behaviors to validate coercive human dominance fails to account for natural ecological contexts. Recognizing that natural canid hierarchies are kin-based, stable, and highly cooperative aligns much more closely with the evolutionary history of early human kin groups and nomadic hunter-gatherer bands, which relied on mutual support rather than internal violence for survival 86.
Hunter-Gatherer Egalitarianism and Reverse Dominance
For the vast majority of Homo sapiens' approximately 300,000-year history, human beings lived exclusively in nomadic foraging bands 127. Anthropological and ethnographic observations of extant immediate-return hunter-gatherer societies - such as the Ju/'hoansi (or !Kung) of the Kalahari Desert, the Mbuti of the Congo rainforest, the Hadza of Tanzania, and various Australian Aboriginal groups - reveal a striking and consistent absence of formal hierarchy, centralized authority, or material inequality 1415817.
In these societies, individuals live in small, self-governing bands of roughly 20 to 50 people. Because they move continuously to exploit seasonal resources, the accumulation of surplus material wealth is practically impossible, thereby eliminating the economic foundations of social class 1218. However, the absence of material wealth does not automatically guarantee political equality. Anthropologist Christopher Boehm theorizes that human egalitarianism is not the result of a biological lack of dominance drives; rather, humans retain the phylogenetically ancient primate impulse to dominate 1214. Hunter-gatherer equality is instead an active, meticulously engineered social construct known as a "reverse dominance hierarchy" 51519.
Active Leveling Mechanisms
In a reverse dominance hierarchy, the rank and file of the community cooperatively unite to actively dominate and suppress any individual who attempts to assert uninvited authority, hoard resources, or elevate themselves above the group 51519. The maintenance of equality requires constant social vigilance and the proactive deployment of specific cultural leveling mechanisms.
When a group member exhibits boastfulness, arrogance, or aggressive status-seeking behavior, the group responds with a calibrated escalation of social sanctions. These sanctions invariably begin with subtle social cues, such as criticism, ignoring the individual, and direct, public ridicule 5159. A thoroughly documented ethnographic example of this phenomenon is the practice of "insulting the meat," observed among the Ju/'hoansi and other African foragers. When a hunter successfully kills a large animal, they are strictly expected to display extreme humility. The rest of the band deliberately and persistently mocks the size and quality of the kill - suggesting the meat is scrawny or worthless - ensuring that the hunter's heart is "cooled" and that they are prevented from using their hunting prowess to leverage political power or demand subservience 810.
Furthermore, the distribution of vital resources is heavily codified to prevent the accumulation of social debt. Among many groups, the "own kill rule" dictates that the hunter cannot consume or distribute the meat from their own kill. The distribution rights are often assigned to the individual who owned the arrow that killed the animal, and the meat is partitioned by designated community members in a public setting 81022.
If ridicule and institutional humility fail to suppress a domineering individual, hunter-gatherer groups predictably escalate to disobedience, temporary shunning, and ultimately, outright ostracism 51519. In extreme cases within prehistoric and some documented ethnographic contexts, anthropologists, including Richard Wrangham, hypothesize that coordinated capital punishment was utilized. Subordinate males would form murder conspiracies to eliminate violently aggressive, despotic individuals, an evolutionary process sometimes referred to as human self-domestication 12. Through this process, early human communities exerted intense selective pressure against reactive aggression, bullying, and the monopolization of mates, fundamentally shaping the human genome toward prosociality 12.
Play and Social Cohesion
In addition to punitive sanctions, play serves as a vital evolutionary counterweight to dominance in humans, much as it does in other highly social mammals like bonobos 5. Hunter-gatherer societies are characterized by a pervasive playfulness that actively undermines hierarchical tension. Religious rituals, conflict resolution mechanisms, and the daily distribution of resources are often mediated through humor, self-deprecation, and play 5819.
Play inherently requires the temporary suspension of rank and the adoption of an egalitarian stance among participants; one cannot effectively play while simultaneously asserting dominance 59. The integration of play into the foundational practices of the band ensures that social bonds are continuously renewed and that the physiological stress associated with hierarchical jockeying is consistently dissipated.
The Dual Strategies Theory of Human Status
While humans possess deeply ingrained egalitarian mechanisms and lived without formal hierarchies for hundreds of millennia, the advent of agriculture, the domestication of animals, the storage of food surpluses, and the transition to sedentary, dense populations approximately 10,000 to 12,000 years ago facilitated the rapid re-emergence of highly stratified societies 12141911. To navigate both the deeply egalitarian environments of our evolutionary past and the highly stratified environments of the Holocene, humans evolved a complex, bifurcated psychological approach to status acquisition.
The "Dual Strategies Theory," formulated by evolutionary anthropologists Joseph Henrich and Francisco Gil-White, posits that humans compete for social rank within groups using two distinct, viable evolutionary pathways: Dominance and Prestige 121314.
Dominance Dynamics
Dominance is the phylogenetically older strategy, directly homologous with the aggressive dominance hierarchies observed in nonhuman primates 121528. It involves the use of force, implied physical or economic threat, coercion, and general intimidation to seize and maintain power 121529. In a dominance-based hierarchy, status is not granted voluntarily; rather, subordinates defer to superiors strictly out of fear of punishment, injury, or the withholding of critical resources 121314.
Individuals who successfully utilize the dominance strategy typically display specific personality traits and affective states, including high levels of assertiveness, interpersonal aggression, manipulativeness, and hubristic pride, paired with relatively low levels of agreeableness and empathy 1214. While dominance can effectively ensure rapid compliance and coordinate group action - making it highly prevalent and sometimes selectively favored during times of severe intergroup conflict or acute crisis - these hierarchies are inherently unstable over the long term 1129. Subordinates operating within human dominance hierarchies frequently harbor deep resentment toward their superiors and will actively coordinate to overthrow, sabotage, or undermine the dominant individual when the opportunity or numerical advantage arises 29.
Prestige Pathways
Prestige is considered an evolutionary innovation unique to the human lineage. It is believed to have evolved in tandem with our species' extreme reliance on cumulative culture, complex tool use, and sophisticated social learning 131528. Status gained through prestige is freely and voluntarily conferred by followers upon an individual who demonstrates exceptional, culturally valued skills, wisdom, knowledge, or generosity 121329.
Unlike dominant individuals who forcibly seize status, prestigious individuals attract deference. Subordinates voluntarily submit to and seek close association with prestigious individuals in exchange for the opportunity to learn from them, receive guidance, or benefit from their high competence 1315. Prestige-based hierarchies are characterized by positive-sum outcomes; they rely on persuasion, agreeableness, and prosocial behavior 2916. Because the relationship is mutually beneficial, prestige hierarchies are significantly more stable and socially tolerated than dominance hierarchies 29.
Both strategies remain viable and operate simultaneously in modern human environments. Depending on the specific cultural context, organizational structure, or external environmental threats, human groups may lean heavily toward either prestige-granting or dominance-enforcing mechanisms 1115.
| Feature | Dominance Pathway | Prestige Pathway |
|---|---|---|
| Mechanism of Influence | Coercion, intimidation, threat of costs | Persuasion, skill demonstration, benefit generation |
| Origin of Status | Seized forcefully by the individual | Freely conferred by the community |
| Subordinate Motivation | Fear, avoidance of punishment or harm | Respect, desire to learn, admiration |
| Evolutionary Origin | Ancient mammalian (shared with primates) | Unique to humans (tied to cumulative culture) |
| Associated Traits | Aggression, hubristic pride, lower empathy | Generosity, high competence, agreeableness |
| Hierarchical Stability | Low (prone to subordinate rebellion and leveling) | High (mutually beneficial and cooperative) |
Neurobiological Architecture of Social Rank
Navigating the complexities of human hierarchies requires highly specialized neural architecture capable of instantly assessing rank, predicting social value, updating expectations, and calculating the behavioral consequences of interacting with superiors and subordinates. Functional magnetic resonance imaging (fMRI) and electroencephalography (ERP) studies have mapped the human "social brain network," identifying the specific cortical and subcortical regions heavily implicated in hierarchical processing.
Brain Networks and Social Tracking
The perception of dominance and the assessment of status cues occur rapidly, automatically, and early in the stream of visual processing 1718. Visual tracking of status symbols - such as dominant postures, facial expressions, or cultural decorations - engages the fusiform gyrus for deep facial encoding, the salience network for immediate evaluation of relevance, and the inferior frontal gyrus for interpreting abstract face ornaments or titles as symbols of rank 19.
When human subjects perceive individuals of higher status, they exhibit increased metabolic activity in several key brain regions. The ventral striatum, a subcortical region fundamentally associated with reward processing, prediction errors, and motivation, is heavily recruited to evaluate the expected value of social interactions and the allocation of resources within a group 14202122. The ventromedial prefrontal cortex (vmPFC) and the medial prefrontal cortex (mPFC) are deeply engaged for social cognition; these regions integrate social information regarding the self in relation to others and assign value to self-related information during decision-making 3723. Additionally, the dorsolateral prefrontal cortex (DLPFC) is recruited for executive control and cognitive flexibility, enabling the complex behavioral adjustments required to interact appropriately and safely with a superior 223739.
The tracking of social rank is highly evident even in naturalistic settings. For example, neuroimaging studies of adolescents navigating the popularity of peers within their real-world school networks demonstrate that the vmPFC and striatum precisely track high-status, popular individuals. Conversely, regions such as the temporoparietal junction (TPJ), the insula, and the midcingulate cortex are selectively engaged to track unpopular, low-status peers 39.
Hierarchy Instability and Stress Response
The neurological burden and physiological cost of hierarchy are highly dependent on the stability of the system. Functional MRI experiments, particularly those conducted by Zink and colleagues using interactive virtual games, demonstrate a profound neurological divergence based on hierarchy stability. Interacting within a stable hierarchy - where rank is fixed and cannot be changed by the outcome of a contest - primarily elicits activity in perceptual-attentional and cognitive processing systems, such as the striatum and DLPFC 171822.
However, when the hierarchy is unstable - meaning rank is fluid and can be lost or gained based on performance - viewing a superior rival triggers massive, additional recruitment of the amygdala and the mPFC 1722. This shift indicates that fluid social ranks are processed by the brain as emotionally salient, highly threatening environments that require constant behavioral readiness and heightened emotional processing 22.
Crucially, this stress is not distributed evenly; it is disproportionately borne by those at the top who face downward mobility. Controlled studies manipulating hierarchies in male mice reveal that social reorganization causes the most significant physiological stress and gene expression changes in the medial amygdala of the alpha males 2425. The findings confirm a direct correlation: the further a dominant individual falls in rank, the more extreme their physiological stress response.
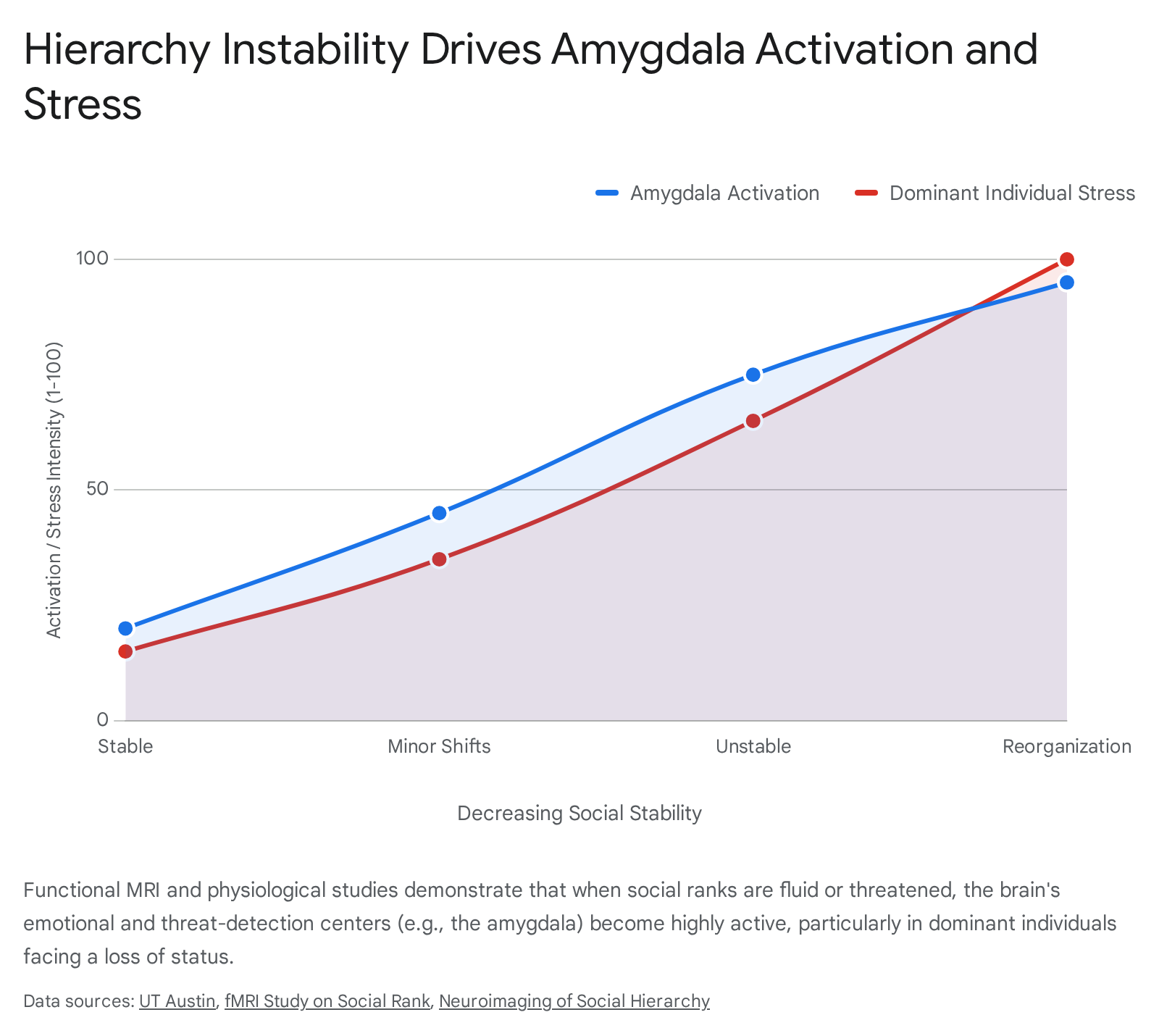
This demonstrates that maintaining the pinnacle of a dominance hierarchy, particularly an unstable one, requires immense, neurologically taxing effort, validating the evolutionary observation that it is indeed "not easy at the top" 2425.
Endocrinological and Neurochemical Mechanisms
Hormones and neurotransmitters provide the foundational chemical signaling required for status-seeking behaviors, social calibration, and the suppression of hierarchy. While traditional biological models painted a broad, reductive picture linking testosterone directly to aggression and dominance, modern behavioral endocrinology has developed highly nuanced, conditional models detailing how chemical signals interact to regulate social rank.
The Dual-Hormone Hypothesis
The Dual-Hormone Hypothesis represents a significant refinement in the understanding of endocrine regulation of status. It posits that testosterone's influence on dominance, status-seeking, and risk-taking behavior in humans is highly conditional, heavily modulated by cortisol - a glucocorticoid released primarily by the hypothalamic-pituitary-adrenal (HPA) axis during times of psychological stress or social avoidance 2627.
According to this hypothesis, naturally occurring or elevated testosterone levels are positively associated with assertive leadership, competitive drive, and dominance only when cortisol levels are concurrently low 2728. If an individual experiences high stress (reflected by high cortisol), the status-seeking behavioral effects of testosterone are either entirely blocked or reversed, potentially motivating lower-status behaviors 27. This dynamic suggests that individuals who possess a specific biological profile - high endogenous testosterone coupled with low baseline stress reactivity - are neuroendocrinologically primed to comfortably attain and maintain high-status positions. This specific interplay has been supported by analyses examining real-world hierarchies, showing that executives with high testosterone and low cortisol govern the highest number of subordinates 2729.
It is necessary to note, however, that the Dual-Hormone Hypothesis remains contested within the literature. Recent large-scale p-curve analyses and rigorous empirical evaluations of the hypothesis have indicated that many published studies documenting this effect suffered from remarkably low statistical power (estimated at roughly 16 percent power to reliably detect the claimed effects) 2630. Furthermore, independent replication attempts have occasionally yielded null findings when predicting status-striving personality features from the testosterone-cortisol interaction 2630. Thus, while the dual-hormone interaction provides a compelling theoretical model for conditional status-seeking, the precise hormonal mechanics remain a subject of ongoing debate and require further empirical validation.
Serotonin, Dopamine, and Social Valuation
At the neurochemical level, the processing of hierarchy and the execution of social decisions are heavily dependent on the continuous interplay between dopamine and serotonin. Studies monitoring these neurotransmitters in the human brain via carbon-fiber microelectrodes during awake deep brain stimulation surgeries have revealed highly specialized roles for each chemical during social bargaining (e.g., ultimatum games) 3132. Dopamine, functioning within the striatal pathways, acts as a continuous tracking system for relative value. It tracks whether an incoming social offer or interaction is better or worse than the previous one, processing the ongoing context of fairness and adjusting expectations accordingly 3132.
Conversely, serotonin acts as an immediate evaluator of current value and a regulator of social predictability. The dorsal raphe nucleus (DRN), which serves as the brain's primary headquarters for serotonin production, functions as a highly complex algorithm for social learning and hierarchy calculation 2833. Serotonin is crucial for decoding latent or ambiguous dominance hierarchies, allowing individuals to map predictive relationships and perceived control within their environment 33.
Furthermore, increased serotonergic function is deeply linked to prosocial behavior, harm avoidance, and the positive valuation of others' outcomes. Pharmacological studies demonstrate that elevated serotonin reduces negative reciprocity in the dorsal striatum and promotes cooperative, charitably minded decisions 34. High serotonin environments within the brain facilitate social conformity and bonding by mitigating the negative, antisocial impulses that drive zero-sum dominance competitions 2334.
Neuropeptides and Hierarchy Flattening
The ancient neuropeptides oxytocin and arginine vasopressin act as potent modulators of hierarchical behavior, serving largely to "flatten" social stratifications across primate species. Genetic analyses indicate that variation in the vasopressin receptor gene (AVPR1A) - specifically the length of the RS3 microsatellite located in the promoter region - is deeply associated with differences in social bonding, empathy, and prosociality among humans, chimpanzees, and bonobos 353637.
Experimental administrations of oxytocin and vasopressin via inhalation or injection in male rhesus macaques - a species historically noted for extremely despotic, aggressive dominance hierarchies - yield profound behavioral shifts. Neuropeptide exposure rapidly suppresses aggressive dominance behaviors and enhances egalitarian, prosocial interactions 3638. Under the influence of these hormones, high-ranking, dominant monkeys become relaxed and socially tolerant, reducing their threat displays. Concurrently, subordinate monkeys become significantly more confident, engaged, and less vigilant 38. Notably, this hierarchy-flattening effect occurs even if only one individual in a dyad receives the hormone, indicating that the neuropeptides alter subtle, non-verbal social cues and gestures that subsequently recalibrate the entire hierarchical dynamic of the group toward egalitarianism 38.
Similarly, in bonobos, oxytocin exposure significantly increases eye contact, a behavior tightly linked to human-like social bonding and empathy. This suggests that the evolutionary divergence of prosocial, egalitarian apes from their more violently competitive ancestors was heavily facilitated by adaptations within the oxytocinergic and vasopressinergic systems 39.
Ultimately, human neurobiology does not dictate an inescapable destiny of rigid, aggressive dominance. Instead, our biology provides a highly sensitive, socially responsive, and computationally complex toolkit. It equips humans with the capacity to assert dominance when threatened, the cognitive architecture to confer prestige upon the competent, the physiological stress responses to navigate instability, and the neurochemical mechanisms - driven by serotonin, oxytocin, and vasopressin - to successfully suppress hierarchy and forge fiercely egalitarian bonds when survival depends upon mutual cooperation.